










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.1 Bogotá Jan./June 2014
Effect of larval nutrition on the development and mortality of Galleria mellonella (Lepidoptera: Pyralidae)
Efecto de la nutrición larval en el desarrollo y mortalidad de Galleria mellonella (Lepidoptera: Pyralidae)
ABEDELSALAM ANWAR MOHAMED1, MOHAMED JAVED ANSARI2, AHMAD AL-GHAMDI3, MOHAMED OMAR MOHAMED4 AND MANPREET KAUR5
1 Abdullah Ahmad Bagshan for Bee Research, Department of Plant Protection, College of Food and Agriculture Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Kingdom of Saudi Arabia.
1,2 Ph.D. abdhassan@ksu.edu.sa.
1,3 Ph.D. mjavedansari@gmail.com corresponding author.
1,4 Ph.D. aalkhazim@gmail.com.
1,5 Ph.D. dr_momar@hotmail.com.
2 Forest Botany Division, Forest Research Institute, P.O. Box New Forest, Dehradun 248006, India. manpreetchhina@gmail.com
Received: 10-June-2013 • Accepted: 7-May-2014
Abstract: The greater wax moth, Galleria mellonella, is an important pest of beekeeping industry. The effects of five natural diet materials on the developmental biology and mortality of all life stages of the greater wax moth were investigated in a laboratory trial. Effect of all five natural diets on the developmental biology and mortality varied significantly. New wax comb (NW) was found to be the poorest larval diet inducing prolonged immature stage duration, shortening adult moth longevity, decreasing egg productivity, retarding oviposition, prolonging the entire life-cycle duration and causing significantly higher overall life stage mortality. In contrast, feeding larvae on diets of old wax comb (OW), old wax comb containing 10% w/w added pollen (OWP), new wax comb containing 10% w/w added pollen (NWP), and bee-collected pollen (BP), positively affected the development and vitality of all life stages. The significance of these results with respect to the control measures are proposed and discussed.
Key words: Greater wax moth. Galleria mellonella. Larval nutrition. Development. Wax comb.
Resumen: La polilla mayor de la cera, Galleria mellonella, es una plaga importante en la apicultura. Se evaluaron los efectos de cinco materiales naturales encontrados en su dieta bajo condiciones de laboratorio. Todas las dietas afectaron la biología del desarrollo así como la mortalidad, la cual varió significativamente. El panal nuevo de cera (PN) representó la dieta más pobre para las larvas al inducir una prolongada duración de la etapa inmadura, acortar la longevidad de adultos, disminuir la productividad de huevos, retardar la oviposición, alargar la duración total del ciclo de vida y, en general, inducir una mortalidad significativamente mayor. Por el contrario, la alimentación de las larvas con las otras dietas (i.e. panal viejo (PV), panal viejo con polen 10% w/w (PV + P), panal nuevo con polen 10% w / w (PN + P) y polen de abeja recogido (PA), favorecieron el desarrollo y la vitalidad de todas las etapas de desarrollo. Se discute la importancia de estos resultados a la luz de medidas de control.
Palabras clave: Gran polilla de la cera. Galleria mellonella. Nutrición larval. Desarrollo. Panal de cera.
