










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.38 no.2 Bogotá July/Dec. 2012
First records for Cissia themis (Lepidoptera: Nymphalidae)
from Colombia and Venezuela
Primer registro para Cissia themis (Lepidoptera: Nymphalidae) de Colombia y Venezuela
SHINICHI NAKAHARA1, MARIO A. MARÍN2 and WATARU MITSUHASHI3
1 Biologist. 4-9-34-220 Shimoochiai, Shinjuku, Tokyo 161-0033 Japan. amathonte1@yahoo.co.jp, corresponding author.
2 M. Sc. Grupo de Investigación en Sistemática Molecular, Universidad Nacional de Colombia, sede Medellín. Calle 59A No 63-20. Bloque 16-112 Medellín, Colombia.
3 Dr. 3-26-5-402 Kamiogi, Suginami, Tokyo 161-0043 Japan.
Received: 30-Mar-2012 - Accepted: 11-Jul-2012
Abstract: A new country record for Cissia themis from Colombia and Venezuela is reported here. This record is also the first report for South America except for the island of Trinidad. Specimens of C. themis are often misidentified and found mixed in with its congener C. similis in public collections and so the morphology of C. themis is compared to its congener, C. similis.
Key words: Dry forest. Euptychiina. Morphology. Taxonomy. Neotropical Region.
Resumen: Se presenta aquí un nuevo reporte para Cissia themis en Colombia y Venezuela. Este reporte es también el primer Sur América excepto para la isla de Trinidad. Con frecuencia los especímenes de C. themis se identifican mal y se encuentran mezclados con su congénere C. similis en colecciones públicas, así se compara la morfología de C. themis con su congénere C. similis.
Palabras clave: Bosque seco. Euptychiina. Morfología. Taxonomía. Región Neotropical.
Introduction
Some of the most poorly known groups among the neotropical butterflies are members of the subfamily Satyrinae in the family Nymphalidae. As a result of their evolutionary success, it is known to be the most diverse subfamily within the family (Ackery et al. 1998; Peña and Lamas 2005; Marín et al. 2011). Most of the members of the subtribe Euptychiina (euptychiine Satyrinae species) inhabit lowland forest of the neotropics. Elements of the group's taxonomy are often poorly known or understood and many undescribed taxa remain. Additional comprehensive work in laboratories as well as in the field are needed to clarify the taxonomy of this group (DeVries 1994; DeVries et al. 1997; Peña and Lamas 2005; Huertas et al. 2009; Marín et al. 2011). A true understanding of taxonomy and geographical distribution of these poorly known taxa will be extremely valuable towards improving our understanding of Neotropical biodiversity. In this paper, we focus on the genus Cissia, which includes 15 described species, two-thirds of whose members are residents of Central America (Montero-Ramírez and González-Maya 2009). Cissia themis (Butler, 1867) is one of these Central American species and to date no record from the South American continent (delimited by the Darién watershed along the border of Colombia and Panama) is known for this species except for the island of Trinidad (Barcant 1970; Singer and Ehrlich 1991). Here, we report four newly discovered localities of occurrence for C. themis in Venezuela. The first author also detected a couple of Colombian specimens misidentified as “Cissia similis” in the MEFLG which are actually C. themis. All of these records are new country records.
Methods
Identification of Cissia themis was based on the original description included in Butler (1867) and the syntype in the NHM. Variations of wing pattern have been studied by the authors by examining specimens in private and public collections. Dissections for observing the male genitalia were made of specimens from localities spanning the range of this species in order to examine the variation and stability of characters within species. We compared these characters to the male genitalia of C. similis figured in Forster (1964). Adult abdomens were soaked in hot 10% KOH for 10-15 minutes, dissected and subsequently stored in glycerine. Body morphology and dissections were studied using a binocular microscope at 50x magnification. The terminology for genital structures largely follows Klots (1956), and nomenclature for venation follows Neild (1996). The following collection acronyms are used throughout this paper.
