Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.2 Bogotá July/Dec. 2014
SECCIÓN MÉDICA / MEDICAL
Análisis sucesional de Calliphoridae (Diptera) en cerdo doméstico en pastizales (Cogua, Cundinamarca, Colombia)
Successional analysis of Calliphoridae (Diptera) using domestic pig in grassland (Cogua, Cundinamarca, Colombia)
Andrés F. Sánchez RestrepoI; Giovany FaguaII
IBiólogo. Laboratorio de Entomología, Pontificia Universidad Javeriana. Carrera 7 No. 43 - 82. Bogotá, Colombia. andres.sanchez@javeriana.edu.co. Autor correspondencia
IIBiólogo, M. Sc., Profesor Asociado, Laboratorio de Entomología, Pontificia Universidad Javeriana. Carrera 7 No. 43 - 82. Bogotá, Colombia. fagua@javeriana.edu.co
RESUMEN
La identificación de especies de insectos encontradas en cadáveres es el primer paso que la entomología forense tiene en cuenta en la estimación del intervalo post-morten. Los Calliphoridae constituyen una porción importante de las especies de insectos presentes en investigaciones forenses. Este estudio evaluó la sucesión de especies de califóridos asociadas a cuerpos en descomposición de cerdos (Sus scrofa) en pastizales de Cogua (Cundinamarca). Los dípteros fueron colectados durante siete meses posteriores al depósito de los cuerpos utilizando trampas tipo Schoenly modificadas y el método tradicional. Se colectaron 970 individuos de cinco especies de califóridos, siendo las más comunes Compsomyiops verena y Chrysomya albiceps. Las variaciones en las abundancia de cada especie coincidieron con cada una de las etapas de descomposición, encontrando las mayores abundancias al inicio y final de la etapa de hinchado. Al comparar los métodos se encontró que las trampas Schoenly presentaron mayor eficiencia de colecta de adultos; sin embargo, mediante la metodología tradicional se encontró la mayoría de los estadios inmaduros. Se confirma la importancia de los califóridos como grupo adecuado para el establecimiento del intervalo post morten debido a que la secuencia de las especies ayuda a delimitar las diferentes etapas de descomposición cadavérica. Además se confirma la eficacia y complementariedad del uso de las trampas Schoenly en este tipo de estudios.
Palabras clave: Entomología forense; Intervalo post-mortem; Descomposición cadavérica; Fauna sarcosaprófaga.
ABSTRACT
The first step to estimate the post-mortem interval is the identification of species of insects found on carcasses. Blowflies are an important portion of the insect species in forensic researches. Here, we evaluated the succession of blowfly species associated with pig carcasses (Sus scrofa) on pastures of Cogua (Cundinamarca). The blowflies were collected during seven months after the deposit of carcasses using Schoenly modified traps and the traditional methodology. We collected 970 specimens of five blowfly species, where the most common species were Compsomyiops verena and Chrysomya albiceps. The variations in abundance per species were coincident with each decay stages; we found the highest abundances at the beginning and at the end of the bloated stage. When we contrasted the results of both methodologies, we found that Schoenly traps presented the highest efficacy of adult's collection in comparison with the traditional methodology; however, we collected the majority of immature stages using the traditional methodology. The importance of blowflies as an adequate group to obtain the post-mortem interval was confirmed; the sequence of species helps to delimit the different stages of the cadaveric decomposition. Additionally, we confirmed the efficacy and complementary of the Schoenly trap in this kind of forensic studies.
Key words: Forensic Entomology; Post-mortem interval; Cadaver decomposition; Sarcosaprophagous Fauna.
Introducción
La identificación de especies de insectos encontrados en asociación con cadáveres es generalmente el primer paso que tiene en cuenta la entomología forense en el intento de estimación del intervalo post morten (Goff y Odom 1987; Anderson 2000), es decir el tiempo que transcurre desde la muerte de un organismo hasta el hallazgo de su cuerpo sin vida. Esta herramienta ha sido la de mayor uso en la medicina forense e implica la definición del proceso de sucesión de la fauna cadavérica mediante el seguimiento de la descomposición de un modelo biológico internacionalmente definido, un cadáver de cerdo (Sus scrofa Linnaeus, 1758), cuyo proceso de descomposición es monitoreado mediante la recolección de los insectos asociados al cadáver.
Conocer cuáles especies de insectos se encuentran presentes y en qué momento, permite identificar las fases de arribo, el tiempo de uso del cadáver y partida de las diferentes especies de la fauna sarcosaprófaga. Además, facilita dilucidar posibles vínculos sobre las circunstancias de muerte o establecer si el cadáver fue traslado desde uno a otro sitio (Byrd y Castner 2010). Para los insectos el cadáver representa un recurso de reciente disponibilidad que sobrelleva un proceso de colonización inicial por moscas califóridas y algunos coleópteros (como sílfidos y derméstidos; Barreto et al. 2002), que hacen uso inmediato del recurso (el cadáver), y los que excluyen o facilitan la penetración a éste de otros organismos en oleadas sucesionales (Connel y Slayer 1977).