Introduction
The greater wax moth or honeycomb moth, Galleria mellonella L. (Lepidoptera: Pyralidae) is the major destructive and economically important pest of wax comb because of their feeding habits and tunnelling through the combs (Jackman and Drees 1998; Chandel et al. 2003). The moth is widely distributed throughout the world, causing serious problems in temperate, tropical and subtropical beekeeping regions, where the warm temperature favour the rapid development of the moth (Spangler 1989). Also, the development and metamorphosis of G. mellonella have been reported to be influenced by relative humidity and diet (Abdel-Naby et al. 1983; Mohamed 1983; Gulati and Kaushik 2004). Wax moth damage only during their larval stage. It has been suggested that the larvae obtain nutrients from honey, castoff pupal skins, pollen, wax and other impurities found in the beeswax, but not the beeswax itself (Jindra and Sehnal 1989). Nevertheless, utilizing the beeswax by G. mellonella larvae was stated by Niemierko and Wlodawer (1950), thus the larvae can obtain a large amount of energy from wax, yet wax may not provide sufficient other materials e.g. protein, for successful development. Also, in the recent years, researches have focused on the economic importance of the greater wax moth due to its susceptibility to a wide range of biological control agents, for instance, entomopathogenic nematodes, viruses, fungi along with the natural enemies of predators and parasites (Dindo et al. 2001; Armendariz et al. 2002; Ueno 2002; Ansari et al. 2003; Parthasarathy and Rabindra 2003; Shapiro-Ilan et al. 2003; Tkaczuk et al. 2003; Hussaini 2003). Accordingly, mass rearing of the greater wax moth has been experimented, but mostly on artificial diets, to study various biological parameters viz, duration of developmental life stages, fertility and survival, before choosing the tested materials as suitable diets. Artificial diet composition was described and modified in previous works (Chandel et al. 2003; Coskun et al. 2006; Birah et al. 2008; Kulkarni et al. 2012; Ellis et al. 2013).
The present study was designed to investigate the efficacy of natural diets of wax comb contents on the development and mortality of G. mellonella life stages. The obtained findings could, subsequently, offer basic information for integrated pest management of this important pest.
Materials and methods
Experimental. A laboratory stock culture of G. mellonella was raised from a honey infested wax comb and maintained in complete darkness in an incubator at 30 ± 1 oC and 50 ± 5% RH in Department of Plant Protection, King Saud Uni versity Riyadh, Saudi Arabia (Abdel-Naby et al. 1983). For the present, pairs of emerged moths (male and female) were kept each in a plastic container for egg collecting. A microscope slide, covered on its upper surface with double-sided sell sellotape, was designated to collectively mount eggs deposited in the same day and placed in a Petri-dish. Daily observations were made on eggs for hatching and newly hatched larvae of the same age (≤ 24 h old) were collected. Groups of 25 larvae were picked out and placed each in 10 ml Petri-dish containing 30 g of one of the following 5 diets: (a) bee-collected pollen (BP) mixed with sucrose solution (1:1 w/w); (b) empty old wax comb aged ≥ 3 years (OW); (c) old wax comb containing 10% w/w added pollen (OWP); (d) empty new wax comb aged ≤ 2 years (NW) and new wax comb containing 10% w/w added pollen (NWP). Prior of populating Petri dishes with larvae, the dishes were exposed to 49 oC for 80 min to eliminate any associated stages of G. mellonella (Cantwell and Smith 1970).
Effect of larval nutrition on the development and mortality of life stages
Immature stages
Larval food consumption. A group of three Petri-dishes populated with larvae was designated to each tested diet and returned to the incubator. By the end of the larval duration, folded filter paper was placed in each dish; fully grown larvae that started spinning cocoons were daily observed, collected and thereafter transferred to a 12 ml. Petri-dish for pupation. When all larvae were transferred the remaining diet was separated from the solid discards (faces and silk) and weighed. The consumed diet was then determined.
Larval and pre-pupal durations. Daily counts of larvae that spun cocoons were conducted and larval duration and mortality were calculated. Through the glass Petri-dish, the pupated larvae could easily be observed (El-Sawaf 1950) and pre-pupal stage duration was subsequently assessed.
Pupal duration. For each tested diet, three groups of 5 pupae each were selected from larvae that pupated on the same day. Observations were made daily, numbers of emerged male and females were recorded, the pupal durations and percentages of emerged adult moths were then computed.
