










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.39 no.1 Bogotá Jan./June 2013
Mielato de Stigmacoccus asper (Hemiptera: Stigmacoccidae):recurso melífero de bosques de roble en Colombia
Honeydew of Stigmacoccus asper (Hemiptera: Stigmacoccidae): a bee-honey resource in oak forests of Colombia
Fermín J. Chamorro1,2, Guiomar Nates-Parra1,3 y Takumasa Kondo4
1 Laboratorio de Investigaciones en Abejas (LABUN ), Departamento de Biología, Universidad Nacional de Colombia, sede Bogotá. Carrera 30 No. 45-03, Edificio 421, Bogotá, D.C., Colombia.
2 Licenciado en Biología, Estudiante de Posgrado. fjchamorrog@unal.edu.co. Autor para correspondencia.
3 M. Sc. Profesora Titular. mgnatesp@unal.edu.co.
4 Ph. D. Investigador. Corporación Colombiana de Investigación Agropecuaria (COR POI CA), Centro de Investigación Palmira, km 1, Contiguo al Penal, Palmira, Valle, Colombia. takumasa.kondo@gmail.com.
Recibido: 30-nov-2012 - Aceptado: 30-may-2013
Resumen: Se reporta el mielato (excremento liquido azucarado) del insecto escama Stigmacoccus asper (Hemiptera: Stigmacoccidae) como un recurso importante para la producción de miel de abeja. S. asper se encontró en bosques de roble Quercus humboldtii (Fagaceae) de once localidades, ubicadas entre los 2.400 y 2.800 msnm, en los departamentos de Boyacá y Santander, en la Cordillera Oriental de Colombia. Los bosques de roble donde se encontró S. asper corresponden a fragmentos altamente intervenidos y los árboles con mielato son abundantes en el borde de los fragmentos. Con base en un estudio palinológico se estimó que el mielato es tan importante como el néctar floral en la producción de miel de las zonas estudiadas, y en algunas localidades y épocas del año se produce miel de mielato. Como elementos indicadores de mielato se identificaron hifas, esporas (asociadas con hongo fuliginoso) y polen de roble encontrados en las muestras de miel, y se determinó su relación con tipos polínicos de plantas nectaríferas, permitiendo clasificar las mieles en tres tipos: mieles de mielato, mieles florales y mezclas. Se identificaron 67 tipos polínicos de plantas nectaríferas o potencialmente nectaríferas. Los apicultores reportaron una mayor producción de mielato en épocas secas y las mieles obtenidas presentan un color oscuro y sabor a caramelo intenso. El mielato de S. asper en los bosques de roble se presenta como un nuevo recurso melífero que puede ser la base para la obtención de la miel de mielato, un producto forestal no maderable novedoso para el país, con beneficios económicos y especialmente ambientales.
Palabras clave: Apicultura. Miel de mielato. Producto forestal no maderable. Stigmacoccus.
Abstract: The honeydew produced by the scale insect Stigmacoccus asper (Hemiptera: Stigmacoccidae) is recorded an important source for honey production by honey bees. S. asper was found on oak trees, Quercus humboldtii (Fagaceae) at eleven sites in the states of Boyacá and Santander, at 2,400-2,800 masl, in the Oriental Cordillera of Colombia. The oak forests where S. asper was found are highly disturbed and fragmented and trees with honeydew are most abundant on the edge of the fragments. Based on a palynological study, we estimated that honeydew is as important as floral nectar in honey production in the areas studied. In some localities and seasons, honeydew is the main ingredient of local honey. Presence of honeydew in honey was estimated by the amount of hyphae, spores (associated with sooty molds) and oak pollen found in it. Their abundance was compared with pollen from nectar-producing plants, thus allowing the classification of sampled honeys into three types: honeydew honey, floral honey and mixed honey. 67 types of nectar-producing or potentially nectar-producing plants were identified. Beekeepers reported increased production of scale insect honeydew during dry periods and described the honey produced from the honeydew of S. asper as being dark and with an intense caramel flavor. Honeydew from oak forests is suggested as a resource for honey, which has the potential to become a new apicultural product in Colombia. Furthermore, honeydew honey is an economically important non-timber product, and is also environmentally important because it promotes the conservation of native oak forests.
Key words: Beekeeping. Honeydew honey. Non-timber forest product. Stigmacoccus.
Introducción
Aunque la mayor parte de la miel producida en el mundo procede del néctar floral (mieles florales: monoflorales, multiflorales), las abejas pueden elaborarla a partir de otras fuentes de azúcares como el mielato o miel de rocío (honeydew en inglés) y en algunos países la miel de mielato (honeydew honey) representa un porcentaje importante del total de su producción, incluso por encima de las mieles florales (Santas 1983; Kunkel 1997; Gounari 2003). El mielato es el exceso de líquido azucarado que eliminan los insectos fitófagos del orden Hemiptera, suborden Sternorrhyncha, después de tomar grandes cantidades de savia, con el fin de satisfacer sus necesidades nutricionales (Malumphy 1997). Este recurso es recolectado y tratado por las abejas igual que el néctar, sufriendo los mismos procesos enzimáticos y de almacenamiento (Barth 1970; Maurizio 1975).