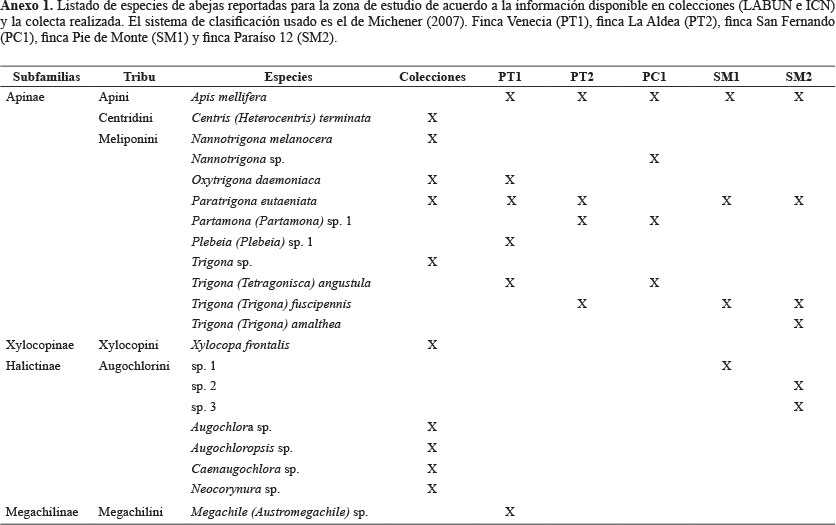
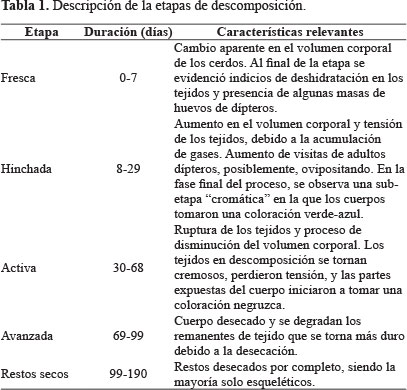
Los cadáveres son recursos efímeros donde los primeros colonizadores poseen un efecto significativo sobre la estructura y composición final de la comunidad, por su intensidad de consumo y su habilidad de eliminación competitiva (Ordoñez 2004). Los cambios físico-químicos que sufre un cadáver durante el proceso de descomposición genera que diferentes especies se especialicen en distintas etapas de descomposición, generando una sucesión faunística temporal que, una vez conocida, puede ser previsible (Arnaldos et al. 2006). Este cambio temporal permite distinguir diferentes etapas sucesionales o seriales cuyo número puede variar de las cuatro etapas propuestas por Mégnin (1884) o las tres de Fuller (1934). Hasta el momento, el modelo de Bornemissza (1957) es el más seguido pues mostró ser semejante para todas las regiones europeas (Nuorteva 1977) y ha venido siendo utilizado por buena parte de los estudios tanto en regiones paleárticas, como neárticas, tropicales africanas y neotropicales (Ordoñez et al. 2008). Bornemissza (1957) señala cinco etapas de la descomposición de un cadáver: 1) fresca, 2) hinchada, 3) descomposición activa, 4) descomposición avanzada y 5) restos secos.
La secuencia de arribo de artrópodos y otros organismos invertebrados depende tanto de las condiciones ambientales como de la composición de la fauna local (Payne 1965; Catts y Goff 1992); el poder determinar el efecto de estas dos variables y su relación con el proceso de sucesión es uno de los principales objetivos al tratar de definir un intervalo post mortem. Entre los insectos que constituyen una porción importante de la fauna cadavérica se encuentran las moscas de la familia Calliphoridae. Estas suelen ser las primeras especies de insectos en colonizar el cadáver y participan activamente en su proceso de descomposición (Wolff et al. 2001; Segura et al. 2005). Son de hábitos principalmente descomponedores, sarcosaprófagos y/o coprófagos (Amat et al. 2008); las hembras depositan sus huevos sobre los cadáveres frescos permitiendo que las larvas se desarrollen usando enzimas proteolíticas para poder penetrar el cadáver. Generalmente, la tasa de crecimiento y número de la población de estas moscas es directamente proporcional al tiempo de descomposición del cadáver, lo cual ayuda a estimar el intervalo post-morten (Goff y Odom 1987).