Adult stage
Adult longevity and female oviposition period. Five replicates, each comprised a pair of newly emerged adult male and female, were placed in 6 x 3 cm plastic tubes and covered with a plastic lid. A modified technique was adopted to facilitate egg laying and collecting (Mohamed 1983), by using nylon organdy wider than the tube placed on the top under the plastic lid. The little space between the tube edge and the lid proved ideal for the female to lay eggs in one plate. Deposited eggs were collected daily and counted using a binocular dissecting stereomicroscope at X12.5 magnification. The pre-oviposition, oviposition, and post-oviposition periods and female fecundity (No. of deposited eggs/female) were subsequently assessed. The adult longevities were also calculated following adult moth death.
The incubation period. The incubation period of the eggs was determined for each treatment using 5 groups of 100 eggs of one plate, deposited on the same day, mounted on a microscope slide as described above. Daily observations were made for counting hatched eggs. Average egg hatchability and incubation period were thereafter calculated.
Statistical analysis. The data collected in the above manner were subjected to suitable statistical analysis. To determine which diet was different from one another, data were statistically analysed using analysis of variance (ANOV A), followed by comparing means with the Tukey's test. A level of P ≤ 0.05 was accepted as being statistically significant. The SAS 9.1.3 program (SAS Institute 2004) was employed for conducting statistical analysis.
Results
Effect of larval nutrition on the development of life stages
Immature stages
Larval duration. Feeding newly hatched larvae on natural diets of wax comb i.e. empty old wax comb aged ≥ 3 years (OW), old wax comb with pollen (OWP), bee-collected pollen (BP), empty new wax comb aged ≤ 2 years (NW) and new wax comb with pollen (NWP); generally affected immature stage duration. Larvae finished feeding on all tested diets and progressed to pre-pupae. However, significant differences (P < 0.05) were observed in the duration of larval development of all tested diets (Table 1). The longest larval duration averaged 114.5 ± 0.6 days for larvae which had fed on NW. Larval duration decreased sharply to 22.7 ± 0.3, 27.1 ± 0.2, 30.0 ± 0.6 and 52.5 ± 0.3 days on BP, OWP, NWP and OW, respectively (P Ë 0.05). Concerning food consumption, larvae fed on NW ate most food (Table 1). Larvae fed on other diets did not differ in the amount of food they ate.
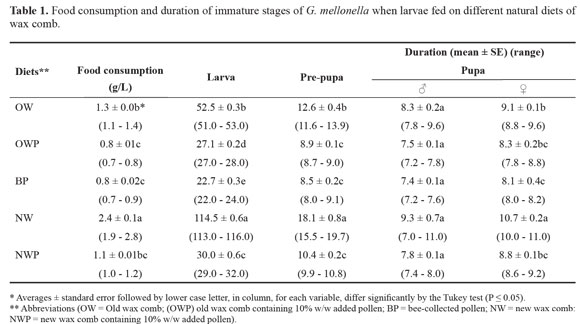
Pre-pupal duration. Similar trends were observed in the pre-pupal stage where the recorded durations were 18.1 ± 0.8, 8.5 ± 0.2, 8.9 ± 0.1, 10.4 ± 0.2 and 12.6 ± 0.4 days for the larvae fed on NW, BP, OWP, NWP and OW diets, respectively (P Ë 0.05) (Table 1) .
Pupal duration. The longest male pupal duration lasted for an average of 9.3 ± 0.7 days on NW diet, while the shortest was 7.4 ± 0.1 days on BP diet; these differences were minor and not significant. However, larvae which fed on NW diet showed a significantly longer female pupal duration (10.7 ± 0.2 days) compared to those from all other diets (Table 1). There were no significant differences detected between OW, NWP, OWP and BP diets (9.1 ± 0.1, 8.8 ± 0.1, 8.3 ± 0.2 and 8.1 ± 0.4 days, respectively (P Ë 0.05).
Adult duration. With all tested diets, larvae progressively developed to adult stages. Adult male longevity was significantly affected by larval diet (Table 2). Larvae fed on BP or OWP diets had prolonged male adult longevities (18.4 ± 0.5 and 17.2 ± 1.1 days, respectively), however they differ significantly when compared with the shortest longevity obtained when larvae were fed on NW diet (10.0 ± 0.4 days). Significant differences were seen between BP and each of OW and NWP diets.
