










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.1 Bogotá Jan./June 2014
Development and reproduction of Neoleucinodes elegantalis (Lepidoptera: Crambidae) on tomato (Solanum licopercum) cultivars
Desarrollo y reproducción de Neoleucinodes elegantalis (Lepidoptera: Crambidae) en cultivares de tomate (Solanum licopercum)
CARLA PEDROSO DE MORAES1 AND LUÍS AMILTON FOERSTER2
1 Doctoral candidate. Department of Phytotechnology and Plant Health, Agricultural Sciences, Federal University of Parana, CP 19.061, CEP: 81.531-990, Brazil. kalengagri@yahoo.com.br. Corresponding author.2 Ph. D. Departament of Zoology, Biological Sciences, Federal University of Parana, Brazil.
Received: 25-Apr-2013 • Accepted: 16-Feb-2014
Abstract: The small tomato borer Neoleucinodes elegantalis (Lepidoptera: Crambidae) is one of the most representative pests of Solanaceae in Brazil and other South American countries. Due to the restricted knowledge about the influence of tomato cultivars on the biology of the species, the development and reproduction of N. elegantalis was evaluated on three tomato cultivars. In the hybrid Giuliana the development from oviposition to adult emergence was longest (56.2 days) in comparison to the cultivar Santa Clara (50.2 days) and the hybrid Paronset (50.7 days). Pupal weight was significantly lower in Giuliana. The mean fecundity of females fed on Giuliana during the larval stage was 88.6 eggs per female, 82.3 in Paronset and 37.0 on Santa Clara, but egg viability was higher for females fed during the larval stage on Santa Clara (79.0%). The hybrid tomato Giuliana had the highest impact on the development and reproduction of N. elegantalis. These findings can be useful as a plant resistance factor in the field to reduce the number of generations of the pest and to favour the action of natural enemies by decreasing the fitness of the pest.
Key words: Small tomato borer. Biology. Solanaceae.
Resumen: El pequeño barrenador del tomate Neoleucinodes elegantalis (Lepidoptera: Crambidae) es una de las plagas más destructivas de las solanáceas en Brasil y otros países suramericanos. Debido al conocimiento limitado acerca de la influencia de los cultivares de tomate sobre la biología, se evaluó el desarrollo y la reproducción de N. elegantalis en tres cultivares de tomate. En el híbrido Giuliana el desarrollo de la oviposición hasta la emergencia del adulto fue más larga (56,2 días) en comparación con el cultivar Santa Clara (50,2 días) y el híbrido Paronset (50,7 días). El peso pupal fue significativamente menor en Giuliana. La fecundidad media de hembras alimentadas con Giuliana, Paronset y Santa Clara durante la etapa larval fue de 88,6; 82,3 y 37,0 huevos por hembra, respectivamente. Sin embargo, la viabilidad de los huevos fue mayor para las hembras alimentadas durante la etapa larval en Santa Clara (79,0%). El híbrido de tomate Giuliana tuvo el mayor impacto en el desarrollo y la reproducción de N. elegantalis. Estos hallazgos pueden ser útiles como un factor de resistencia de la planta en el campo para reducir el número de generaciones de la plaga y para favorecer la acción de los enemigos naturales por la disminución del "fitness" de la plaga.
Palabras clave: Perforador del fruto del tomate. Biología. Solanaceae.
Introduction
Tomato crops (Solanum licopercum L.) have a great economic and social importance worldwide (Jordão and Nakano 2002). The small tomato borer Neoleucinodes elegantalis (Guenée) (Lepidoptera: Crambidae) is a pest of economic importance due to the damage caused in the fruits and thus reducing the commercial value of the crop (Leiderman and Sauer 1953; Picanço et al. 2007). The distribution of N. elegantalis is strictly Neotropical including both Central and South America (Capps 1948).
The damage caused by N. elegantalis is direct, a single larva is sufficient to depreciate and render the fruit unsuitable for consumption (Toledo 1948). Yield losses are estimated at as much as 50% (Gallo et al. 2002), 79% (Miranda et al. 2005) and 90% (Carneiro et al. 1998). According to Blackmer et al. (2001) under field conditions the females lay their eggs preferably beneath the sepals of small (23.0 mm diameter) green fruits and soon after hatching the larva penetrates the fruit and causes the irreversible damage.