NHM The Natural History Museum, London, UK
USNM National Museum of National History, Smithsonian Institution, Washington, DC, USA
MGCL McGuire Center for Lepidoptera and Biodiversity, Florida Museum of Natural History, Florida, USA
MUSM Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Peru
MEFLG Museo Entomológico Francisco Luis Gallego, Universidad Nacional de Colombia, Medellín, Colombia
MEL Museo Entomológíco de León, León, Nicaragua
AN Andrew Neild collection, London, UK
SN Shinichi Nakahara collection, Tokyo, Japan
Examined specimens
Cissia themis Syntype: 1♀. (Figs. 2A, B). no locality data [NHM].
VENEZUELA. 1♀.Zulia. Río Chiquito zona de Reserva del Río Pueblo Viejo. 220 m. 28-jan-1998. A. Viloria & J. Camacho [MUSM].1♂1♀.Barinas. Bosque de Caimital. 200 m. 1-Nov-1999. A. Neild [AN]. 1♂. Sucre. El Rincón. 50 m. 28-Aug-2011. S. Nakahara [SN]. 1♂1♀. (Fig. 1.).Aragua. Villa de Cura. M. D. Escobar [photo record]; COLOMBIA. 3♂1♀. (Figs. 2C, D). Arauca. Arauca UNAL sede. (07°05'05”N 70°45'42”W) 200 m. 20-Jun-2003. C. F. Álvarez [MEFLG]. 3♂3♀.Antioquia. Santa Fe de Antioquia. Cotove. (06°32'06”N 75°48'52”W) 519 m. 7-Mar-2011. A. M. Vélez [MEFLG]. 1♂. Caldas. Anserma. el 41 - HDA Canoas. (05°10'42.079"N 75°40'43.547"W) 810 m. 29-Sep-2010. M. A. Marín [MEFLG]; PANAMA. 1♂1♀.Los Santos. Cerro Canajagua. 600 m. 24-Jan-1976. [USNM]. 1♂. Chiriquí. Bugaba. Jul. [USNM]; COSTA RICA. 1♀. Espata. 900 m. Oct-1906. [USNM]. 1♂. Escobal. Dec. [USNM]; NICARAGUA. 1♀. Granada. Nandaime. 62 m. 24-Jun-2011. J. M. Maes [MEL]; HONDURAS. 1♂1♀. Cortés. San Pedro Sula (Bella Vista Forest). 24-Jul-1972. [USNM]; MEXICO. 1♀. Chiapas. Bombana. 1-Aug-1975. [USNM]. 1♂. Guerrero. Aug-1916. [USNM].


Cissia similis Syntype: GUATEMALA. 1♂. (Figs. 2E, F). Centr Valleys. [NHM].
NICARAGUA. 1♀. Rivas. Cardanas. 50 m. 2-Mar-2009. J. M. Maes [MEL]. MEXICO. 1♀. Oaxaca. Landelaria. 500 m. 3-Sep-1982. [MEL]. 1♀. Veracruz. Catemaco. 23-Sep-1972. [USNM].