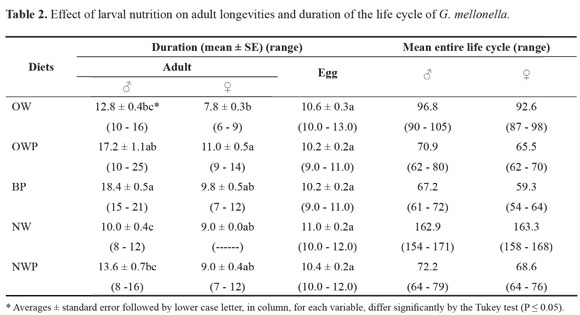
Adult female longevity was not greatly affected by larval diet. The only significant difference was observed between OWP and OW diets which exhibited the longest and shortest adult female longevities (11.0 ± 0.5 and 7.8 ± 0.3 days, respectively). Generally, on all tested diets, males survived longer than females (Table 2).
Incubation duration. Incubation period of eggs laid by adult females fed on different tested diets during larval stages was ranged from 10.2 ± 0.2 to 11.0 ± 0.2 days and did not differ significantly between treatments (Table 2).
Entire life cycle. The entire life cycle of the greater wax moth varied greatly with larval diet. Generally, with all tested diets, the female life cycle was slightly shorter than that of male (Table 2). The development of larvae and progressive life cycle stages were slowed, but not inhibited, on NW diet, where the duration of the life cycle was the longest at 162.9 and 163.3 days, for adult male and female, respectively. On the contrary, the shortest life cycle was achieved by feeding larvae on BP diet, where it lasted for 67.2 and 59.3 days for male and female, respectively.
Pre-oviposition and post-oviposition durations. Larvae fed on OWP or BP diet resulted in shortest pre-oviposition period while the longest pre-oviposition duration was recorded for female moths fed as larvae on NW diet where they started late in laying eggs when compared with those fed on any other diet (Table 3). Significant differences in the pre-oviposition periods were observed between NW and other tested diets. However, non-significant differences were noticed in post-oviposition periods of all diets (P Ë 0.05).
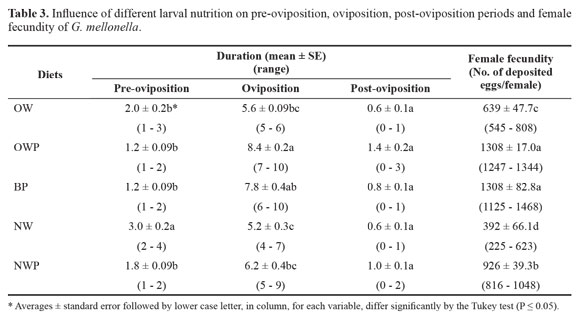
Oviposition duration. Depositing eggs lasted for a short period of 5.2 ± 0.3 days for NW diet. Significant increases in oviposition duration were noted for BP and OWP diets, which showed oviposition durations of 7.8 ± 0.4 and 8.4 ± 0.2 days, respectively (Table 3).
Female fecundity. Adult female fecundity, as measured by the mean number of deposited eggs/female, fluctuated tremendously among different tested diets (Table 3). The highest was achieved where the moths fed as larvae either on OWP or BP diet (1308 eggs/female, each). Meanwhile, adult females reared on NW diet showed the significantly lowest fecundity (392 eggs/female) followed by moths fed as larvae on OW (639 eggs/female) and on NWP (926 eggs/female) with significant differences among them (Table 3).