Sin embargo para producir miel de mielato es necesario que: 1) la eliminación de mielato por el insecto sea alta por unidad de tiempo; 2) las poblaciones de insectos sean altas en una localidad dada, y 3) estas poblaciones sean accesibles para las abejas y para los apicultores (Kunkel 1997). Las abejas recolectan mielato cuando lo encuentran por casualidad, en su búsqueda de néctar y/o agua (Kunkel 1997). Por tal razón, en varios países del mundo (p.ej., Bulgaria, Francia, Grecia, Nueva Zelanda, Turquía), los apicultores ubican sus colmenas en bosques con abundante oferta de mielato, para facilitar que las abejas encuentren el recurso y así obtener cosechas de miel (Kunkel 1997). En Grecia, se ha calculado que alrededor de 15.000 toneladas de miel de mielato (60% de la producción nacional de miel) son colectadas cada año, principalmente derivada del mielato del insecto escama Marchalina hellenica (Gennadius, 1883) (Hemiptera: Marchalinidae) que se alimenta de diferentes especies de pino, Pinus spp. (Pinaceae)(Santas 1983; Gounari 2003). En las montañas de California y Oregon, EE.UU., se produce la "miel de cedro blanco" (white cedar honey), producida por las abejas a partir del mielato de la escama Xylococculus macrocarpae (Coleman, 1908) (Hemiptera: Xylococcidae) que se encuentra especialmente en la corteza de Calocedrus decurrens (Torr.) Florin (Cupressaceae) (Kunkel 1997). También, en los bosques de Nothofagus (Fagaceae) de Nueva Zelanda se produce miel aprovechando el mielato eliminado por Ultracoelostoma assimile (Maskell, 1890) (Hemiptera: Coelostomidiidae), la cual se exporta a Europa desde 1970 y se considera una miel de alta calidad por sus propiedades antimicrobianas y antioxidantes (Crozier 1981). Por otra parte, en Vosges, Francia, la miel de mielato producida a partir del mielato eliminado por diferentes especies de áfidos del género Cinara Curtis (Hemiptera: Aphididae) que se alimentan de árboles de abeto, Abies pectinata Gilib. y A. alba Mill. (Pinaceae), se encuentra amparada por una "Indicación Geográfica Protegida" y los apicultores transportan sus colmenas a los bosques que albergan estos insectos, basados en sistemas de información que predicen la aparición del mielato y las intensidades de sus flujos (Morlot 2008).
En Colombia, a pesar de que la norma técnica para miel de abejas (NTC 1273, ICONTEC 2007) incluye una definición de la miel de mielato, en ninguna región se habían registrado producciones de miel a partir de este recurso. Sin embargo, en el transcurso de una serie de muestreos de flora asociada a la actividad apícola realizados en el año 2008, en los municipios de Caldas y Chiquinquirá, en el occidente del departamento de Boyacá, se registró la presencia del insecto productor de mielato Stigmacoccus asper Hempel, 1900 (Hemiptera: Stigmacoccidae) en bosques de roble Quercus humboldtii Bonpl. (Fagaceae); el uso del mielato de este insecto por abejas melíferas y el reconocimiento por parte de los apicultores como recurso importante para la producción de miel. Estas observaciones hacen parte de los resultados de este trabajo y fueron el motivo para determinar si la presencia de S. asper se limitaba a las zonas identificadas o se extendía a otras localidades de la Cordillera Oriental, considerando que en esta región se encuentran los remanentes más importantes de bosques de roble en Colombia (Avella y Cárdenas 2010) y la apicultura es una actividad productiva más o menos difundida en esta región (Martínez 2006) (Fig. 1). Igualmente, con el objetivo de estimar la contribución del mielato a la composición de la miel y así evaluar su importancia como recurso alternativo para la producción de miel en la Cordillera Oriental, se realizaron análisis palinológicos de mieles producidas en zonas donde se identificaron poblaciones de S. asper.
Materiales y métodos
El estudio se realizó en tres zonas de producción apícola, ubicadas en la vertiente occidental de la Cordillera Oriental de Colombia, en los departamentos de Boyacá, Cundinamarca y Santander. En estas zonas como en otras regiones del país, los apicultores manejan abejas melíferas africanizadas, descendientes de Apis mellifera scutellata Lepeletier, 1836 (Hymenoptera: Apidae) (Nates-Parra 2006). La primera zona de producción apícola se localiza al sur del departamento de Cundinamarca, en la cuenca del río Sumapaz, la segunda entre los departamentos de Cundinamarca y Boyacá, en la cuenca alta del río Suárez y de la Laguna de Fúquene y la tercera al norte del departamento de Boyacá y sur oriente del departamento de Santander, en la cuenca media del río Chicamocha (Fig.1) (Tabla 1).
Entre agosto de 2009 y enero de 2011 se estudiaron las tres zonas de producción apícola con el fin de identificar apiarios con bosques de roble en su área de influencia y la presencia de insectos productores de mielato en los bosques. Para esto se visitaron fincas donde hay apiarios y en las que se identificaron bosques de roble, se realizaron recorridos tanto dentro como fuera de estos, examinando los árboles de roble y determinando la presencia de insectos productores de mielato. Un árbol se consideró infestado si el filamento anal de al menos un espécimen de S. asper era visible y/o se observaba el crecimiento de fumaginas (Fig. 2). Las fumaginas son hongos superficiales que se caracterizan por tener micelios oscuros y solamente se pueden desarrollar en presencia de grandes cantidades de mielato, que les sirve como medio de crecimiento (Barth 1970; Mibey 1997). En las localidades donde se determinó la presencia de insectos productores de mielato, se tomaron muestras de estos, extrayéndolos con una navaja y depositándolos en tubos con alcohol al 70% para su preservación. Posteriormente en laboratorio los insectos se prepararon y montaron en láminas permanentes (por T. Kondo) siguiendo el método de Williams y Granara de Willink (1992) para Pseudococcidae. La identificación taxonómica de S. asper se realizó usando la clave de Hodgson et al. (2007). Las láminas se depositaron en el laboratorio de entomología de Corpoica, Centro de Investigación Palmira, km 1, contiguo al Penal, Palmira, Valle del Cauca, Colombia.

De las localidades donde se registraron apiarios con bosques de roble y oferta de mielato se solicitó a los apicultores muestras de miel para evaluar la contribución del mielato a la composición de éstas. Se obtuvieron 24 muestras de miel, con las cuales se elaboraron preparaciones microscópicas al natural (sin acetólisis) y se realizaron análisis palinológicos. Las muestras de miel son una mezcla de mieles en diferentes partes de la colmena. El estudio microscópico de los componentes del sedimento obtenido de una miel permite no sólo identificar tipos polínicos asociados a plantas nectaríferas o potencialmente nectaríferas sino también detectar y cuantificar la presencia de elementos indicadores de mielato, como hifas y esporas de las fumaginas, que también son llevadas por las abejas cuando recolectan mielato. De este modo es posible determinar cuáles han sido las fuentes de azúcares utilizadas por las abejas para la producción de miel. De cada muestra de miel se utilizó 10 g, los cuales se disolvieron en agua destilada caliente por partes (1: 4 ml y 2: 3 ml, cada parte se disolvió en 15 ml de agua destilada), para facilitar la concentración de las partículas y elementos contenidos en esta. Entre cada disolución se centrifugó a 3000 rpm por 10 minutos. Del sedimento resultante se realizó un montaje de dos alícuotas por lámina (dos áreas de conteo por lámina) con gelatina glicerinada y selladas con parafina. Se utilizó un microscopio Leica DM LS2 para la identificación de tipos polínicos y detección de elementos fúngicos de las fumaginas, así como para hacer conteos de estos. Se tomó como criterio de conteo un mínimo de 300 granos de polen por muestra. El reconocimiento taxonómico de los tipos polínicos hasta nivel de especie, género y/o familia, se hizo con base en catálogos de polen (Roubik y Moreno 1991; Velásquez 1999), así como por comparación directa con la palinoteca del Laboratorio de Investigaciones en Abejas (LABUN) de la Universidad Nacional de Colombia, sede Bogotá, Bogotá, D.C., Colombia.