Dadas las variadas condiciones ambientales y topográficas en Colombia, la fauna cadavérica cambia dependiendo del hábitat, vegetación, tipo de suelo y variables ambientales (humedad y temperatura). Esta variación afecta la descomposición de los restos, haciéndola particular para cada zona, al igual que la forma en que se da la sucesión de la fauna cadavérica (Anderson 2004; Arnaldos et al. 2006). En el país se han realizado varios estudios que reportan la sucesión de la fauna cadavérica usando cerdos como modelo; entre estos se destacan los desarrollados en Medellín (Wolff et al. 2001; Pérez et al. 2005), Bogotá (Camacho 2005; Segura et al. 2009), en ecosistemas acuáticos (Barrios y Wolff 2011), en ecosistemas de páramo (Martínez et al. 2007) y en la región cafetera (Grisales et al. 2010). Adicional a estos se han realizado estudios que analizan los ciclos de vida de varias especies de dípteros de importancia forense (Camacho y Usaquén 2004; Camacho 2005; Segura et al. 2005; Vélez y Wolff 2008).
Por otro lado, la mayoría de estudios realizados han empleado el método tradicional de colecta, que consiste en realizar una inspección del cadáver con pinzas y jama. Sin embargo, este procedimiento puede ser subjetivo en cuanto depende de la habilidad del colector, de la intensidad del muestreo y, en la mayoría de los casos, implica manipular el cadáver. Para solucionar esta deficiencia se propuso el uso de trampas que tienen un proceso permanente de colecta de los organismos sarcosaprófagos (Schoenly 1981; Schoenly y Reid 1987; 1989; Ordoñez 2004); esto obvia el problema de las condiciones microclimáticas del momento de la colecta, la habilidad del colector y tampoco requiere de remover el cadáver (Ordoñez et al. 2008). Para Colombia, Ordoñez et al. (2008) proponen las trampas Schoenly modificadas con el uso de conejos como cebo, en las cuales se registra la mayor eficiencia en cuanto a la abundancia y número de especies colectadas respecto a los métodos tradicionales (principalmente colecta manual). A partir de estos estudios, el presente trabajo busca evaluar la sucesión de especies de califóridos asociadas a cuerpos en descomposición del modelo biológico internacionalmente definido en estudios forenses (Sus scrofa) en pastizales de Cogua (Cundinamarca), comparando el método tradicional y trampas tipo Schoenly modificadas. Estas trampas modificadas, inicialmente propuestas por Schoenly (1981), podría resultar ser más promisoria y versátil permitiendo generar un protocolo estandarizado y menos dependiente del colector y del evento de colecta.
Materiales y métodos
Sitio de estudio. El material base para la realización del trabajo fue obtenido del proyecto "Corroboración de la eficiencia del uso de trampas Schoenly en la definición del intervalo postmortem: Modelo Sus scrofa" realizado por el laboratorio de entomología de la Pontificia Universidad Javeriana. Este proyecto se desarrolló en la Finca Experimental San Javier (05º03'37,0"N 073º56'41,6"O; 2590 m) ubicada en el municipio de Cogua (Cundinamarca, Colombia). El sitio se caracteriza por poseer pastizales usados en actividades de ganadería de leche. Hace parte de la altiplanicie cundiboyacense con temporadas secas y lluviosas que se alternan durante todo el año, con temperaturas, generalmente, menores de 20 ºC. Estos pastizales se suelen inundar con facilidad debido a las frecuentes lluvias y a la cercanía del río Neusa.
Fase de campo. Las moscas fueron colectadas durante siete meses posteriores al depósito de los cuerpos utilizando trampas tipo Schoenly modificadas y el método tradicional siguiendo el modelo de trampa propuesto por Ordoñez et al. (2008) (Fig. 1A). Se definieron tres unidades, con seis réplicas por tratamiento: A) Trampas tipo Schoenly cebadas con cadáveres de cerdo (Sus scrofa); B) Cadáveres de cerdo dispuestos al descubierto sobre una reja metálica y cubiertos por una jaula, según el método tradicional; y C) Trampas Schoenly vacías utilizadas como control para medir la eficacia de colecta y distinguir la fauna incidental de aquella asociada al cadáver.
Los cerdos, de tres meses y con peso promedio de 18,75 kg, fueron sacrificados mediante un golpe en la cabeza y puñalada en el corazón. El sacrificio se realizó a mediados de marzo de 2012. Las observaciones y colectas se efectuaron hasta que los restos secos de todos los cadáveres de los cerdos ya no contenían material orgánico en descomposición, proceso que se completó en septiembre del mismo año de sacrificio. Los muestreos (eventos de colecta) se realizaron cada cuatro horas durante las primeras 48 horas, a diario durante el primer mes, cada 48 horas durante el segundo mes, cada 72 horas para el tercer mes, y semanalmente desde el cuarto hasta el séptimo mes. Para todos los tratamientos las muestras fueron conservadas en viales de vidrio con alcohol al 96% y sus correspondientes etiquetas de identificación. En las trampas Schoenly las muestras, de cada evento de colecta, correspondieron a todos los insectos capturados en los frascos colectores de la trampa Schoenly (Fig. 1A), tanto para el tratamiento "con cerdo" como "sin cerdo". En las jaulas las muestras cada evento de colecta correspondió a todos los insectos presentes en el cadáver, capturados de forma manual con pinzas y red entomológica durante 15 minutos. En total se obtuvieron 6700 muestras provenientes de los tres tratamientos. A partir del segundo mes todas las colectas se realizaron en horas de la mañana, entre las 09:00 y 11:00 A.M. Se registraron datos de temperatura y humedad ambiental durante todo el muestreo usando Dataloggers (modelo Hobo U12 Temp/RH).