Daily egg laying pattern. The daily number of deposited eggs/female, as percentages of total eggs, laid, is illustrated in Fig 1. Tracing egg laying activity throughout the successive days of oviposition period, it could be clearly noticed that, for most tested diets except for NW, the proportion of deposited eggs (> 50%) were laid in the first day of oviposition, where the percentages of deposited eggs were 70.5,63.3, 54.3 and 50.1% on diets of BP, OWP, NWP and OW, respectively.
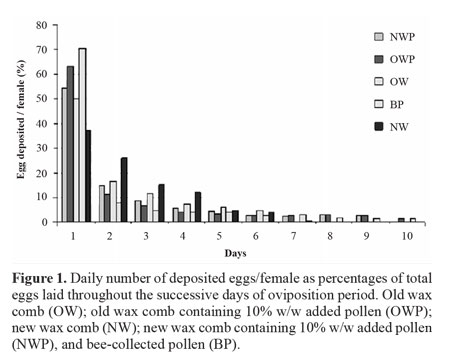
Thereafter, the percentages of deposited eggs on either BP or OWP diets decreased sharply in the 2nd day of oviposition to reach 8.7 and 11.2%, respectively. Adult females fed as larvae on NW, laid 37.1% of their eggs on the 1st day of oviposition, a gradual decrease, thereafter, occurred from the 2nd day to the end of oviposition period.
Life stages mortality
Larval mortality. Larval mortality was significantly the highest (71.7%) on NW diet followed by OW diet (48%), whereas, larvae fed on OWP showed the lowest mortality of 6.7% (Fig. 1). Nonsignificant differences were detected between OWP and either BP or NWP diets, where larval mortality were 10.7 and 14.7% for the last two diets, respectively.
Pupal mortality. The same trend was noticed for pupal mortality since the NW diet resulted in a pupal mortality significantly higher (44.3%) than those of other tested diets, followed by NWP diet (20.0%). Moreover, there were no significant differences among the percentages of the other diets. All pupated larvae on the BP diets successfully emerged to adults.
Egg mortality. Moths of G. mellonella when fed during their larval stage on NW diet, laid eggs with the lowest survival; 27.1% of the deposited eggs failed to hatch (Table 4). Significant differences in egg mortality were noticed between NW and OWP, BP, NWP or OW treatments. The percentages of unhatched eggs in theses later treatments were 1.1, 1.4, 4.2 and 4.9%, respectively (Table 4).
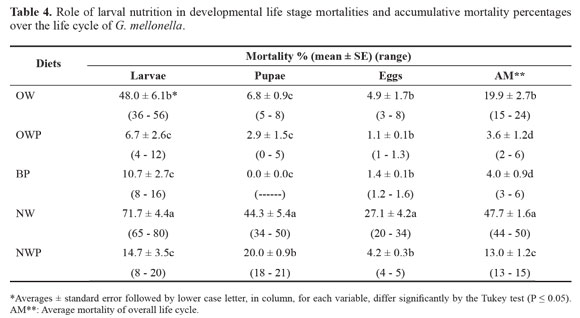
Accumulative mortality over all the life cycle. Larva fed on NW diet had the significantly highest mortality of 47.7%. Difference in mortality between OWP and BP diets was insignificant (3.6 and 4.0%, respectively). However, these percentages were significantly lower than those of NWP and OW diets (13.0 and 19.9%, respectively). The diets can be arranged in descending order according to their suitability for development of G. mellonella as follows: OWP > BP > NWP > OW > NW.
Discussion
The greater wax moth, Galleria mellonella, is a major pest of beekeeping industry (Chandel et al. 2003). The present study was conducted to explore the effects of five natural diet materials on the developmental biology and mortality of all life stages of the greater wax moth Galleria mellonella in the laboratory trial. Guazzugli and Campadell (1976) suggested that the only impact of larval diet was in relation to cholesterol; where larvae fed on zero cholesterol diet developed more slowly in the 1st generation. However, the present investigations demonstrated the influence of larval diets on the development and survival of all life stages of G. mellonella. These findings are in agreement with the observations of Chandel et al. (2003), Birah (2008) and Kulkarni et al. (2012), who reported that the investigation of detailed biology is necessary to evaluate the artificial diet modifications.