Posteriormente se procedió a hacer un análisis cualitativo de los conteos siguiendo los criterios propuestos por Louveaux et al. (1978) y Barth (2005). Los tipos polínicos identificados se clasificaron en tipos polínicos de plantas nectaríferas y tipos polínicos de plantas no nectaríferas (poliníferas y/o anemófilas), según lo reportado en la literatura sobre aspectos de biología floral y polinización para las familias, géneros o especies identificados en las muestras. Con base en lo anterior se excluyeron de los conteos los tipos polínicos de plantas no nectaríferas y se calcularon abundancias relativas (%) de los elementos indicadores de mielato (elementos fúngicos de las fumaginas) y de los tipos polínicos de plantas nectaríferas. Las mieles se clasificaron como miel de mielato o miel floral cuando el aporte de mielato o de néctar fue mayor o igual a 70%, respectivamente. Las muestras con aportes de néctar o mielato mayor de 30% y menor de 70%, respectivamente, se clasificaron como mezclas. Este rango se definió con base en lo reportado en la literatura cuando se combinan criterios palinológicos y fisicoquímicos para estimar el porcentaje de mielato en una muestra de miel (Soria et al. 2004, 2005). También se calculó la abundancia relativa de cada tipo polínico de planta nectarífera con respecto al total contado por muestra para determinar la dominancia o no de alguna especie de planta en el aporte de néctar. Para tal fin se siguieron los términos propuestos por Louveaux et al. (1978): Dominante > 45%, Secundario ≥ 16 ≤ 45%, Aislado importante ≥ 3 < 16%, Aislado < 3% y se definió el tipo de aporte de néctar en términos de: monofloral cuando un tipo polínico fue dominante, bifloral cuando dos tipos polínicos presentaron frecuencia relativa secundaria y los dos sumaron más de 45%, oligofloral cuando dos o más tipos polínicos con frecuencia relativa secundaria pertenecían a una misma familia, y multifloral cuando se presentó un tipo polínico secundario y más de cuatro tipos polínicos aislados importantes o aislados o cuando ningún tipo polínico fue secundario y los tipos polínicos identificados fueron aislados importantes o aislados.
Resultados
Áreas de producción apícola con bosques de roble y oferta de mielato. Se registraron 17 apiarios con bosques de roble en su zona de influencia (Tabla 1), los cuales se encuentran ubicados en fincas con predominio de actividades agropecuarias, donde además de bosques de roble, se observaron otros tipos de coberturas vegetales generales asociadas: matorrales, pastizales, cultivos y plantaciones forestales. Los bosques de roble en la mayoría de las localidades corresponden a fragmentos altamente intervenidos, similares a una plantación forestal, sin sotobosque definido y con evidencia de pastoreo. Los apicultores tienen sus apiarios en los matorrales que limitan con los fragmentos de bosques de roble, con 10 colmenas en promedio y dedicados a la producción de miel, polen o a los dos productos. En general, según los apicultores del área de estudio, la producción de miel (12 kg por colmena) se da en los primeros meses del año y la de polen (18 kg por colmena) durante la mayor parte del año, siendo en algunos apiarios mayor que la de miel.
Se identificaron poblaciones de S. asper en los bosques de roble de 11 localidades: nueve de las 17 donde hay actividad apícola y dos donde no hay apiario instalado aunque los apicultores se encuentran negociando su instalación con el propietario de la finca (Tabla 1). Los apicultores manifiestan que hay mayor producción de mielato en épocas secas (enero y julio), a los cuales les siguen producciones de miel caracterizadas por su color oscuro y sabor a caramelo intenso, que denominan "miel de roble". Antes de la realización de este trabajo, los apicultores no conocían que el mielato que recolectan sus abejas es producido por insectos escama, pero si habían observado a sus abejas recolectando activamente este recurso y que la miel producida posteriormente, presentaba características diferentes de las otras mieles.
Stigmacoccus asper en los bosques de roble. Este insecto escama se ubica debajo de la corteza de los árboles de roble Q. humboldtii y elimina mielato en forma de gotas a través de un filamento ceroso hueco conectado al ano (Fig. 2). El mielato cae sobre la corteza de los árboles y en el suelo, superficies donde crecen fumaginas, que en algunos árboles se observaban cubriendo completamente el tronco y las ramas (Figs. 2A y C). Los árboles infestados con S. asper se encontraron tanto en el interior como en el borde de los fragmentos de bosque, así como en árboles de roble aislados en los potreros. Aunque no se realizó una cuantificación de los niveles de infestación de S. asper en las diferentes zonas, la observación de árboles con tronco y ramas completamente cubiertos de fumaginas principalmente en el borde de los fragmentos, sugieren que hay un mayor nivel de infestación en estas zonas. En las localidades de Quipe (municipio de Caldas, occidente de Boyacá), Peñuela (municipio de San Mateo, norte de Boyacá) y Robles (municipio de Enciso, Sur Oriente de Santander), se observó una mayor cantidad de robles infestados por S. asper. No se encontraron poblaciones de S. asper en Cundinamarca.
