










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.39 no.2 Bogotá July/Dec. 2013
Sección Médica Nota científica
Efecto de inhibidores de anhidrasa carbónica en larvas de Anopheles albimanus (Díptera, Culicidae)
Effect of carbonic anhydrase inhibitors on Anopheles albimanus larvae (Diptera: Culicidae)
LEONARDO ROCHA O.1,2, CAROLINA TORRES G.1,3, MARÍA CORENA-McLEOD4, IYÁN D. VÉLEZ B.1,5 y SARA M. ROBLEDO R.1,6
1 PECET, Programa de Estudio y Control de Enfermedades Tropicales, Carrera 53 # 61-30, Torre 2 Laboratorio 632, Sede de Investigación Universitaria, Universidad de Antioqui a, Medellín, Antioquia.
2 M. Sc. Investigador PECET. lrocha94@yahoo.com.
3 M. Sc. Profesora, Universidad de Antioquia, Medellín. aniloract@gmail.com.
4 Ph. D. Directora World Solutions Against Infectious Diseases-WSAID, 4500 San Pablo Road.Jacksonville, FL 32224. pilarcorena@ aol.com.
5 Ph. D. Profesor, Universidad de Antioquia, Medellín. Director del PECET. idvelez@pecet-colombia.org.
6 Post Doc. Profesora, Universidad de Antioquia, Medellín. Investigador PECET. srobledo@saludpublica.udea.edu.co. Autor para correspondencia.
Recibido: 5-jul-2012 - Aceptado: 18-ago-2013
Resumen: La anhidrasa carbónica (AC) cataliza la conversión reversible del CO2 a bicarbonato y participa en mecanismos de alcalinización en el intestino de mosquitos. Se evalúo la toxicidad de cuatro inhibidores de anhidrasa carbónica (IAC): acetazolamida (AZM), metazolamida (MZM), etoxzolamida (ETX) y dorzolamida (DZA) en larvas de Anopheles albimanus por monitoreo de mortalidad 24, 48 y 72 horas post aplicación, a una concentración de 50 ug/ ml diluido previamente en dimetil sulfóxido. Todos los IAC redujeron la población de larvas en proporciones variables. La ETX mostró la mayor toxicidad que las AZM, MZM y DZA alcanzando una mortalidad superior al 80% después de 24 horas y del 98% después de 72 horas de su aplicación. Nuestros resultados indican que de los IAC, la etoxzolamida, en particular, constituye un candidato valioso como una nueva alternativa de control de formas inmaduras de An. albimanus, considerado el vector primario de malaria en Colombia.
Palabras clave: Larvicida. Control de mosquitos. Vectores. Paludismo.
Abstract: Carbonic anhydrase (CA) catalyzes the reversible conversion of CO2 to bicarbonate and participates in mechanisms of alkalinization in the intestine of mosquitoes. The toxicity of four CA inhibitors (CAI): acetazolamide (AZM), methazolamide (MZM), ethoxolamide (ETX) and dorzolamide (DZA) were evaluated in larvae of Anopheles albimanus by monitoring mortality 24, 48 and 72 hours post application, at a concentration of 50 ug/ml diluted in dimethyl sulfoxide previously. All IAC reduced the population of larvae in variable proportions. ETX showed the highest toxicity, achieving more than 80% mortality after 24 hours and 98% after 72 hours of application. The CAI, AZM, MZM and DZA showed less toxicity (<50% mortality). Our results indicate that the CAI, including ETX in particular, is a worthy candidate as an alternative for the control of An. albimanus, which is considered a primary vector of malaria in Colombia.
Key words: Larvicide. Mosquito Control. Vectors. Malaria.