Fase de laboratorio. El material fue analizado e identificado en el laboratorio de entomología de la Pontifica Universidad Javeriana. La mayoría del material se conservó en viales con alcohol al 96% y se realizaron montajes en seco de cada sexo representativos de cada especie. Para la identificación se emplearon las claves de Amat et al. (2008) para adultos y las de Flórez y Wolff (2009) para inmaduros. El material fue corroborado con la colección de referencia del Museo Javeriano de Historia Natural Lorenzo Uribe S.J. de la Pontificia Universidad Javeriana (MPUJ).
Se utilizó estadística descriptiva para evaluar abundancia de cada especie de califóridos capturados durante el muestreo en relación con las etapas de descomposición observadas y cada uno de los tratamientos. Con el fin evaluar si la composición y abundancia de especies presentaba alguna asociación con las etapas de descomposición se empleó la técnica un análisis multivariado de componentes principales (PCA) empleando el programa Past v2.17 (Hammer et al. 2001). Para calcular la similitud entre los muestreos de acuerdo a las diferentes etapas de descomposición se utilizaron análisis de agrupamiento de Bray-Curtis, mediante ligamiento por promedio de grupos, tomando como valor critico el 70% de similaridad para los dos casos con el empleo del programa Biodiversity Pro (McAleece et al. 1997). Para establecer si existía diferencias entre los tratamientos se realizó una prueba de Leveve para la homogeneidad de varianzas y una prueba de normalidad (Kolmogorov-Smirnov), y en caso de no cumplir con los supuestos de normalidad una prueba de Kruskal- Wallis; todas estas pruebas realizadas con el programa SPSS (IBM 2012).
Resultados
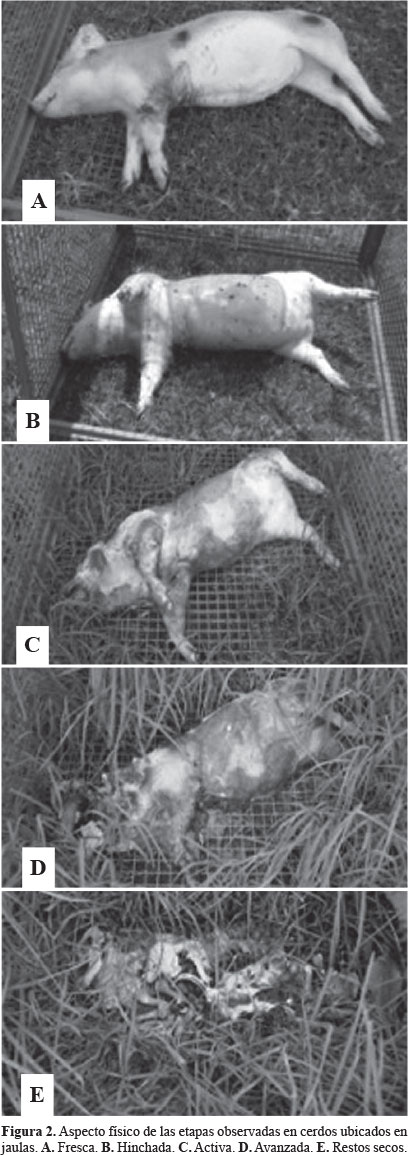
Descomposición de los cerdos. El proceso de descomposición de los cerdos fue seguido mediante registro fotográfico (Fig. 2) que permitió hacer una diferenciación de las etapas de acuerdo a las características físicas como tensión de los tejidos, color de la piel, cambio del volumen corporal (causada por los gases generados por las bacterias detritívoras) y consistencia de los tejidos (Tabla 1). Esta diferenciación sirvió como base para los posteriores análisis y comprobar si efectivamente las características físicas del recurso afectaban la composición de las especies de moscas califóridas.