En todas las localidades visitadas donde se encontró S.asper, se observó la especie A.mellifera recolectando activamente mielato, en vuelo, tomando las gotas que penden de los filamentos anales pero principalmente recolectando el mielato acumulado sobre el tronco de los árboles (Fig. 2F). Durante estas observaciones también se registró el consumo de mielato por otras especies animales: colibríes, avispas y abejas sin aguijón Parapartamona zonata (Smith, 1854)(Hymenoptera: Apidae: Meliponini) (Fig. 2G). Las abejas sin aguijón se encontraron al occidente de Boyacá, en las veredas Laja (municipio de Buenavista) y Monte Pinal (municipio de Pauna), localidades donde la actividad apícola es casi nula.
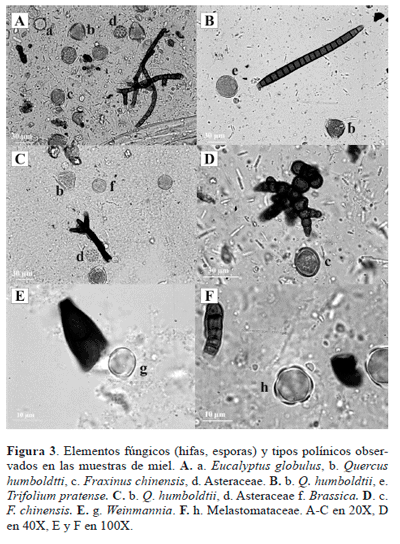
Contribución del mielato a la producción de miel. La cantidad de elementos indicadores de mielato (hifas, esporas) (Fig. 3) encontrados en las muestras de miel con relación a la de tipos polínicos de plantas nectaríferas, permitió clasificar 2 muestras como mieles de mielato, 10 muestras como mieles florales y 12 muestras como mezclas de miel de mielato y miel floral (Tabla 2). Con respecto al aporte de néctar, en las 24 muestras de miel analizadas, se identificaron 67 tipos polínicos de plantas nectaríferas o potencialmente nectaríferas (tipos polínicos identificados a nivel de familia e indeterminados), de los cuales 27 tipos polínicos aportaron más del 90% de los recursos (Tabla 3). Sin embargo, en términos generales, el polen de tipo Brassica (Brassicaceae), Eucalyptus globulus Labill. (Myrtaceae), Weinmannia (Cunoniaceae), Asteraceae y Melastomataceae fueron las principales fuentes de néctar. También se encontró en menor cantidad polen de Ericaceae, Escallonia pendula (Ruiz & Pav.) Pers., E. paniculata (Ruiz & Pav.) Roem. & Schult. (Escalloniaceae), Gaiadendron punctatum (Ruiz & Pav.) G. Don (Loranthaceae), Hesperomeles y Rubus (Rosaceae), Hypochaeris radicata L. (Asteraceae), Oreopanax (Araliaceae), Trifolium pratense L. y T. repens L. (Fabaceae) entre otras (Tablas 1 y 2).
Discusión
Stigmacoccus asper es una especie neotropical, al igual que todas las especies del género, de la cual ya existen registros para Colombia, pero asociada a árboles de guamo Inga sp. (Fabaceae) en zonas cafeteras de Villavicencio, departamento del Meta (Hodgson et al. 2007). También se ha registrado su presencia en Brasil, Panamá y Venezuela, igualmente en árboles de la familia Fabaceae (Cassia sp.e Inga spp.) (Hodgson et al. 2007). Sin embargo, no se había registrado como parásito de Q. humboltdtii, aunque si se conocía la relación de la especie Stigmacoccus garmilleri Foldi, 1995 (Hemiptera: Stigmacoccidae) con seis especies de Quercus en paisajes fragmentados de Veracruz, México (Gamper et al. 2011).
Así como otros insectos productores de mielato, S. asper utiliza sus estiletes para perforar los vasos del floema y succionar savia (Figs. 2D y E), proceso alimenticio que tiene como fin captar aminoácidos para la síntesis de proteínas (Malumphy 1997). Debido a que la savia del floema presenta bajas concentraciones de aminoácidos y altas concentraciones de azúcares, los insectos deben ingerir grandes cantidades de alimento para satisfacer sus necesidades nutricionales y para crecer. Su canal alimentario se ha modificado para formar una "cámara filtrante", que facilita la concentración de los aminoácidos para su absorción y filtra el exceso de agua y azúcares tomados del floema, pasando directamente al recto para su eliminación a través del ano (Malumphy 1997). Este exceso de agua y azúcares es el mielato, que en S. asper pasa al filamento ceroso hueco conectado a su ano para su eliminación final (Fig. 2D). El filamento es secretado por un anillo de poros tuberculados que rodean el ano del estado de quiste (Hodgson et al. 2007). Esta forma de eliminación final del mielato también se presenta en otras especies del género Stigmacoccus, así como en los géneros Ultracoelostoma Cockerell (Malumphy 1997) y Xylococculus Morrison (T. Kondo, obs. pers.). La eliminación de mielato en S. asper se produce durante el primer instar (etapa de gateador) y los estados de quiste, en ambos sexos, pero no en los adultos porque estos no tienen partes bucales (Hodgson et al. 2007).
Stigmacoccus asper se encontró tanto al interior como en el borde de fragmentos de bosques de roble y también en arboles aislados en los potreros, aunque se observaron más árboles infestados por este insecto en el borde de los fragmentos. Este patrón de distribución y abundancia es similar al descrito para S. garmilleri en México (Gamper et al. 2011) y para U. assimile en Nueva Zelanda (Crozier 1981). En estos lugares se ha encontrado que altos niveles de infestación en el borde de los bosques y en árboles por fuera de estos, se relacionan positivamente con aumentos en temperatura provocados por la exposición a la luz solar, que en los bosques de Quercus de México tiene su origen en la continua remoción de madera de los bosques. Puede pensarse también que la distribución ‘natural' o del pasado de estos insectos, cuando los bosques no estaban fragmentados, estaba restringida a las ramas altas de los robles. Por este motivo, la interacción entre S. garmilleri y Quercus spp., se considera una característica de bosques montanos altamente fragmentados (Gamper y Koptur 2010; Gamper et al. 2011). Igualmente para bosques templados en Inglaterra (Ozanne et al. 2000) y bosques tropicales amazónicos en Brasil (Fowler et al. 1993), se ha registrado que los insectos escama y los áfidos reaccionan positivamente a la fragmentación, observándose un incremento de sus poblaciones en árboles aislados y en el borde de los bosques. De acuerdo con lo anterior, es bastante probable que S. asper se encuentre en bajas densidades en los bosques de roble, pero debido a la fragmentación y alteración de estos ecosistemas por causas antrópicas como la explotación maderera del roble (Avella y Cárdenas 2010), se han creado condiciones ambientales (p.ej., mayor exposición solar, homogeneidad de los bosques, disminución de enemigos naturales) que podrían estar favoreciendo el aumento de sus poblaciones. Las presiones antrópicas, principalmente la apertura de nuevos potreros, aún persisten sobre los bosques de roble (Avella y Cárdenas 2010), como se pudo observar en los bosques de roble de las localidades estudiadas.