Introducción
Anopheles (Nyssorhynchus) albimanus (Wiedemann, 1820) (Diptera: Culicidae) es uno de los vectores principales de Plasmodium de humanos en Centro y Sur América (Zimmerman 1992; Sinka et al. 2010; Montoya-Lerma et al. 2011). En Colombia, An. albimanus es considerado vector primario de malaria en áreas rurales y periurbanas de la Costa Pacífica y probablemente en la Costa Caribe (Gutiérrez et al. 2008). En el control de la malaria se presentan algunas limitaciones que incluyen el desconocimiento de actividades adecuadas para la protección y prevención de la transmisión; resistencia de algunas poblaciones de Plasmodium a los medicamentos de uso actual para el tratamiento de la malaria; errores en la aplicación de insecticidas o aun, resistencia de los vectores a los insecticidas; polución del medio ambiente y efectos adversos de los insecticidas en humanos y otros organismos (Georghiou y Taylor 1977, Olano et al. 1997; Hemingway y Ranson 2000; Carmona-Fonseca 2003, 2004; Nauen, 2007). Por esta razón, es fundamental la vigilancia de la resistencia a los insecticidas en uso (Fonseca-González et al. 2010) y la susceptibilidad de los estadios inmaduros a los insecticidas.
El control de inmaduros es una estrategia más factible que el de los adultos, dado que limita la capacidad de dispersión de los mosquitos (Blanco et al. 2002). El control de formas inmaduras de anofelinos, requiere el conocimiento de las condiciones de calidad de agua que puede ocupar una especie (Sinka et al. 2010). Además, se debe conocer aspectos biológicos básicos que permitan el descubrimiento de blancos necesarios para el desarrollo de nuevos y mejores larvicidas. Un aspecto fundamental en larvas de mosquitos es la alcalinización, la cual es indispensable para la supervivencia (Co-rena et al. 2004). Las enzimas relacionadas con este proceso pertenecen a la familia de las anhidrasas carbónicas (AC), las cuales catalizan la conversión reversible del CO2 a bicarbonato (Corena et al. 2002; Smith et al. 2007). La AC es inac-tivada por dos clases de compuestos: aniones complejos de metales y sulfonamidas no sustituidas, las cuales se unen al ión de zinc de las AC. Algunos de estos compuestos, como la acetazolamida (AZM) y metazolamida (MZM), en larvas de mosquitos como Aedes aegypti, Ae. albopictus, Culex quin-quefasciatus, Cx. nigripalpus, Ochlerotatus taeniorhynchus, An. quadrimaculatus, ocasionan un efecto letal variable de acuerdo con la especie (Corena et al. 2004). Sin embargo, hasta el momento se desconoce el efecto de estos compuestos en larvas de An. albimanus. El objetivo de este trabajo fue evaluar el efecto letal de cuatro IAC en larvas de esta especie, bajo condiciones de laboratorio.
Materiales y métodos
Mosquitos. Larvas de An. albimanus, cepa Cartagena (Carrillo et al. 1981), donada por el Instituto Nacional de Salud de Colombia, fueron criadas y mantenidas en el insectario del PECET, Programa de Estudio y Control de Enfermedades Tropicales. En la cría se usaron bandejas plásticas con agua reposada y una combinación de levadura y peletizado para peces (1:1) como alimento. Los experimentos se desarrollaron a 27 ± 2 °C, ciclo de luz/oscuridad 12:12 y 75 ± 10% humedad relativa.
Compuestos. Los IAC: acetazolamida (AZM), matazolami-da (MZM), etoxzolamida (ETX) (Sigma-Aldrich, St. Louis, MO, EEUU) y dorzolamida (DZA) (Merck Sharp & Doh-me, Chibret, Francia) se obtuvieron comercialmente. Estos son medicamentos de uso humano empleados principalmente como diuréticos y en el tratamiento de enfermedades como el glaucoma. Todos los IAC se prepararon en dimetil sulfóxi-do (DMSÜ) al 0,5% (Sigma-Aldrich) (Corena et al. 2005). Como control positivo se incluyó un grupo de larvas tratadas con temefos (Abate® 1SG granulado) al 1%.
Bioensayos. Se realizaron de acuerdo con las guías establecidas para evaluación de larvicidas en laboratorio y campo ÜMS (2005). Veinte larvas de III estadio tardío o IV temprano se transfirieron a recipientes que contenían 100 ml de agua destilada. El experimento se repitió tres veces en días distintos empleando una concentración de 50 ng/ml para cada IAC. Esta concentración fue seleccionada de acuerdo con trabajos de Corena et al. (2004). Los controles utilizados consistieron en un grupo tratado con temefos a 50 ng /ml (control positivo), un grupo con DMSÜ al 0,5% (diluyente de los IAC) y un grupo sin tratamiento (control negativo). La lectura de la mortalidad se realizó a las 24, 48 y 72 horas post aplicación.