Results and Discussion
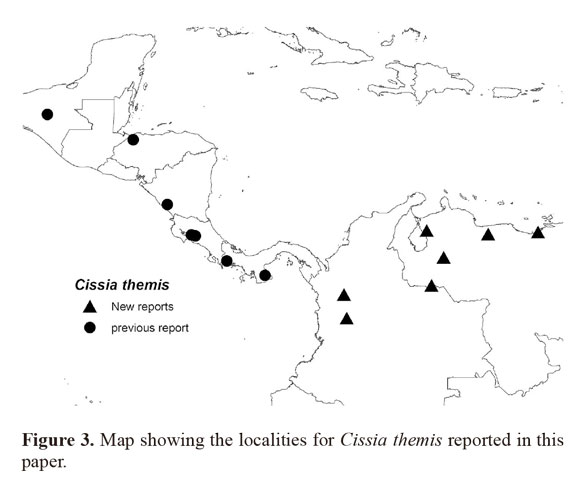
All the localities reported here for C. themis are shown in Figure 3. Six new localities are all from the foothills of mountainous districts within Colombia and Venezuela (Venezuela: Zulia, Barinas, Sucre, Aragua; Colombia: Aracua, Antioquia, Caldas) which is the region where Viloria et al. (2010) report that the satyrine subtribe Pronophilina shows faunal affinities. Two Colombian localities are both associated with the Andes (Cordillera Central and Cordillera Oriental), while Venezuelan localities are also at the base of huge mountain ranges thought to receive influence from the Andes, the Cordillera de Mérida and to a lesser extent, the Sierra de Turimiquire (Viloria et al. 2010). One of the Venezuelan localities, El Bosque de Caimital, is an isolated dense, mature secondary forest (A. Neild, pers. comm.), but with seasonally semi-deciduous vegetation along its four borders; on the other hand, El Rincon is a fragmented dry semi-deciduous forest. The two Colombian localities also belong to a dry semi-deciduous forest. Satyrine species are appropriate indicators of floristic patterns and are reported to show strong affinities with particular forest types (Kremen 1994; DeVries et al. 1997), so it is possible that this species is a typical resident of dry forests below 1000m, and possibly more specifically of those that are rather fragmented and human-dominated. In the Neotropical region, humid/cloud forests that hold a rich biodiversity merit special attention from entomologists (Mittermeier 1988; McNeely et al. 1990), but dry forests are usually given little attention. This probably explains why this species has eluded recognition in Colombia and Venezuela, and also alerts us to the necessity of further research and field work in this type of habitat. Although not yet recorded from Western Ecuador (K.Willmott, pers. comm.), it is very possible that this species is found there given the strong presence of other Central American elements (e.g. Morpho amathonte Deyrolle,1860). It is interesting to note that C. themis was found on the eastern slopes of the Andes (Cordillera Oriental), Cordillera de Mérida and at the far eastern base of the Sierra de Turimiquire which are not the regions where elements of the Central American fauna are usually found. However given this species occurrence in Trinidad these distributional records are not surprising especially since El Rincon is located within a biogeographical region which shows strong affinities with the island of Trinidad (Neild 1996).
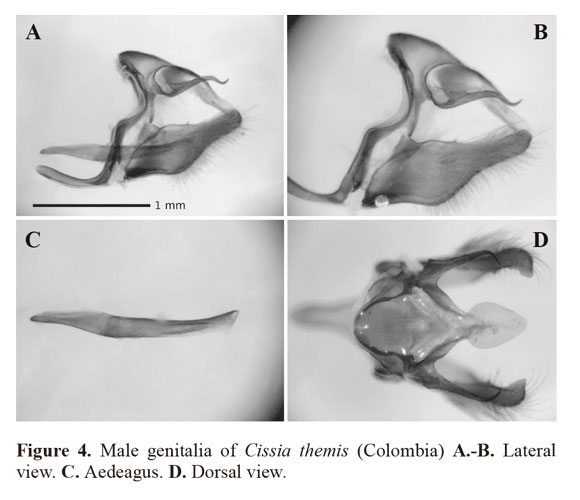
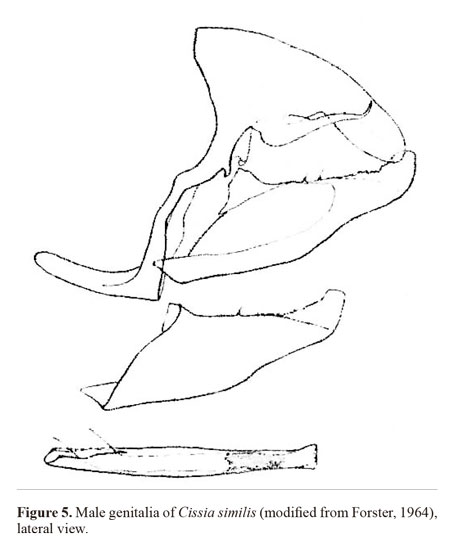