El uso del mielato por A. mellifera se conoce ampliamente, tanto en áreas donde son nativas como introducidas (Kunkel 1997), pero el uso por abejas sin aguijón ha sido poco documentado (Lorenzon y Matrangolo 2005; Koch et al. 2011). En ambos casos se considera que las abejas explotan el mielato cuando los recursos nectaríferos son escasos. De igual forma, en un trabajo experimental controlado con la abeja solitaria Osmia bicornis (L., 1758) (Hymenoptera: Megachilidae), se concluyó que la explotación del mielato por estas abejas, depende del tipo del mielato y de la ausencia o escasez de néctar (Konrad et al. 2009). En comparación con el néctar, el mielato se considera una fuente de azúcares inferior, esto debido a su a veces limitada accesibilidad (producto de la evaporación y la presencia de azúcares como rafinosa y melecitosa) pero principalmente a su composición química particular (nuevos olisacáridos sintetizados por los insectos y/o metabolitos secundarios derivados de las plantas) (Konrad et al. 2009). Con relación al mielato de S. asper como fuente de azúcares para animales, solamente se conoce su uso por colibríes Amazilia viridigaster, Chlorostilbon sp. y Metallura sp., en árboles de guamo (Inga sp.) de zonas cafeteras de Villavicencio, Colombia (Köster y Stoewesand, 1973). No obstante, el mielato de otras especies de Stigmacoccus se registra que es consumido por ácaros, abejas melíferas, avispas, hormigas, moscas y aves, siendo estas últimas sus principales consumidores (Hodgson et al. 2007). En bosques secos de República Dominicana, 15 especies de pájaros, migratorios y residentes, han sido observados alimentándose del mielato producido por Stigmacoccus sp., destacándose Dendroica tigrina (Gmelin, 1789) (Passeriformes: Parulidae) que pasa más del 85% de su actividad diaria en el consumo y defensa del mielato (Latta et al. 2001). Igualmente, en los bosques de Quercus de Veracruz, México, alrededor de 25 especies de aves, entre residentes y migratorias, se han observado consumiendo el mielato de S. garmilleri, de las cuales Dendroica coronata auduboni (J.K. Townsend, 1837) (Passeriformes: Parulidae) es el consumidor más activo y dominante, principalmente de árboles con mielato aislado en los potreros (Gamper y Koptur 2010). También en Hidalgo, México, cuatro especies de colibríes se alimentan del mielato de Stigmacoccus sp., presentándose una distribución espacio-temporal del uso del mielato entre las especies, como estrategia de coexistencia y explotación efectiva del recurso (Lara et al. 2011). En general se considera que el mielato eliminado a través de filamentos anales como los de S. asper, puede ser una fuente importante de azúcares para animales, dado su fácil acceso y reconocimiento en campo (gotas brillantes que penden de los filamentos) (Kunkel 1997).
En cuanto al aporte del mielato a la producción de miel, el análisis microscópico de la miel permitió clasificar dos muestras como miel de mielato y 12 como mezclas de un total de 24 muestras. Estos resultados indican que el mielato es tan importante como el néctar en la producción de miel en las zonas estudiadas. Incluso, el aporte de mielato podría ser más alto de lo estimado a través del análisis microscópico, considerando que: 1) así sea en bajas cantidades, en todas las muestras analizadas se identificaron elementos indicadores de mielato y esta es una característica ya definida para mieles de mielato de roble de la región mediterránea de Europa donde los principales insectos proveedores de mielato son Tuberculatus annulatus (Hartig. 1841) y T. borealis (Krzywiec 1971) (Hemiptera: Aphididae) que parasitan diferentes especies de Quercus (Ricciardelli D' Albore 1998); 2) las características organolépticas (color oscuro y sabor a jarabe o caramelo intenso) de la mayoría de las muestras analizadas en este trabajo se han establecido como típicas para mieles de mielato de Europa (Piana 2008); y 3) resultados de la caracterización fisicoquímica de las muestras analizadas en este trabajo, muestran que más del 50% de estas, presentan una conductividad eléctrica superior a 0,8 mS/cm, el parámetro principal utilizado para diferenciar mieles de mielato de mieles florales y más del 35% cumplió con los valores de referencia europeos para mieles de mielato, entre los cuales se encuentran: alto contenido de cenizas, altos valores de pH (> 4,5), altos niveles de oligosacáridos, especialmente melecitosa superior a 0,5 g/100 g y valores positivos de rotación específica (Gamboa-Abril, com. pers.).