The knowledge on the biology of the small tomato borer is scarce; Marcano (1991a) evaluated the development time and reproduction of N. elegantalis on tomato fruits of the cultivar Rio Grande. Other studies include the ecology and behavior of N. elegantalis (Salas et al. 1991) and the role of pheromones (Badji et al. 2003). Blackmer et al. (2001) and Barbosa et al. (2010) evaluated the preference for oviposition of N. elegantalis on tomato and Jaffe et al. (2007) investigated the role of chemical stimulus on the sex attraction of N. elegantalis. Benvenga et al. (2010) conducted field samplings of the small tomato borer for decision making on the control in staked tomato plants.
Apart from tomato, N. elegantalis is able to develop and reproduce in other cultivated Solanaceae, green peppers, eggplant and scarlet eggplant (Toledo 1948, Zucchi et al. 1993). On scarlet eggplant Picanço et al. (1997) found all stages of N. elegantalis and described the tunneling caused by the larvae in the fruits. Uncultivated solanaceous plants also host the pest (Toledo 1948, Zucchi et al. 1993).
Due to the limited efficiency of chemical control measures against the damage of boring insects such as N. elegantalis, research on cultivar performance is a sound alternative to reduce the larval damage on tomato yield. This work was conducted to evaluate the effect of three tomato cultivars on the growth, reproduction and survival of N. elegantalis.
Materials and methods
A colony of N. elegantalis was established in laboratory from larvae collected on tomato hybrid Alambra crops in Southern Paraná state (25°19'S 49°18'W). Larvae were reared on tomato fruits of the group Salad. Field collected larvae were introduced into the colony every week in order to maintain the genetic diversity of the population. One cultivar (Santa Clara) and two hybrids (Giuliana and Paronset) were employed to compare the development cycle, reproduction and longevity of N. elegantalis. These materials were chosen for being commonly employed by local tomato growers and their high commercial value. Tomato plants were grown in a greenhouse in 5 L pots. Green tomato fruits, 23 to 40 mm in diameter from each cultivar were used in the experiment. For surface disinfection, the fruits were immersed in water + 1% sodium hypochlorite for five minutes, rinsed under running water and dried with paper towels. Three neonate larvae were incubated in each fruit totaling 99 larvae per tomato cultivar. The experiment was conducted in a climatic chamber at a constant temperature of 20 ± 1 °C, relative humidity of 60 ± 10% and photoperiod of 12:12 h (light: dark). At the end of the larval period, the insects left the tomatoes to pupate outside the fruits. Pupae were kept in 7x4 cm polyethylene vials and after 72 hours were weighed and checked for the presence of deformities. Sex determination was made after adult emergence according to Muñoz et al. (1991) and Carneiro et al. (1998).
The biological parameters evaluated were development time, larval and pupal viability, pupal weight and sex-ratio (females / males + females). To evaluate the influence of the cultivar consumed in the larval stage on adult performance, twenty pairs were formed from each treatment. Each pair was kept in polyethylene tubes 20 cm high x 10 cm diameter closed at the bottom and on the top with a polyethylene cover. Tomato leaves of each cultivar were placed inside the cages as stimulants for oviposition. The adults were fed with 10% honey diluted in water, provided in a cotton ball soaked in the solution. Food renewal and egg collection were carried out daily. The eggs were separated according to the female and kept in moistured paper towel to assess the incubation period and viability.
The effect of the cultivars consumed by larvae on the adult stage of N. elegantalis was evaluated through longevity, length of pre-oviposition and oviposition periods, number of eggs laid and their viability.
The experimental design was completely randomized, the condition of normality of the data was submitted to Shapiro- Wilks test. Results were subjected to analysis of variance and the means were compared by Tukey's test or nonparametric Kruskal-Wallis test (P < 0.05). Data were analyzed with Statistica 7.0 software (Statsoft 2004).
Results and discussion
The incubation period of N. elegantalis at 20 ± 1 °C lasted from 7.3 to 7.4 days, did not differ significantly among the cultivars (Table 1). There are no other references on the development of N. elegantalis other than the work of Marcano (1991a) which describes an incubation period of 7.1 days at 20 °C.

Larval survival was not significantly different among the treatments (P = 0.162) (Table 1). The larval stage was significantly longer when the larvae were reared on Giuliana fruits (29.5 days) in comparison to Paronset (26.1 days) and Santa Clara (25.8 days) (Table 1). There are no data in the literature comparing the influence of different tomato cultivars on the biological traits of N. elegantalis. Marcano (1991a, 1991b) estimated the development of N. elegantalis on tomato and eggplant; when feeding on tomato, the larval period lasted 22.7 days compared to 31.4 days for larvae reared on eggplant fruits. According to Muñoz (1991) newly hatched larvae move by means of a silk thread until a point chosen for penetration through the epicarp and then feed on the mesocarp and later reach the endocarp where the rate of feeding is higher. In the endocarp the larvae tunnel galleries and feed on the placenta and seeds provoking most of the damage seen on the fruits.