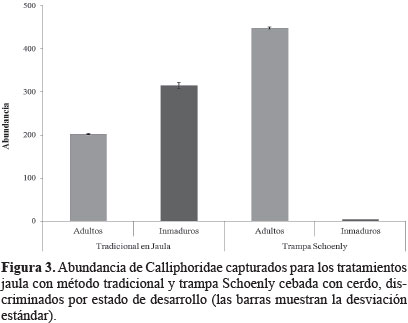
Composición y abundancia de los Calliphoridae. De las muestras obtenidas 453 tenían califóridos, para un total de 970 individuos. De las trampas Schoenly con cerdo se obtuvieron 452 individuos en dónde el 99,1% correspondía a individuos adultos y menos del 1% a inmaduros (Fig. 3). En las jaulas con metodología tradicional se obtuvieron 518 individuos en dónde el 40% correspondía a individuos adultos y 60% a inmaduros (Fig. 3). Ninguno individuo de la familia fue capturado por alguna de las trampas Schoenly sin cebo. En adelante, cuando se mencione trampa Schoenly, se referirá únicamente las trampas Schoenly cebadas con cerdo.
Las especies encontradas fueron: Calliphora nigribasis Macquart, 1851, Calliphora vicina Robineau-Desvoidy, 1851, Chrysomya albiceps (Walker, 1849), Compsomyiops verena (Walker, 1849) y Sarconesiopsis magellanica (Le Guillou, 1842). Al analizar la abundancia de cada especie durante todo el tiempo de descomposición de los cerdos se encontró que las más abundantes y relativamente constantes durante todo el estudio fueron C. verena y C. albiceps. La presencia de todas las especies durante todo el tiempo de descomposición no fue constante, presentándose solo en algunos momentos del proceso (Fig. 4). Adicional, se observaron dos aumentos representativos en la abundancia de estas especies al inicio y final de la etapa de hinchado, seguido de un decrecimiento progresivo durante las etapas de avanzada y no hubo presencia alguna de estas especies en la etapa de restos secos.
La matriz de ocurrencia (Anexo 1) mostró que las primeras especies en colonizar el cadáver fueron Sarconesiopsis magellanica, Chrysomya albiceps y Comsomiops verena. Mientras Calliphora vicina solo se presentó en la etapa de hinchado. Calliphora nigribasis solo estuvo presente en las etapas de hinchado y activa. Sarconeiopsis magellanica permaneció durante las etapas fresca, hinchado y activa. Ninguna se encontró presente en la etapa de restos secos.
Al realizar un análisis de similitud de Bray-Curtis (Fig. 5) se observó que si bien se generaron agrupaciones por encima de 70% de similitud, no se evidencia agrupaciones consistentes de cada una de las etapas. Igualmente, en el análisis de componentes principales (PCA, fig.6) no se presentaron agrupamientos claros, aunque se evidenció una tendencia a organizarse de acuerdo a las etapas de descomposición, en especial una tendencia de asociación de la mayor abundancia de C. verena y C. albiceps con la etapa de hinchado.
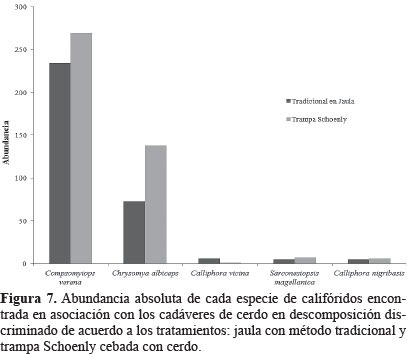
Eficiencia de las metodologías de colecta. El hecho que no se capturaran individuos de califóridos en las trampas Schoenly vacías, nos permite asumir que los califóridos colectados no fueron fauna incidental. Los tratamientos de Schoenly y Jaula con metodología tradicional capturaron las mismas especies, ninguna fue exclusiva. Sin embargo, al analizar la abundancia de individuos adultos e inmaduros capturados en cada una de las trampas, encontramos que la mayor cantidad de adultos fue capturada por medio de trampas tipo Schoenly y la mayor cantidad de inmaduros (huevos y larvas) fue capturada por medio del método tradicional (Fig. 3). Al discriminar por especies (Fig. 7) se encontró la trampa Schoenly presentó mayor abundancia en todas las especies excepto en C. vicina. Esto evidencia una diferencia, aproximada, al doble de las capturas para C. albiceps, pero que al presentar valores de abundancia bajos no se podría tomar como un patrón que refleje el comportamiento de la especie en la frecuencia de captura.
Los datos no presentaron homogeneidad de varianzas (Levene = 0,003; P < 0,05) ni se distribuyeron normalmente (Kolmogorov-Smirnov = 0.00; P < 0,05), por lo que se procedió a realizar una prueba de Kruskal-Wallis de muestras independientes que constató que no había diferencias significativas en cuanto al número de individuos colectados entre los métodos utilizados (Kruskal = 0,639; P < 0,05).