Sin embargo, según Barth (1970) la miel de mielato nunca se produce de forma pura y siempre va estar acompañado por porcentajes variables de miel floral, puesto que las abejas al mismo tiempo que recolectan mielato no dejan de visitar flores para recoger néctar. Como se expuso anteriormente, las abejas prefieren el néctar floral ante el mielato y algunas de las principales fuentes de néctar identificadas en este trabajo (Brassica, E. globulus, Weinmannia, Asteraceae y Melastomataceae) se encuentran entre las más utilizadas por las abejas y asociadas a la producción de mieles monoflorales y biflorales en zonas altoandinas (Bogotá et al. 2001; Nates-Parra y Chamorro, obs. pers.). Estas plantas son ampliamente cultivadas (i.e., E. globulus), abundantes y frecuentes en los bosques y matorrales andinos (i.e., Weinmannia) y de rápido crecimiento entre y durante épocas de cultivo, cubriendo áreas importantes de terreno (i.e., Brassica), condiciones que favorecen la concentración de los recursos en un área determinada y su recolección por parte de las abejas. En cuanto a Asteraceae, aunque no fue posible identificar a nivel específico varios de los tipos polínicos de esta familia, es muy probable que estos correspondan a especies típicas de estados sucesionales tempranos que limitan con los bosques, como los géneros Baccharis y Pentacalia, identificados en los espectros polínicos y muy comunes en las diferentes coberturas vegetales alrededor de los apiarios. Finalmente, las plantas de la familia Melastomataceae se consideran en general como plantas poliníferas y no productoras de néctar. No obstante, especies como Miconia ligustrina (Sm.) Trianay M. squamulosa Triana (Melastomataceae) presentes en los bosques y matorrales del área de estudio producen néctar, recurso recolectado de forma frecuente por abejas melíferas y abejorros Bombus hortulanus Friese, 1904 (Hymenoptera: Apidae) (F.J. Chamorro, obs. pers.).
Implicaciones para el manejo y conservación del roble. Los bosques dominados por Q. humboltdtii son ecosistemas que albergan una rica biodiversidad (p.ej., más de 550 especies de plantas vasculares) y prestan importantes servicios ecosistémicos como retención y regulación del flujo hídrico, asociadas a las condiciones de humedad y sombra generadas por las densas copas de estos bosques que permiten la presencia de un gran número de especies de briófitos, líquenes, bromelias, orquídeas y helechos (Avella y Cárdenas 2010). Sin embargo, debido a la extracción intensiva para madera, el roble se ha declarado en estado vulnerable y se ha establecido veda nacional para su aprovechamiento forestal (Salinas y Cárdenas 2007).
Si bien se prohíbe la tala de árboles de roble, también se considera el manejo sostenible y el de los bosques asociados a esta especie, como un mecanismo fundamental para su conservación y un tema prioritario para aportar información en este sentido se relaciona con potenciar y reconocer los usos no maderables de los recursos del bosque (Avella y Cárdenas 2010). Las comunidades locales han incorporado el uso del roble (alimento, colorante, medicinal, entre otros) a sus sistemas de vida, ayudando a consolidar su base económica y/o alimentaria (Ariza et al. 2010). Teniendo en cuenta estos antecedentes y los resultados de este trabajo, la apicultura se presenta como otra alternativa de uso no maderable del roble, a través del aprovechamiento del mielato para la producción de miel. Si se parte de que la fuente primaria de esta miel sería la savia del roble, la miel de mielato de roble podría catalogarse como producto forestal no maderable y un incentivo a la conservación de esta. En los mercados alternativos, los apicultores pueden conseguir un valor agregado para la miel de mielato, que los motivaría a llevar a cabo prácticas de conservación del bosque como parte del manejo apícola. Para que este potencial de uso se pueda desarrollar y no se comprometa la sustentabilidad del recurso y de la misma actividad, y que los mercados funcionen para los productores, se hace necesario profundizar en el conocimiento del mielato de S. asper, particularmente sobre aspectos productivos, ecológicos y de agregación de valor y así planificar el aprovechamiento apícola. Hay que determinar el potencial melífero del mielato, considerando variables temporales y espaciales, lo cual permitirá hacer estimaciones de cuanto más es posible producir y evaluar la capacidad de carga de las zonas o visto de otra forma la cantidad de abejas que se necesitan para explotar el recurso sin comprometer su supervivencia y las de otros consumidores del mielato. Las abejas melíferas podrían competir con la fauna nativa establecida, para quienes el mielato podría ser un recurso alimenticio importante (Hodgson et al. 2007). El mielato de los bosques de roble es consumido por diferentes animales, como P. zonata, especie de abeja sin aguijón endémica de los andes colombianos y ecuatorianos (González y Nates-Parra 1999). Por otra parte, con un mayor conocimiento de los aspectos productivos y ecológicos del mielato, los apicultores podrán orientar sus prácticas de manejo hacia el aseguramiento de la producción de la "miel de roble", cosechando después de ocurrida la oferta de mielato y evitando que se mezcle con el néctar floral. Si bien los apicultores tienen ubicadas sus colmenas cerca de los bosques de roble, lo cual puede facilitar la recolección del mielato por parte de las abejas, se hacen necesarias estas prácticas considerando la oferta de recursos nectaríferos atractivos para las abejas en las zonas de producción.
Además, es importante complementar el conocimiento de la contribución real del mielato a la producción de miel, no solamente en términos de composición sino también en la identificación de propiedades bioactivas heredadas de este recurso, con el fin de agregar un mayor valor al producto y al mismo tiempo a los bosques de roble. Este conocimiento permitiría a los apicultores de la Cordillera Oriental, consolidar una alternativa de diversificación productiva, que actualmente tiene relevancia a nivel comercial. Hasta hace algunos años, las mieles de mielato eran tratadas por igual, pero con la identificación de propiedades especiales (antimicrobianas y antioxidantes), relacionadas con la diversidad de fuentes (árboles e insectos) y de áreas productivas, se está trabajando para incrementar su conocimiento y posicionarlas en el mercado de los productos apícolas. En ese sentido, ya se han realizado dos Simposios Mundiales de Mieles de Mielato (IHC 2008, 2010). La miel de roble de los Andes orientales sería el primer producto de esta naturaleza (miel no floral, miel de mielato) en Colombia y el segundo relacionado a los insectos del género Stigmacoccus. En Brasil, los apicultores obtienen la "miel negra" a partir del mielato de Stigmacoccus paranaensis Foldi, 2006 (Hemiptera: Stigmacoccidae), el cual parasita árboles de la familia Fabaceae (Hodgson et al. 2007). Finalmente, la miel de mielato de roble podría ser comercializada por su origen geográfico, dado que al contener en baja o alta proporción granos de polen de diferentes tipos de plantas nectaríferas, estos podrían servir como indicadores de origen geográfico (Barth 1970).