Criterio de toxicidad y análisis estadístico. La toxicidad de los IAC sobre las larvas se calculó de acuerdo con el porcentaje de mortalidad observada, y sus respectivos intervalos de confianza al 95%, en cada tiempo de evaluación. Se realizó corrección por fórmula de Abbot, cuando se encontró mortalidad en los controles entre el 5 y 20%. Se hizo una prueba de Shapiro Wilk para determinar normalidad y un test de Kruskal-Wallis para establecer si había diferencias significativas en los promedios de mortalidad entre los tratamientos. Luego, se aplicaron pruebas no paramétricas de comparaciones múltiple mediante el test de Tukey. Los niveles de significancia correspondieron con un valor P < 0.05. Se utilizó el programa Graph Pad Prism 5.
Resultados
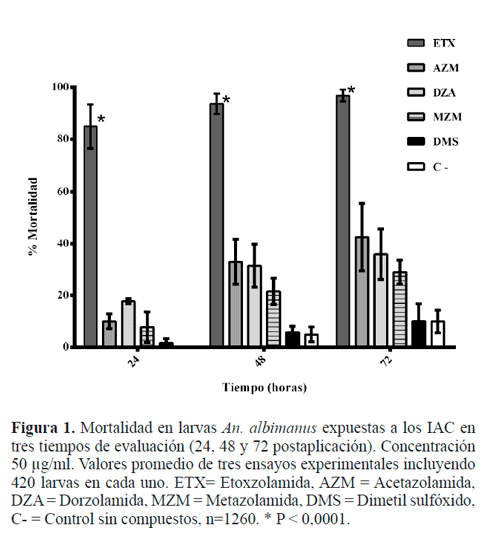
El tratamiento de larvas An. albimanus con los IAC produjo una toxicidad variable siendo ETX el que mostró mayor toxicidad y mortalidad. Ésta fue mayor al 80% después de 24 horas de aplicación y después de 48 y 72 horas alcanzó su valor máximo de 98% (Fig. 1). Este porcentaje fue significantemente diferente (P < 0,0001), con respecto a los otros IAC y el control sin tratamiento. En contraste, las mortalidades con AZM, MZM y DZA fueron menores al 60%. El control con DMSÜ no arrojó diferencias estadísticamente significativas con respecto al control. El control positivo registró una mortalidad de 100% en larvas 24 horas después de la aplicación (datos no mostrados).
Discusión
El efecto de mortalidad de algunos IAC sobre estadios inmaduros de insectos ha sido estudiado en pocas especies como Ae. aegypti, An. quadrimaculatus, Cx. quinquefasciatus, Cx. nigripalpus, Oc. taeniorhynchus y Ae. albopictus (Corena et al. 2004). Nuestros hallazgos indican que la MZM produjo el menor efecto tóxico (< 40% mortalidad), contrario a lo encontrado por Corena et al. (2004), donde MZM causó una mortalidad del 80% en larvas del mismo género a una concentración similar a la utilizada en el presente estudio (10-4 M). La mortalidad en larvas se atribuye al desequilibrio en el pH del intestino medio y la inhibición de los procesos de transporte de iones (Corena et al. 2004). Las diferencias en el porcentaje de mortalidad para cada especie, pueden obedecer a un mecanismo de respuesta específico o la presencia de una familia particular de AC (Corena et al. 2004). Una de las formas de comprender mejor el efecto de los IAC, es el análisis de los productos de excreción liberados en el agua. Si bien este aspecto no se incluyó en este trabajo, sería interesante considerarlo para estudios futuros dado que se pueden esperar variaciones de acuerdo no solo con la especie sino con el estadio evaluado (Corena et al. 2004). El efecto tóxico de ETX (mortalidad cercana al 100%) después de 24 horas, es un resultado importante, comparable al efecto larvicida logrado por compuestos como el temefos®, utilizado en el control de Ae. aegypti y An. nueztovari en condiciones laboratorio y campo (González y Palacio Mosquera, 2009; Maestre et al. 2009). La inclusión del temefos sirvió exclusivamente como un control positivo y no con el propósito de establecer comparaciones con los IAC. ETX posee propiedades inhibitorias sobre el dominio soluble de la alfa-ACl que se encuentra en Ae. aegypti, mostrando afinidad a nivel nanomolar (Fisher et al. 2006). Es muy probable que un dominio homólogo exista en An. albimanus y, por tanto, el efecto de ETX en larvas se observe en mayor medida. Además, de acuerdo con las propiedades físicas de la ETX, se indica que es un compuesto prácticamente insoluble en el agua, hecho que favorece su permanencia en la superficie del agua, lugar donde se alimentan y permanecen la mayor parte del tiempo las larvas del género Anopheles logrando de esta forma una mayor mortalidad.