Discusión
Este estudio permitió conocer y establecer las especies de califóridos que colonizaron la descomposición de los cerdos estudiados. Aunque en los últimos años han aumentado los estudios sobre la sucesión de la fauna sarcosaprófaga en diferentes localidades del país (Camacho 2005; Segura et al. 2009; Segura et al. 2005; Wolff et al. 2001; Ordoñez 2004; Pérez et al. 2005; Ordoñez et al. 2004; Barrios y Wolff 2011), sigue siendo necesario realizar estudios que describan las dinámicas temporales y procesos sucesionales en cadáveres de cerdos como modelo biológico, los cuales son dinámicos, varían localmente y se encuentran sujetos a la influencia de un amplio espectro de procesos ambientales (Mavárez-Cardozo et al. 2005).
En el proceso de descomposición de los doce cerdos se pudo identificar cada una de las fases utilizando las características físicas de los cadáveres, ajustadas a las fases propuestas por Bornemissza (1957). El proceso de descomposición desde la muerte hasta que los restos secos de todos los cadáveres de los cerdos, sin material orgánico en descomposición, duró 160 días en promedio para todos los cerdos, casi el doble a lo registrado por Segura et al. (2009) en sectores semi-rurales cercanos a la ciudad de Bogotá. Esta diferencia puede asociarse con las bajas temperaturas nocturnas registradas al inicio del estudio y con las fuertes y frecuentes lluvias ocurridas durante la mayor parte del muestreo, lo que posiblemente alargó la duración de la etapa de fresco. Posiblemente esto puede haber generado una disminución de la actividad de descomposición a nivel microbiológico y, por lo tanto, retrasar los cambios físicos propios de la etapa fresca a la de hinchado. Nuestros resultados son aún más interesantes si se compara este trabajo con el de Martínez et al. (2007), quienes reportan una duración de 83 días de descomposición de cadáveres de cerdo en ambientes de páramo. Por otro lado, se ha encontrado que los patrones de lluvia y el agua depositada son factores que retardan la descomposición cadavérica, debido a que la temperatura de los restos disminuye el doble de rápido que en el aire, y en muchos casos el agua causa la muerte de pupas y larvas por ahogamiento (Smith 1986; Mavárez-Cardozo et al. 2005; Grisales et al. 2010). Justo, los pastizales de la sabana de Bogotá cercanos a ríos y cuerpos de agua tienen a inundarse en épocas de lluvia, como sucede en la Finca Experimental San Javier. Este evento ocurrió durante el desarrollo del estudio, a mediados de la etapa de hinchado (días 15 al 24 de descomposición), momento en el que se presentaron lluvias que anegaron el sitio, lo que muy posiblemente contribuyó a el retardo de la descomposición de los cerdos. Incluso esto podría explicar por qué la abundancia de todas las especies decrece a mediados de esa etapa (Fig. 4), coincidiendo con el periodo de mayor precipitación.
Los califóridos han sido reportados como unos de los primeros colonizadores de cuerpos en descomposición (Segura et al. 2005; Byrd y Castner 2001; Wolff et al. 2001) y, generalmente, conforman una porción importante de la fauna sarcosaprófaga (Pérez et al. 2005). En este estudio no son una excepción y conforman un aproximado de 20% de los dípteros capturados durante el estudio, que sumado a su gran abundancia en las muestras, confirman su importancia y valor al momento de servir como referentes para calcular el intervalo post morten (IPM).
En cuanto a las especies de moscas califóridas, ya se habían reportado las especies registradas en este estudio para zonas cercanas a Bogotá. Sin embargo, la secuencia de colonización y las ocurrencias difieren. Camacho (2005), en su estudio realizado en el área urbana de Bogotá, reporta a Calliphora vicina como una de las primeras especies colonizadoras. Por el contrario, Segura et al. (2005; 2009), en áreas semi-rurales de Bogotá, reportaron a Sarconesia magellanica y Compsomyiops verena como las primeras colonizadoras, coincidiendo parcialmente con el presente estudio. Esta diferencia se puede explicar por una posible estacionalidad en las especies de califóridos de acuerdo a lo mencionado por Segura et al. (2005) pero, más probable, por las diferencias en composición que presenta la comunidad de califóridos en cada hábitat en dónde se han realizado este y los otros estudios.
Diferentes trabajos han definido a C. vicina como especie predominante de ambientes urbanos (Pinilla-Beltrán et al. 2012) y señalan que su abundancia y frecuencia disminuyen en ambientes rurales (Souza y Zuben 2012; Battán-Horenstein et al. 2012; Lambiase y Camerini 2011). Este aspecto refuerza la importancia de realizar estudios en lugares con condiciones diferenciales, que permitan generar una línea de base mucho más objetiva y establecer el IPM con mayor precisión (Arnaldos et al. 2004). La abundante presencia de C. verena y C. albiceps puede deberse a que ambas son exóticas y suelen ser más abundantes que las especies nativas en ambientes transformados (Guimaraes et al. 1978; Byrd y Castner 2010), pero eso nos las excluye de su relevancia para el cálculo del IPM, como se pudo comprobar en este estudio.