Conclusiones
El mielato de los bosques de roble constituye una fuente de azúcares para las abejas melíferas tan importante como el néctar en los paisajes andinos fragmentados estudiados en la Cordillera Oriental, lo cual representa una alternativa de diversificación productiva para los apicultores de estas zonas a través de la obtención de un producto apícola novedoso para el país como sería la miel de mielato. Sin embargo, los apicultores deben trabajar en su diferenciación a nivel de manejo productivo, dada la oferta de recursos nectaríferos que pueden resultar más atractivos para las abejas melíferas en las épocas de producción de mielato. Igualmente estos resultados proporcionan una base sólida para evaluar a la apicultura como alternativa de uso sostenible de los bosques de roble y en particular del roble, su especie representativa y hospedero primario de S. asper, especie productora de mielato. En este sentido, la miel de mielato se presenta como un producto forestal no maderable, que al estar directamente asociado al roble puede generar un mayor incentivo de conservación y asimismo facilitaría la disminución de presión sobre estos ecosistemas. Finalmente, estos resultados reafirman la importancia del conocimiento de la biodiversidad asociada a la actividad apícola, que en Colombia adquiere especial relevancia dada la gran diversidad de especies y ecosistemas al interior de las diferentes regiones.
Agradecimientos
A la Universidad Nacional de Colombia y el Ministerio de Agricultura y Desarrollo Rural, que a través del proyecto Diferenciación por origen botánico de mieles y polen de cuatro especies de abejas en Colombia(Contrato No. 2007C3476 118-584/2007), facilitaron los recursos para realizar este trabajo. A las asociaciones de apicultores ASOAPIBOY (Asociación de Apicultores de Boyacá) y ASOAPIGAR (Asociación de Apicultores de la Provincia de García Róvira, Santander), que además de facilitar las muestras y prestar apoyo en campo, fueron los primeros en acercarnos al conocimiento del mielato de los bosques de roble. A Viviana Gamboa, por su colaboración en la fase de campo y obtención de muestras de miel. A la Dra. Penny Gullan (Universidad Nacional de Australia, Camberra) por la identificación inicial de S. asper, y por la revisión del texto del resumen en inglés.
Literatura citada
AVELLA, A.; CÁRDENAS, L. M. 2010. Conservación y uso sostenible de los bosques de roble en el corredor Guantiva-La Rusia-Iguaque, departamentos de Santander y Boyacá. Colombia Forestal 13 (1): 5-26. [ Links ]
ARIZA, W.; HUERTAS, C.; HERNÁNDEZ, A.; GELTVEZ, J.; GONZÁLEZ, J.; LÓPEZ, L. 2010. Caracterización y usos tradicionales de productos forestales no maderables (PFNM) en el Corredor de Conservación Guantiva-La Rusia-Iguaque. Colombia Forestal 13 (1): 117-140. [ Links ]
BARTH, O. M. 1970. Análise microscópica de algumas amostras de mel. 5: melato ("honeydew") em mel de abelhas. Revista Brasileira de Biologia 30: 601-608. [ Links ]
BARTH, O. M. 2005. Análise polínica de mel: avaliação de dados e seu significado. Mensagem Doce 81: 2-6. [ Links ]
BOGOTÁ, R. G.; RANGEL CH., J. O.; JIMÉNEZ, L. C. 2001. Análisis palinológico de mieles de tres localidades de la Sabana de Bogotá. Caldasia 23 (2): 455-465. [ Links ]
CROZIER L. R. 1981. Beech honeydew: forest produce. New Zealand Journal of Forestry 26 (2): 2-29. [ Links ]
FOWLER H. G.; SILVA, C. A.; VENTINCINQUE, E. 1993. Size, taxonomic and biomass distributions of flying insects in central Amazonia: forest edge vs. understory. Revista de Biología Tropical 41: 755-760. [ Links ]
GAMBOA-ABRIL, V. Caracterización de mieles originarias de mielato, recurso valioso del corredor de bosques de robles de Colombia (Tesis de maestría en curso). Posgrado en Ciencia y Tecnología de Alimentos, Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá. Fecha de comunicación: 16 de febrero de 2012. [ Links ]
GAMPER, H. A.; KOPTUR, S. 2010. Honeydew foraging by birds in tropical montane forests and pastures of Mexico. Journal of Tropical Ecology26: 335-341. [ Links ]
GAMPER, H. A.; KOPTUR, S.; GARCÍA-FRANCO, J.; STAPPER, A. P. 2011. Alteration of forest structure modifies the distribution of scale insect, Stigmacoccus garmilleri, in Mexican tropical montane cloud forests. Journal of Insect Science11: 124. Disponible en: http://www.insectscience.org/11.124. [Fecha revisión: 10 febrero 2012] [ Links ].
GONZÁLEZ, V. H.; NATES-PARRA, G. 1999. Sinopsis de Parapartamona (Hymenoptera: Apidae: Meliponini), un género estrictamente andino. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 23 (Suplemento especial): 171-179. [ Links ]
GOUNARI, S. 2003. Seasonal development and ovipositing behavior of Marchalina hellenica (Hemiptera: Margarodidae). Entomologia Hellenica 15: 27-37. [ Links ]
HODGSON, C. J.; GAMPER, H.; BOGO, A.; WATSON, G. 2007. A taxonomic review of the Margarodoid genus Stigmacoccus Hempel (Hemiptera: Sternorrhyncha: Coccoidea: Stigmacoccidae), with some details on their biology. Zootaxa1507: 1-55. [ Links ]
INSTITUTO COLOMBIANO DE NORMAS TÉCNICAS Y CERTIFICACIÓN (ICONTEC). 2007. Norma Técnica Colombia NTC 1273, Miel de abejas. ICONTEC, Bogotá, D.C., Colombia. 21 p. [ Links ]
INTERNATIONAL HONEY COMMISSION (IHC). 2008. 1st World Honeydew Honey Symposium. Program and abstracts. Tzarevo, Bulgaria. 34 p. Disponible en: http://bee-hexagon.net/. [Fecha revisión: 5 mayo 2010] [ Links ].