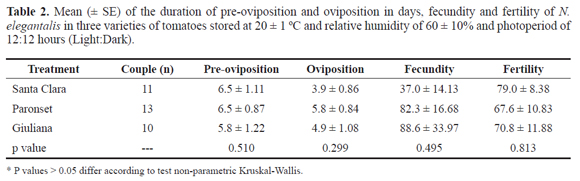
Between the end of the larval stage and the beginning of the pupal phase N. elegantalis remained over five days in the pre-pupal stage (Table 1). No deformed pupae were recorded among the three cultivars. Likewise the larval development, adults took longer to emerge from the pupae proceeding from Giuliana in comparison to the other cultivars (Table 2). These values are similar to the ones reported by Marcano (1991a, 1991b) for eggplant and tomato fruits for the pupal stage of N. elegantalis (13.9 days). Paredes et al. (2010) also recorded a pupal period of 13 days for the species.
The sex-ratio of N. elegantalis was 0.4 in all cultivars. A predominance of males was also recorded by Salas (1992), Carneiro et al. (1998) and Jaffe et al. (2007). Fernández and Salas (1985) found in 688 sexed pupae 370 females and 318 males.
During the reproductive period, no statistical differences were observed among the treatments in relation to the preoviposition (P = 0.510) and oviposition (P = 0.299) periods, oviposition rate (P = 0.495) and egg viability (P = 0.813). Mating success of N. elegantalis is low in laboratory; only 10 out of 20 pairs evaluated laid eggs on Giuliana, 11 on Santa Clara and 13 on Paronset (Table 2). According to Jaffe et al. (2007) N. elegantalis copulates monogamously, more vigorous males have a faster answer to the sexual pheromone released by females, thus functioning as a process of natural selection. This behavior explains the low number of females ovipositing, since the couples were randomly formed and no option were given for the choice of partners by the females.
The pre-oviposition period (Table 2) was more than twice longer than the values reported by Marcano (1991a) and Muñoz et al. (1991). These authors used tomato leaves as stimulants for oviposition instead of fruits as we did and the longer pre-oviposition period may be due to the less stimulant effect of tomato fruits in comparison to leaves. The oviposition period in N. elegantalis lasted less than a week for females emerged from the three cultivars (Table 2), similar to the value found by Carneiro et al. (1998), but longer than the two days of oviposition described by Marcano (1991a).
The mean number of eggs laid by each female was more than twice as high in Giuliana and Paronset than in Santa Clara (Table 2). However due to the wide amplitude in the number of eggs laid per female, no statistical difference was detected among the treatments, indicating that the cultivar had no effect on the fertility of N. elegantalis. Marcano (1991a) obtained a mean of 52.3 eggs per female, also with a large variation in the number of eggs per female among the replicates (19 to 77 eggs/female).
No differences were recorded on egg viability among treatments, which ranged from 67.6% in females reared in the larval stage on Paronset to 79.0% in Santa Clara (Table 2). Similar values were described by Fernández and Salas (1985) in tomatoes, while Muñoz et al. (1991) recorded 98.0% of egg viability when the larvae were reared on nightshade fruits, a perennial solanaceaus species, Solanum quitoense grown in Northwestern South America.
The longevity of males and females was longer than 15 days, and no differences were found both among cultivars and between the sexes (Table 3). These values are more than twice the longevity recorded by Marcano (1991a, 1991b) for N. elegantalis kept at the same temperature and reared during the larval stage on eggplant and tomato fruit.

Development from egg hatching to adult emergence resulted significantly longer when larvae were reared on the cultivar Giuliana (56.2 days) in comparison to Paronset and Santa Clara (50.2 and 50.7 days, respectively). This difference may be due to the effect of antibiosis caused by cultivar, because according to Smith (2005) antibiotic effects may be expressed in a moderate way, increasing the duration of development and reducing weight or decreasing the adult life span. These results differ from those found by Marcano (1991a), which reported the development cycle in 43.7 days using cultivar Rio Grande. In the field the cultivar Santa Clara is considered as susceptible to the major insect pests and diseases of tomato (Bettiol et al. 2004) and thus is employed in laboratory tests as a pattern for susceptibility. According to the companies responsible for the hybrids (Sakata Seeds for Giuliana and Syngenta Seeds for Paronset), these hybrids possess genetic resistance to numerous diseases, but little is known about their tolerance to insect attack.