Los incrementos de la abundancia de C. verena y C. albiceps corresponden con importantes eventos de cambios en el estado en descomposición de los cerdos. Comportamiento que resalta la importancia de contabilizar la abundancia de cada una de las especies de califóridos observadas durante el proceso de descomposición. Los tratamientos mostraron diferencias en cuanto a la abundancia de capturas por cada uno, pero sin alcanzar diferencias estadísticas. Ordoñez et al. (2008) evidenciaron la eficiencia de la captura de dípteros sarcosapróficos en trampas Schoenly, pero usando conejos como cebo. En este caso pudimos evidenciar el potencial de la trampa para la captura de estos insectos usando como cebo cerdos. En cuanto a la diferencia presentada entre adultos e inmaduros, dónde se encontraron la mayoría de los estadios inmaduros por método tradicional y la mayoría de los adultos por medio de las trampas, posiblemente se deba a que el diseño de la Schoenly es más específico para insectos voladores pero son necesarios experimentos adicionales para corroborar esta observación.
Cada sitio tiene una fuerte influencia en la composición y colonización de las especies de califóridos asociados a cadáveres; para el caso de la sabana de Bogotá, un factor adicional a tener en cuenta son las lluvias y exposición al agua que retrasan el proceso de sucesión cadavérica de la fauna de insectos sarcosaprófagos. Además, este trabajo confirma el potencial y complementariedad del uso de las trampas Schoenly en estudios forenses, en particular para la captura de insectos adultos.
Agradecimientos
A la Vicerrectoría de Investigación de la Pontificia Universidad Javeriana y Colciencias (Proyectos: 120352128541 y P-2011-00168) por el apoyo financiero del proyecto. A José María Vásquez Alzate, Oscar y Gina de la Finca San Javier. A los miembros del laboratorio de entomología de la Pontificia Universidad Javeriana por su constante apoyo y asesoría.
Literatura citada
AMAT, E.; VÉLEZ, M. C.; WOLFF, M. 2008. Clave ilustrada para la identificación de los géneros y las especies de califóridos (Diptera: Calliphoridae) de Colombia. Caldasia 30 (1): 231-244. [ Links ]
ANDERSON, G. S. 2000. Minimum and maximum development rates of some forensically important Calliphoridae (Diptera). Journal of Forensic Sciences 45 (4): 824-832. [ Links ]
ANDERSON, G. S. 2004. Determining time of death using blow fly eggs in the early postmortem interval. International Journal of Legal Medicine 118 (4): 240-241. [ Links ]
ARNALDOS, M. I.; GARCÍA, M. D.; ROMERA, E.; PRESAA, J. J.; LUNA, A. 2004. Estimation of postmortem interval in real cases based on experimentally obtained entomological evidence. Forensic Science International 149: 57-65. [ Links ]
ARNALDOS, M. I.; PRADO E CASTRO, C.; PRESA, J.J.; LÓPEZ-GALLEGO, E.; GARCÍA, M. D. 2006. Importancia de Los estudios regionales de fauna sarcosaprófaga - Aplicación a la práctica forense. Ciencia Forense 8 (1): 63-82. [ Links ]
BARRETO, M; BURBANO, M. E; BARRETO, P. 2002. Flies (Calliphoridae, Muscidae) and beetles (Silphidae) from human cadavers in Cali, Colombia. Memórias do Instituto Oswaldo Cruz 97 (1): 137-138. [ Links ]
BARRIOS, M.; WOLFF, M. 2011. Initial study of arthropods succession and pig carrion decomposition in two freshwater ecosystems in the Colombian Andes. Forensic Science International 212 (1-3): 164-72. [ Links ]
BATTÁN-HORENSTEIN M., ROSSO, B.; GARCÍA, M. D. 2012. Seasonal structure and dynamics of sarcosaprophagous fauna on pig carrion in a rural area of Cordoba (Argentina): Their importance in forensic science. Forensic Science International 217 (1): 146-156. [ Links ]
BORNEMISSZA, G. M. 1957. An analysis of arthropods succession in carrion and effect of its decomposition on soil fauna. Australian Journal of Zoology 5 (1): 1-12. [ Links ]
BYRD, J. H.; CASTNER, J. L. 2010. Chapter 2: Insect Forensic Importance. En: Byrd, J. H.; Castner, J. L. (Eds.). Forensic entomology. New YorkWashington, D.C. CRC Press LLC. [ Links ]