In the present study, we found a major effect of larval diet on life stage duration. Larval duration was prolonged on new wax comb diet, while on protein-rich diets i.e. pollen and old wax comb (containing cast exuviae of immature bee stages) with added pollen, it was shorter. Pant and Kapoor (1964) also found that larvae of the greater wax moth were more attracted to old beeswax rather than fresh combs and old refined beeswax because it contained essential amino acids.
Food consumption correlated with the duration of the larval stage. Larvae fed on new wax comb diet consumed more food to enable them to gain their nutritive requirements for development. It seems more likely that new wax comb was an inadequate diet, where pre-pupal and female pupal durations were significantly longer than those of other tested diets. Moreover, adult females emerged from larva fed on new wax comb showed the shortest both longevity or oviposition period and longest egg incubation period. Female fecundity was found to be significantly lower on this diet.
Generally, the entire life cycle duration reflects insufficiency of new wax comb as a diet for development. The duration of the life cycle generation on this diet was substantially longer than that any of the other tested diets. These results are consistent with those obtained by Hassanein et al. (1969) on lesser wax moth Achroia grisella reared on different diets. Mortality over all life stages was significantly higher when larvae were reared on new wax comb. On the contrary, the lowest mortality was seen on either bee-collected pollen, or old wax comb containing pollen. These findings agreed with Hassanein et al. (1969).
Active and b honey bee colonies can protect themselves from the effects of wax moths; damage caused by the greater wax moth can, however, be very serious, especially in comb stores, weakened or queenless colonies (Mangum 1989; Caron 1992). Damage can be accelerating before the beekeeper discovers these infested colonies. Since infestation in uncovered combs is substantially higher than that of fully covered combs (Nielsen and Brister, 1979 and Jouth, 1989). Care must therefore be taken during inspecting colonies, particularly during a long honey flow, or late in the fall before wintering bees to ensure active and populated colonies.
According to the present results, removal of uncovered combs is bly recommended, specifically old brood combs containing pollen (aged ≥ 3 years). Removed old combs must be melted, recycled and not be stored since brood cells of these combs became reduced in size due to the accumulation of cocoons and cast-off larval and pupal skins. Grout (1960) reported that the size of brood cells affected the size and variability of the worker bee and significantly smaller bee were obtained from small cells. Also, comb storage may increase the opportunity for the old combs to be infested with wax moth, unless all life stages of the moth are destroyed before storage. Precautions must, moreover, be maintained to ensure that no other moths can reach those combs (Cantwell et al. 1972). Consequently, for minimizing wax moth infestation, the present study suggests that wax comb within the hive must be renewed every three years.
Conclusion
The diet of new wax comb (NW) could not support the growth and development of G. mellonella successfully with most unsuitable impact on its reproductive potential. In contrast old wax comb contained pollen (OWP) and collected bee pollen (BP) positively affected the biological development parameters of all life stages.