INTERNATIONAL HONEY COMMISSION (IHC). 2010. 2nd World Symposium on Honeydew Honey. Abstracts. Journal of ApiProduct and ApiMedical Science 2 (3): 102-128. [ Links ]
KOCH, H; CORCORAN, C.; JONKER, M. 2011. Honeydew collecting in Malagasy stingless bees (Hymenoptera: Apidae: Meliponini) and observations on competition with invasive ants. African Entomology 19 (1): 36-41. [ Links ]
KONRAD, R.; WÄCKERS, F. L.; ROMEIS, J.; BABENDREIER, D. 2009. Honeydew feeding in the solitary bee Osmia bicornis as affected by aphid species and nectar availability. Journal of Insect Physiology 55: 1158-1166. [ Links ]
KÖSTER, F.; STOEWESAND, H. 1973. Schildläuse als Honigtaulieferanten für Kolibris und Insekten. Bonner Zoologische Beiträge 24: 15-23. [ Links ]
KUNKEL, H. 1997. Scale insect honeydew as forage for honey production. pp. 291-32. En: Ben-Dov, Y.; Hodgson, C. (Eds.). Soft scale insects, their biology, natural enemies and control. World crop pests, Volume 7A. Elsevier Academic Press, Amsterdam, Países Bajos 452 p. [ Links ]
LARA, C.; MARTÍNEZ-GARCÍA, V.; ORTÍZ-PULIDO R.; BRAVO-CADENA, J.; LORANCA, S.; CÓRDOBA-AGUILAR, A. 2011. Temporal-spatial segregation among hummingbirds foraging on honeydew in a temperate forest in Mexico. Current Zoology 57 (1): 56-62. [ Links ]
LATTA, S. C.; GAMPER, H. A.; TIETZ, J. R. 2001. Revising the convergence hypothesis of avian use of honeydew: evidence from Dominican subtropical dry forest. Oikos 9: 250-259. [ Links ] [ Links ]
LOUVEAUX, J.; MAURIZIO, A.; VORWOHL, G. 1978. Methods of Melissopalynology. Bee World 59: 139-157. [ Links ]
MALUMPHY, C. 1997. Honeydew. pp. 269-274. En: Ben-Dov, Y.; Hodgson, C. (Eds.). Soft scale insects, their biology, natural enemies and control. World crop pests, Volume 7A. Elsevier Academic Press, Amsterdam, Países Bajos. 452 p. [ Links ]
MARTÍNEZ, T. 2006. Diagnóstico de la actividad apícola y de la crianza de abejas en Colombia. Ministerio de Agricultura y Desarrollo Rural, Instituto Interamericano de Cooperación para la Agricultura, Bogotá, D.C. 115 p. [ Links ]
MAURIZIO, A. 1975. How bees make honey. pp. 77-115. En: Crane, E. (Ed.). Honey, a comprehensive survey. Heinemann, Londres, Inglaterra. 608 p. [ Links ]
MIBEY, R. K. 1997. Sooty moulds. pp. 275-290. En: Ben-Dov, Y.; Hodgson, C. (Eds.). Soft scale insects, their biology, natural enemies and control. World crop pests, Volume 7A. Elsevier Academic Press, Amsterdam, Países Bajos. 452 p. [ Links ]
MORLOT, M. 2008. Honeydew and Honeydew Honey in France (abstract). pp 16. En: International Honey Commission (IHC). 1st World Honeydew Honey Symposium. Program and abstracts. Tzarevo, Bulgaria. 34 p. Disponible en: http://bee-hexagon.net. Fecha revisión: 5 mayo 2010] [ Links ].
NATES PARRA, G. 2006. Abejas corbiculadas de Colombia. Facultad de Ciencias, Universidad Nacional de Colombia, Sede Bogotá. 158 p. [ Links ]
OZANNE, C. M. P.; SPEIGHT, M. R.; HAMBLER, C.; EVANS, H. F. 2000. Isolated trees and forest patches: Patterns in canopy arthropod abundance and diversity in Pinus sylvestris (Scots Pine). Forest Ecology and Management 137: 53-63. [ Links ]
PIANA, L. 2008. Sensory characterization of honeydew honeys from different countries (abstract). P 8. En: International Honey Commission (IHC).1st World Honeydew Honey Symposium. Program and abstracts. Tzarevo, Bulgaria. 34 p. Disponible en: http://bee-hexagon.net. [Fecha revisión: 5 mayo 2010] [ Links ].
RICCIARDELLI D'ALBORE, G. 1998. Mediterranean mellissopalinology. Universita degli Studi di Perugia, Italia. 466 p. [ Links ]
ROUBIK, D. W.; MORENO, J. E. 1991. Pollen and Spores of Barro Colorado Island. Monographs in Systematic Botany No. 36. Missouri Botanical Garden, St. Louis, Missouri. 268 p. [ Links ]
SALINAS, N; CÁRDENAS, D. 2007. Roble: Quercus humboldtii. pp. 23-29. En: Cárdenas, D.; Salinas, N. (Eds.). Libro rojo de plantas de Colombia. Volumen 4. Especies maderables amenazadas: Primera parte. Serie de libros rojos de especies amenazadas en Colombia. Instituto Amazónico de Investigaciones Científicas (SINCHI), Ministerio de Ambiente, Vivienda y Desarrollo Territorial. Bogotá, Colombia. 232 p. [ Links ]
SANTAS, L. A. 1983. Insects producing honeydew exploited by bees in Greece. Apidologie 14 (2): 92-103. [ Links ]
SORIA, A. C.; GONZÁLEZ, M.; DE LORENZO, C.; MARTÍNEZ-CASTRO, I.; SANZ, J. 2004. Characterization of artisanal honeys from Madrid (Central Spain) on the basis of their melissopalynological, physicochemical and volatile composition data. Food Chemistry 85: 121-13. [ Links ]
SORIA, A. C.; GONZÁLEZ, M.; LORENZO, C.; MARTÍNEZ-CASTRO, I.; SANZ, J. 2005. Estimation of the honeydew ratio in honey samples from their physicochemical data and from their volatile composition obtained by SPME and GC-MS. Journal of the Science of Food and Agriculture 85: 817-824. [ Links ]
VELÁSQUEZ, C. 1999. Atlas palinológico de la flora vascular paramuna de Colombia: Angiospermae. Universidad Nacional de Colombia. Medellín, Antioquia, Colombia. 173 p. [ Links ]
WILLIAMS, D. J.; GRANARA DE WILLINK, M. C. 1992. Mealybugs of Central and South America. CAB International, Wallingford, Inglaterra. 635 p. [ Links ]