Previous results showed significant differences in the performance of N. elegantalis developing on different tomato cultivars (Lara et al. 1980, Moreira et al. 1985, Lyra Netto and Lima 1998). According to Viáfara et al. (1999) increased susceptibility of N. elegantalis to industrial tomato cultivars, is assigned to the higher content of soluble solids, which promote the feeding habits of the larvae.
Conclusion
The increased development time and the lower pupal weight of the small tomato borer reared on the hybrid Giuliana may reduce larval fitness, and thus varietal control can function as a strategy in programs of integrated pest management of N. elegantalis in tomato.
Acknowledgements
We are grateful to the tomato growers in the municipality of Almirante Tamandaré – Paraná, for allowing the collection of larvae in their production areas and to Dr. Gedir de Oliveira Santos of the Department of Botany – UFPR, for the use of a greenhouse for growing tomatoes. To the Support Program for the Restructuring and Expansion of Federal Universities (Reuni) for the scholarship granted to the first author.
Literature cited
BADJI, C. A.; EIRAS, A. E.; CABRERA, A.; JAFFE, K. 2003. Avaliação do feromônio sexual de Neoleucinodes elegantalis Guenée (Lepidoptera: Crambidae). Neotropical Entomology 32 (2): 221-229. [ Links ]
BARBOSA, F. S.; MENEZES, E. L. A; ARRUDA, L. N.; BRITO, D. O.; CARMO, M. G. F.; PEREIRA, M. B. 2010. Preferência de oviposição de broca-pequena-do-fruto em oito variedades de tomate rasteiro. Horticultura Brasileira 28: S634-S640. [ Links ]
BENVENGA, S. R.; DE BORTOLI, A. S.; GRAVENA, S.; BARBOSA, J. C. 2010. Monitoramento da broca-pequena-do-fruto para tomada de decisão de controle em tomateiro estaqueado. Horticultura Brasileira 28: 435-440. [ Links ]
BETTIOL, W.; GHINI, R.; GALVAO, J. A. H.; SILOTO, R. C. 2004. Organic and conventional tomato cropping systems. Scientia Agricola 61: 253-259. [ Links ]
BLACKMER, J. L.; EIRAS, A. E.; SOUZA, C. L. M. 2001. Oviposition preference of Neoleucinodes elegantalis (Guenée) (Lepidoptera: Crambidae) and rates of parasitism by Trichogramma pretiosum (Riley) (Hymenoptera: Trichogrammatidae) on Lycopersicon esculentum in São José de Ubá, Brazil. Neotropical Entomology 30: 89-95. [ Links ]
CAPPS, H. W. 1948. Status of the pyraustid moths of the genus Leucinodes in the world, with descriptions of news genus and species. Proceedings of the United States National Museum 98: 69-85. [ Links ]
CARNEIRO, J. S.; HAJI, F. N. P.; SANTOS, F. A. M. 1998. Bioecologia e controle da broca-pequena-do-tomateiro Neoleucinodes elegantalis. Teresina: Embrapa Meio-Norte / Circular Técnica, 26: 14 p. [ Links ]
FERNÁNDEZ, S.; SALAS, J. 1985. Estúdios sobre la biología del perfurador del tomate Neoleucinodes elegantalis Guenee (Lepidoptera: Pyraustidae). Agronomía Tropical 35: 77-82. [ Links ]
GALLO, D.; NAKANO, O.; NETO, S. S.; CARVALHO, R. P. L.; BATISTA, G. C.; FILHO, E. B.; PARRA, J. R. P.; ZUCCHI, R. A.; ALVES, S. B.; VENDRAMIM, J. D.; MARCHINI, L. C.; LOPES, J. R. S.; OMOTO, C. 2002. Entomologia agrícola. Piracicaba: FEALQ, 920 p. [ Links ]
JAFFE, K.; MIRÁS, B.; CABRERA, A. 2007. Mate selection in the moth Neoleucinodes elegantalis: evidence for a supernormal chemical stimulus in sexual attraction. Animal Behavior 73: 727-734. [ Links ]
JORDÃO, A. L.; NAKANO, O. 2002. Ensacamento de frutos do tomateiro visando ao controle de pragas e à redução de defensivos. Scientia Agricola 59: 281-289. [ Links ]