Literature cited
Abdel-Naby, A. A.; Ata llah, M. A.; Morad, M. G.; Mohamed, A. A. 1983. Effect of different temperatures, relative humidity and light on the immature stages of the greater wax moth (Galleria mellonella L.). Proceeding of 5th Arab Pesticides Conference, Tanta University 1: 94-103. [ Links ]
Ansari, M. A.; Phan , K. L.; Moens, M. 2003. Heterorhabditis bacteriophora (Heterorhabditidae: Rhabditida) parasitic in natural population of white grubs (Coleoptera: Scarabaeidae) in Belgium. Russian Journal of Nematology 11: 57-59. [ Links ]
Armendariz, I.; Down es, M. J.; Griff in, C. T. 2002. Effect of timber condition on parasitization of pine weevil, (Hylobius abietis L.) larvae by entomopathogenic nematodes under laboratory conditions. Biocontrol Science and Technology 12: 225-233. [ Links ]
Birah, A.; Chilana , P.; Shuk la, U. K.; Gupta , G. P. 2008. Mass rearing of greater wax moth (Galleria mellonella) on artificial diet. Indian Journal of Entomology 70: 389-392. [ Links ]
Cantwell, G. E.; Smith, L. J. 1970. Control of the greater wax moth (Galleria mellonella L.) in honey comb and comb honey. American Bee Journal 110(4): 141. [ Links ]
Cantwell, G. E.; Jay, E. G.; Rearman , Jr. G. C. Thompsons, J. V. 1972. Control of the greater wax moth Galleria mellonella (L.) in comb honey with carbon dioxide. American Bee Journal 112 (8): 302-303. [ Links ]
Caron, D. M. 1992. Wax moth. American Bee Journal 132 (10): 647-649. [ Links ]
Chandel, Y. S.; Sharma S.; Verma, K. S. 2003. Comparative biology of the greater wax moth, Galleria mellonella L., and lesser wax moth, Achoria grisella. Forest Pest Management and Economic Zoology 11: 69-74. [ Links ]
Coskun , M.; Kayis, T.; Su lanc , M.; Ozalp, P. 2006. Effects of different honeycomb and sucrose levels on the development of greater wax moth Galleria mellonella larvae. International Journal of Agriculture and Biology 8 (6): 855-858. [ Links ]
Dindo, M. L; Verdinelli, M.; Ba ronio, P.; Serra, G. E. 2001. Laboratory and field performance of in vitro and in vivo reared Exorista larvarum (L.), a natural enemy of cork oak defoliators. In: Villemant, C.; Sousa, E. (Eds.). Integrated protection in oak forests. Proceedings of the IUBC-WPRS working group at Oeiras Lisbonee, Portugal, 01st - 04th Oct., 2001. Bulletin OILB - SRO P 25: 147-150. [ Links ]
ELLIS , J. D.; Graham, J. R.; Mortensen, A. 2013. Standard methods for wax moth research. Journal of Apicultural Research 52 (1): DOI 10.3896/IBR A.1.52.1.10 [ Links ]
El-sawaf , S. K. 1950. The life history of the greater wax moth (Galleria mellonella L.) in Egypt, with special reference of morphology of the mature larvae. Bulletin of the Entomological Society of Egypt 34: 247-297. [ Links ]
Grout , R. A. (Ed.). 1960. The hive and the honey bee. Hamilton, IL , USA; Dadant & Sons 3rd ed. 740 p. [ Links ]
Guazzug li, M. A.; Campa delli, G. 1976. Prove di selezione di popolazioni non soggette a diapausa del Lepidottero Galleria mellonella L. Bollettino dell'Istituto di Entomologia dell'Universita degli Studi di Bologna 32: 153-168. [ Links ]
GulaTi, R.; Kau shik, H. D. 2004. Enemies of honeybees and their management- a review. Agricultural Reviews 25 (3): 189- 200. [ Links ]
Hussaini, S. S. 2003. Progress of research work on entomopathogenic nematodes in India. pp. 27-69. In: Hussaini, S. S.; Rabindra, R. J.; Nagesh, M. (Eds.). Current Status of Research Entomopathogenic Nematodes in India. PDB C, publication, Bangalore, India. [ Links ]