Por otro lado, la ETX en los insectos produce un desequilibrio en el pH del intestino y además causa la inhibición de procesos de transporte de iones (Corena et al. 2004). Las propiedades letales de la ETX sobre la fisiología de las larvas propende su uso en el control, no sin antes considerar su efecto en organismos no blanco.
Según nuestros resultados la ETX es un compuesto importante que puede ser utilizado como una estrategia complementaria con las medidas de control. El alto porcentaje de mortalidad observado en larvas expuestas a ETX, señala que podría considerarse como un compuesto promisorio en la búsqueda de nuevos blancos larvicidas para An. albimanus.
Agradecimientos
A Evencio Mosquera por su valiosa ayuda en la cría del material biológico empleado en este estudio, a María Fernanda Flórez, María Alejandra Duque, Catalina Marín y Esteban Marín por su colaboración en la ejecución de los bioensayos. Este trabajo fue financiado por Colciencias y la Universidad de Antioquia.
Literatura citada
BLANCO CASTRO, S. D.; COLOMBI, E.; FLORES, L. N. CANALES, D. 2002. Aplicación del biolarvicida Bacillus sphaeri-cus-2362 (Griselesf) para el control de la malaria en un área de salud de la República de Honduras. Revista Cubana de Medicina Tropical 54: 134-141. [ Links ]
CARMONA-FONSECA, J. 2003. La malaria en Colombia, Antioquia y las zonas de Urabá y Bajo Cauca. Parte 1. Iatreia 16: 299-317. [ Links ]
CARMONA-FONSECA, J. 2004. La malaria en Colombia, Antioquia y las zonas de Urabá y Bajo Cauca. Parte 2. Iatreia 17: 34-53. [ Links ]
CARRILLO, M. P.; SUÁREZ, M. F.; MORALES, A.; ESPINAL, C. A. 1981. Colonización y mantenimiento de una cepa colombiana de Anopheles albimanus, Wiedemann, 1820 (Diptera: Culicidae). Biomédica 1: 64-66. [ Links ]
CORENA, M. P.; SERON, T. J; LEHMAN, H. K.; OCHRIETOR, J. D.; KOHN, A.; TU, C.; LINSER, P. J. 2002. Carbonic anhydrase in the midgut of larval Aedes aegypti: cloning, localization and inhibition. The Journal of Experimental Biology 205: 591-602. [ Links ]
CORENA, M. P.; FIEDLER, M. M.; VANEKERIS, L.; TU, C.; SILVERMAN, D. N.; LINSER, P. J. 2004. Alkalization of larval mosquito midgut and the role of carbonic anhydrase in different species of mosquitoes. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 137 (3): 207-225. [ Links ]
CORENA, M.; VANEKERIS, L.; SALAZAR, M. I.; BOWERS, D.; FIEDLER, M. M.; SILVERMAN, D.; TU, C.; LINSER, P. J. 2005. Carbonic anhydrase in the adult mosquito midgut. The Journal of Experimental Biology 208: 3263-3273. [ Links ]
FISHER, S. Z.; TARIKU, I.; CASE, N. M.; TU, C.; SERON, T.; SILVERMAN, D. N.; LINSER, P. J.; MCKENNA, R. 2006.Expression, purification, kinetic, and structural characterization of an alpha-class carbonic anhydrase from Aedes aegypti (AaCAl). Biochimica et Biophysica Acta 1764 (8): 1413-1419. [ Links ]
FONSECA, I.; CÁRDENAS, R.; GÓMEZ, W.; SANTACOLOMA, L.; BROCHERO, H.; OCAMPÓ, C. B.; SALAZAR, M.; MCALLISTER, J.; BROGDON, W.; QUIÑONEZ, M. 2010. Dosis diagnósticas para vigilar la resistencia a insecticidas de los vectores de malaria en Colombia. Revista Colombiana de Entomología 36: 54-61. [ Links ]