LARA, F. M.; BARBIERI, J.; CHURATA-MASCA, M. G. C. 1980. Resistance of processing tomatoes (Lycopersicon esculentum) to Neoleucinodes elegantalis (Lepidoptera: Pyraustidae). Anais da Sociedade Entomológica do Brasil 9 (1): 53-66. [ Links ]
LEIDERMAN, L.; SAUER, H. F. G. 1953. A broca-pequena-dofruto do tomateiro. O Biológico 19: 182-186. [ Links ]
LYRA NETTO, A. M. C.; LIMA, A. A. F. 1998. Infestação de cultivares de tomateiro por Neoleucinodes elegantalis (Lepidoptera: Pyralidae). Pesquisa Agropecuária Brasileira 33: 2. [ Links ]
MARCANO, R. V. 1991a. Estudio de la biología y algunos aspectos del comportamiento del perforador del fruto del tomate Neoleucinodes elegantalis (Lepidoptera: Pyralidae) en tomate. Agronomía Tropical 41: 257-263. [ Links ]
MARCANO, R. V. 1991b. Ciclo biológico del perforador del fruto del tomate Neoleucinodes elegantalis (Guenée) (Lepidoptera: Pyralidae), usando berenjena (Solanum melongena) como alimento. Boletín Entomología Venezolana 6: 135-141. [ Links ]
MIRANDA, M. M. M.; PICANÇO, M. C.; ZANUNCIO, J. C.; BACCI, L.; SILVA, E. M. 2005. Impact of integrated pest management on the population of leafminers, fruit borers, and natural enemies in tomato. Ciência Rural 35: 204-208. [ Links ]
MOREIRA, J. O. T.; LARA, M. E.; CHURATA MASCA, M. G. C. 1985. Resistência de cultivares de tomateiro (Lycopersicon esculentum Mill) a broca-pequena-do-fruto. Ciência e Cultura 37 (4): 618-623. [ Links ]
MUÑOZ, L. E.; SERRANO, A. P.; PULIDO, J. I.; DE LA CRUZ, J. L. 1991. Ciclo de vida y enemigos naturales de Neoleucinodes elegantalis (Guenée) (Lepidoptera: Pyralidae) pasador del fruto del lulo Solanum quitoense Lam. en el Valle del Cauca. Acta Agronómica 41: 99-104. [ Links ]
PAREDES, J.; PERALTA, E. L.; GÓMEZ, P. 2010. Gusano perforador de los frutos de naranjilla (Solanum quitoense Lam): identificación y biología. Revista Tecnológica ESPOL – RTE 23: 27-32. [ Links ]
PICANÇO, M. C.; CASALI, V. W. D.; LEITE, G. L. D.; OLIVEIRA, I. R. 1997. Lepidópteros associados ao jiloeiro. Horticultura Brasileira 15: 112-114. [ Links ]
PICANÇO, M. C.; BACCI, L.; SILVA, E. M.; MORAIS, E. G. F.; SILVA, G. A.; SILVA, N. R. 2007. Manejo integrado das pragas do tomateiro no Brasil. pp.199-232. In: Silva, D. J. H; Vale, F. X. R. (Eds.). Tomate: tecnologia de produção. Viçosa: UFV. [ Links ]
SALAS, J. 1992. Integrated pest-insects management program for tomato crops (Lycopersicon esculentum Mill.) Lara State, Venezuela. Acta Horticulturae 301: 199-204. [ Links ]
SALAS, J.; ÁLVAREZ, C.; PARRA, A. 1991. Contribución al conocimiento de la ecología del perforador del fruto del tomate Neoleucinodes elegantalis (Guenée) (Lepidoptera: Pyraustidae). Agronomía Tropical 41: 275-283. [ Links ]
SMITH, C. M. 2005. Plant resistance to arthropods: molecular and conventional approaches. Berlin: Springer. 423 p. [ Links ]
STATSOFT INC. 2004. Statistica for Windows. Tulsa, Statsoft Inc. [ Links ]
TOLEDO, A. A. 1948. Contribuição para o estudo de Leucinodes elegantalis, praga do tomate. O Biológico 14: 103-108. [ Links ]
VIÁFARA, H. F.; GARCÍA, F.; DÍAZ, A. E. 1999. Parasitismo natural de Neoleucinodes elegantalis (Guenée) (Lepidóptera: Pyralidae) en algunas zonas productoras de Solanáceas del Cauca y Valle del Cauca, Colombia. Revista Colombiana de Entomología 25: 151-159. [ Links ]
ZUCCHI, R. A.; SILVEIRA NETO, S.; NAKANO, O. 1993. Guia de identificação de pragas agrícolas. Piracicaba: FEALQ, 139 p. [ Links ]
Suggested citation:
MORAES, C. P. de; FOERSTER, L. A. 2014. Development and reproduction of Neoleucinodes elegantalis (Lepidoptera: Crambidae) in cultivars of tomato (Solanum licopercum L.). Revista Colombiana de Entomología 40 (1): 40-43. Enerojunio 2014. ISSN 0120-0488.