Hassan ein, M.; Ibrahim, M. H.; El-Ban by, M. A.; El- Arousy, A. F. M. 1969. Effect of different diets on some biological aspects of the lesser wax moth, Achroia grisella Fab. Lepidoptera: Galleriidae. Bulletin of the Entomological Society of Egypt 53: 567-572. [ Links ]
Jackman , J. A.; Drees, B. M. 1998. A field guide to Texas insects, Gulf Publishing Company, Houston, Texas. [ Links ]
Jindra, M.; Sehna l, F. 1989. Larval growth, food consumption and utilization of dietary protein and energy in Galleria mellonella. Journal of Insect Physiology 35 (9): 719-724. [ Links ]
Jout h, J. V. A. 1989. Ecological and physiological studies on the greater wax moth Galleria mellonella L. Ph. D. Thesis, Bangolore University, India XI 180 p. [ Links ]
Kulkarni, N.; Kushwa ha, D.K.; Mishra, V.K.; Paun ikar, S. 2012. Effect of economical modification in artificial diet of greater wax moth Galleria mellonella (Lepidoptera: Pyralidae). Indian Journal of Entomology 74 (4): 369-374. [ Links ]
MaNGum, W. 1989. Early methods of wax moth control. American Bee Journal 129 (10): 30-33. [ Links ]
Mohamed, A. A. 1983. Biological and ecological studies on the greater wax moth (Galleria mellonella L.). M. Sc. Thesis, Faculty of Agriculture, Minia University, Egypt 97 p. [ Links ]
Nielsen, R. A.; Brister, C. D. 1979. Greater wax moth: 1. Behavior of larvae. Annals of Entomological Society of America 72: 811-81 [ Links ]
Niemierko, W.; Wlodaw er, P. 1950. Studies in the biochemistry of the greater wax moth (Galleria mellonella L.). 2. Utilization of wax constituents by the larvae. Acta Biologiae Experimentalis 15: 60-78. [ Links ]
Pant , N. C.; Kap oor, S. 1964. A note on the nutritional behavior of the larvae the greater wax moth Galleria mellonella L. (Lepidoptera: Pyralidae). India Journal of Entomology 26: 125-127. [ Links ]
ParAthasarAthy, R.; Ra bindra, R. J. 2003. Pathogenicity of Galleria mellonella nucleo polyhydro virus and Plutella xylostella granulovirus and their combination to diamondback moth (Lepidoptera: Plutellidae). Indian Journal of Plant Protection 31: 1-4. [ Links ]
SAS In stitut e. 2004. The SAS System Version 9.1.3. SAS Institute, Cary, NC. [ Links ]
Shapiro-Ilan , D. I.; Gardner, W. A.; Fuxa , J. R.; Wood, B. W.; Ngu yen, K. B.; Adams, B. J.; Hu mber, R. A.; Ha ll, M. J. 2003. Survey of entomopathogenic nematode and fungi endemic to pecan orchards of the south-eastern United States and their virulence to the pecan weevil (Coleoptera: Curculionidae). Environmental Entomology 32: 187-195. [ Links ]
Spangler, H. G. 1989. The role of ultrasound and pheromone communication of greater and lesser wax moths. Bee World 70 (3): 132-133. [ Links ]
Tkaczuk , C.; Renella, G.; Pap ierok, B. 2003. Occurrence of entomopathogenic fungi in soils from central Italy under different managements. In: Papierok, B. (Ed.). Entomopathogens and insect parasitic nematodes: current research and perspectives in pest biocontrol. Proc. 8th European Meet. IOB C-WPRS Working Group Insect Pathogens and Insect Parasitic Nematodes, Athens, Greece, 29th May - 2 June, 2001. Bulletin-OILB - SRO P 26 (1): 85-89. [ Links ]
Ueno, T. 2002. Biology of the ectoparasitoid wasp, Agrothereutes lanceolatus (Hymenoptera: Ichneumonidae) host acceptance and larval development on a laboratory host. Journal of the Faculty of Agriculture Qushu University 47: 37-43. [ Links ]
Suggested citation:
MOHAMED , ABEDELSALAM ANWAR, MOHAMED JAVED ANSARI , AHMAD AL-GHAMDI , MOHAMED OMAR MOHAMED and MANPREE T KAUR. 2014. Effect of larval nutrition on the development and mortality of Galleria mellonella (Lepidoptera: Pyralidae). Revista Colombiana de Entomología 40 (1): 49-54. Enero-julio 2014. ISS N 0120-0488.