GEORGHIOU, G. P.; TAYLOR, C. E. 1977. Genetic and biological influences in the evolution of insecticide resistance. Journal of Economic Entomology 70 (3): 319-23. [ Links ]
GONZÁLEZ, R.; PALACIO MOSQUERA, J. D. 2009. Evaluación de la susceptibilidad y persistencia de los insecticidas Temephos (Abate) y Temephos (Instarphos), con exposición de larvas de Aedes aegypti y Anopheles nuñeztovari (Diptera: Culicidae). Vectors and Pest Magazine 1: 23-26. [ Links ]
GUTIÉRREZ, L. A.; NARANJO, N.; JARAMILLO, L. M.; MUS-KUS, C.; LUCKHART, S.; CONN J. E.; CORREA, M. M. 2008. Natural infectivity of Anopheles species from the Pacific and Atlantic regions of Colombia. Acta Trópica 107 (2): 99-105. [ Links ]
HEMINGWAY, J.; RANSON, H. 2000. Insecticide resistance in insect vectors of human disease. Annual Review of Entomology 45: 371-391. [ Links ]
MAESTRE, R.; REY, G.; DE LAS SALAS, J.; VERGARA, S. C.; SANTACOLOMA, L.; GOENAGA, S.; CARRASQUILLA, M. C. 2009. Susceptibilidad de Aedes aegypti (Diptera: Culicidae) a temefos en Atlántico-Colombia. Revista Colombiana de Entomología 35 (2): 202-205. [ Links ]
MONTOYA-LERMA, J.; SOLARTE, Y. A.; GIRALDO-CALDE-RÓN, G. I.; QUIÑONES, M. L.; RUIZ-LÓPEZ, F.; WILKER-SON, R. C.; GONZÁLEZ, R. 2011. Malaria vector species in Colombia - A Review. Memórias do Instituto Oswaldo Cruz 106 (suppl. I): 223-238. [ Links ]
NAUEN, R. 2007, Insecticide resistance in disease vectors of public health importance. Pest Managment Science 63 (7): 628-633. [ Links ]
OLANO, V.; CARRASQUILLA, G.; MÉNDEZ, F. 1997. Transmisión de la malaria urbana en Buenaventura, Colombia: aspectos entomológicos. Revista Panamericana de Salud Pública 1 (4): 287-294. [ Links ]
ORGANIZACION MUNDIAL DE LA SALUD (WORLD HEALTH ORGANIZATION) OMS. 2005. Guidelines for laboratory and field testing of mosquito larvicides. WHO/CDS/ WHOPES/GCDPP/2005.3. [ Links ]
SINKA, M. E.; RUBIO-PALIS, Y.; MANGUIN, S.; PATIL, A. P.; TEMPERLEY, W. H.; GETHING, P. W.; VAN BOECKEL, T.; KABARIA, C. W.; HARBACH, R. E.; HAY, S. I. 2010.The dominant Anopheles vectors of human malaria in the Americas: occurrence data, distribution maps and bionomic précis. Parasites & Vectors 3: 1-34. [ Links ]
SMITH, K. E.; VANEKERIS, L. A.; LINSER, P. J. 2007.Cloning and characterization of AgCA9, a novel W-carbonic anhydrase from Anopheles gambiae Giles sensustricto (Diptera: Culicidae) larvae. Journal of Experimental Biology 210: 3919-3930. [ Links ]
ZIMMERMAN, R. H. 1992. Ecology of malaria vectors in the Americas and future directions. Memórias do Instituto Oswaldo Cruz 87 (suppl. III): 371-383. [ Links ]
Citación sugerida:
ROCHA O., LEONARDO; CAROLINA TORRES G.; MARÍA CORENA-McLEOD; IVÁN D. VÉLEZ B. y SARA M. ROBLEDO R. 2013. Efecto de inhibidores de anhidrasa carbónica en larvas de Anopheles albimanus (Diptera, Culicidae). Revista Colombiana de Entomología 39 (2): 226-228.