The main characters which identify C. themis are the two double-pupilled large ocelli (in cells s2 and s5), the two smaller double silver-pupilled ocelli between them on the ventral hindwing and ventrally 1.5mm of the antennae from the anterior end is black, a character not seen in sibling species. The original description clearly refers to a sixth hind wing “ocellus” which is the tiny silvery spot at the anal margin between the postmedian and median brown lines, visible in Fig. 2B. Note that this sixth “ocellus” is a variable criteria, but this could be a diagnostic character when the specimen exhibits it. In the MEN, the MUSM and the MEFLG this species was found to be misidentified as C. similis (as in Marín and Uribe 2009)which is distinguished from C. themis by its single (and blind) posterior ocellus and a lack of silver pupilled ocelli on the ventral hindwing. This misidentification is also seen in DeVries (1987) which may cause misidentification of this species elsewhere.Examining C. themis in public and private collections, the ground coloration of the ventral surface appears to be variable within this species. It shows pale brown/ochre to dark brown/ochre which seems likely to be a variation within individuals rather than within geographical distributions. Despite this variation seen in the ground coloration, the double-pupilled ocelli and silver patches appear to be rather stable but we noticed that some specimens do not clearly exhibit this character. In this case, identification could be confirmed by examining the male genitalia (Fig. 4). The genital structure of this species closely resembles that of C. similis (Fig. 5) but differs as follows: in the lateral view of the genitalia, the uncus is narrower and elongate; the tegumen is smaller; the aedeagus is slightly curved downward and convex at the middle section and flattens towards the anterior below the antero-dorsally open tip, posterior slightly curved upward and tapered, opens posteriorly; the distal half of the valvae are rather rectangular and not convex at the apex.
Acknowledgements
We thank Brian Harris (USNM), Jean Michel Maes (MEL), David Lees (NHM) and Gerardo Lamas (MUSM) who gave us access to the satyrine collections in their care, Keith Willmott (MGCL) and Maria Delia Escobar (Venezuela) for their information. We are grateful to Tropical Andean Butterfly Diversity Project funded by the Darwin Initiative who funded digitisation of the images. We acknowledges the Fundación BBVA (MARIPOSA proyect: BIOCON08_021). We are also grateful to Andrew Neild (UK) and Delano Lewis (Jamaica/USA) for their helpful comments, checking the English, and peer reviewing this manuscript.
Literature cited
ACKERY, P. R.; DE JONG. R.; VANE-WRIGHT, R. I. 1998. The butterflies: Hedyloidea, Hesperioidea and Papilionoidea. pp. 263-300. In: Kristensen, N. P. (Ed.). Lepidoptera: moths and butterflies. 1. Evolution, systematics and biogeography. Handbook of Zoology, vol. IV. Walter de Gruyter, Berlin, Part 35. 491 p. [ Links ]
BARCANT, M. 1970. Butterflies of Trinidad and Tobago. Collins, London. 314 p. [ Links ]
BUTLER, A. G. 1867. Descriptions of some new species of Satyridae belonging to the genus Euptychia. Proceedings of the Zoological Society of London 1867 (1): 104-110, pl. 11-12. [ Links ]
DEVRIES, P. J. 1987. The butterflies of Costa Rica and Their Natural History. Vol.1. Princeton, Princeton University Press. 288 p. [ Links ]
DEVRIES, P. J. 1994. Patterns of butterfly diversity and promising topics in natural history and ecology. pp. 187-194. In: MacDade, L.; Bawa, K. S;, Hartshorn, G.; Hespenheide, H. (Eds.). La Selva: ecology and natural history of a neotropical rainforest. Chicago, University of Chicago Press. 493 p. [ Links ]
DEVRIES, P. J.; MURRAY, D.; LANDE, R. 1997. Species diversity in vertical, horizontal and temporal dimensions of fruit-feeding butterfly community in an Ecuador rainforest. Biological Journal of the Linnean Society 62: 343-364. [ Links ]
FORSTER, V. W. 1964. Beitrage zur Kenntnis der Insecktenfauna Boliviens XIX. Lepidoptera 3, satyridae. Veroffentlichungen der Zoologischen Staatssammlung Munchen. (Munich) 8: 51-35, pl 27-35, 264 f. [ Links ]
HUERTAS, B.; RIOS, C.; LECROM, J. F. 2009. A New Species of Splendeuptychia from the Magdalena Vallay in Colombia (Lepidoptera: Nymphalidae: Satyrinae). Zootaxa 2014: 51-58. [ Links ]
KLOTS, A. B. 1956. Lepidoptera. pp. 97-110. In: Tuxen, S. L. (Ed.). Taxonomists' Glossary of Genitalia in Insects. Munksgaard, Copenhagen, Denmark. 284 p. [ Links ]
KREMEN, C. 1994. Biological inventory using target taxa: a case study of the butterflies of Madagascar. Ecological Applications 4: 407-422. [ Links ]
MARÍN, M. A.; URIBE, S. I. 2009. Actualización sobre Euptychiina (Lepidoptera: Satyrinae) representadas en la colección del Museo Entomológico Francisco Luis Gallego. Boletín del Museo Entomológico Francisco Luis Gallego 1 (2): 22-32. [ Links ]
MARÍN, M. A.; PEÑA, C.; FREITAS, A. V. L.; WAHLBERG, N.; URIBE, S. I. 2011. From the phylogeny of the Satyrinae butterflies to the systematics of Euptychiina (Lepidoptera: Nymphalidae): History, Progress and Prospects. Neotropical Entomology 40 (1): 1-13. [ Links ]
MCNEELY, J. A.; MILLER, K. R.; REID, W. V.; MITTERMEIER, R. A.; WERNER, T. B. 1990. Conserving the World's Biological Diversity. Gland, IUCN. viii + 265 p. [ Links ]
MITTERMEIER, R. A. 1988. Primate Diversity and the tropical forest: case studies from Brazil and Madagascar and the importance of megadiversity countries. pp. 145-154. In: Wilson, E. O.; Peter, F. M. (Eds.). Biodiversity. Washington, National Academy Press. 521 p. [ Links ]
MONTERO-RAMÍREZ, J. J.; GONZÁLEZ-MAYA, J. J. 2009. Lepidoptera, Nymphalidae, Satyrinae, Cissia terrestris: First record from Costa Rica. Check List 5 (1): 083-085. [ Links ]
NEILD, A. F. E. 1996. The butterflies of Venezuela. part1: Nymphalidae (Limenitidinae, Apaturinae, Charaxinae). A comprehensive guide to the identification of adult Nymphalidae, Papilionidae, and Pieridae. Greenwich, London, Meridian Publications. 144 p. [ Links ]
PEÑA, C.; LAMAS, G. 2005. Revision of the butterfly genus Forsterinaria Gray, 1973 (Lepidoptera: Nymphalidae, Satyrinae). Revista Peruana de Biología12: 5-48. [ Links ]
SINGER, M. C.; EHRLICH, P. R. 1991. Host Specialization of Satyrine Butterflies, and their Responses to Habitat Fragmentation in Trinidad. Journal of Research on the Lepidoptera30(3-4): 248-256. [ Links ]
VILORIA, A. L.; PYRCZ, T. W.; ORELLANA, A. 2010. A survey of the Neotropical montane butterflies of the subtribe Pronophilina (Lepidoptera, Nymphalidae) in the Venezuelan Cordillera de la Costa. Zootaxa 2622: 1-14. [ Links ]