CAMACHO, G. P.; USAQUÉN, W. 2004. Ciclo de vida de Lucilia sericata (Diptera: Calliphoridae) como primera especie colonizadora presente en hígado humano realizado en el Instituto Nacional de Medicina Legal y Ciencias Forenses. Revista Instituto de Medicina Legal y Ciencias Forenses 18 (2):31-36. [ Links ]
CAMACHO, G. 2005. Sucesión de la entomofauna cadavérica y ciclo vital de Calliphora vicina (Diptera: Calliphoridae) como primera especie colonizadora, utilizando cerdo blanco (Sus scrofa) en Bogotá. Revista Colombiana de Entomología 31 (2): 189-197. [ Links ]
CATTS, E. P.; GOFF, M. L. 1992. Forensic entomology in criminal investigations. Annual Review of Entomology 37: 253-272. [ Links ]
CONNELL, J. H.; SLAYER, R. O. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist 111: 1119-1144. [ Links ]
FLÓREZ, E.; WOLFF, M. 2009. Descripción y clave de los estadios inmaduros de las principales especies de Calliphoridae (Diptera) de importancia forense en Colombia. Neotropical Entomology 38 (3): 418-429. [ Links ]
FULLER, M. E. 1934. The insect inhabitants of carrion, a study in animal ecology. Bulletin of the Council for Scientific and Industrial Research, Melbourne 82 (1): 5-62. [ Links ]
GOFF, M. L.; ODOM, C. B. 1987. Forensic entomology in the Hawaiian-Islands-3 case-studies. The American Journal of Forensic Medicine and Pathology 8 (1): 45-50. [ Links ]
GRISALES, D.; RUIZ, M.; VILLEGAS, S. 2010. Insects associated with exposed decomposing bodies in the Colombian Andean Coffee Region. Revista Brasileira de Entomología 54 (4): 637-644. [ Links ]
GUIMARAES, J. H.; D'PRADO, A. P.; LINHARES, A. X. 1978. Three newly introduced blowfly in southern Brazil (Diptera: Calliphoridae). Revista Brasileira de Entomología 22: 53-60. [ Links ]
HAMMER, Ø.; HARPER, D. A. T.; RYAN, P. D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontología Electrónica 4 (1): 9. [ Links ]
IBM. 2012. IBM SPSS Statistics for Windows, Version 21.0. Armonk, NY: IBM Corp. [ Links ]
LAMBIASE, S.; CAMERINI, G. 2011. Spread and habitat selection of Chrysomya albiceps (Wiedemann) (Diptera: Calliphoridae) in northern Italy: forensic implications. Journal of Forensic Science 57 (3): 799-801. [ Links ]
MARTINEZ, E.; DUQUE, P.; WOLFF, M. 2007. Successional pattern of carrion-feeding insects in paramo, Colombia. Forensic Science International 166 (2-3): 182-189. [ Links ]
MAVÁREZ-CARDOZO, M. G.; ESPINA DE FEREIRA, A. I.; BARRIOS-FERRER, F. A.; FEREIRA-PAZ, J. L. 2005. La Entomología Forense y el Neotrópico. Cuadernos de Medicina Forense 11 (39): 23-33. [ Links ]
McALEECE, N.; GAGE, J. D. G.; LAMBSHEAD, P. J. D.; PATERSON, G. L. J. 1997. BioDiversity Professional statistics analysis software. Scottish Association for Marine Science y the Natural History Museum London. [ Links ]
MÉGNIN, J. P. 1884. La Faune des Cadavres. Encyclopedie scientifique des Aide-Memorie, G. Masson, Gauthier-Villars et Fils, Paris. 214 p. [ Links ]
NUORTEVA, P. 1977. Sarcosaprophagous insects as forensic indicators. pp. 1072-1095. En: Tedeschi, C. G.; Eckert, W. G.; Tedeschi, L. G (Eds.). Forensic medicine: A study in trauma and environmental hazards Vol. II, Ed Toronto. 1680 p. [ Links ]
ORDOÑEZ, A. N. 2004. Aspectos sucesionales de la entomofauna sarcosaprófaga asociada a cadáveres de conejo en zonas aledañas a Tenjo (Cundinamarca). Trabajo de grado, Carrera de Biología, Pontificia Universidad Javeriana. Bogotá, 172 p. [ Links ]
ORDOÑEZ, A. N.; GARCIA, M. D.; FAGUA, G. 2008. Evaluation of efficiency of Schoenly trap for collecting adult sarcosaprophagous dipterans. Journal of Medical Entomology 45 (3): 522-532. [ Links ]
PINILLA-BELTRÁN, Y. T.; SEGURA, N. A.; BELLO, F. J. 2012. Synanthropy of Calliphoridae and Sarcophagidae (Diptera) in Bogotá, Colombia. Neotropical Entomology 41 (3): 237-242. [ Links ]
PAYNE, J. A. 1965. A summer carrion study of the baby pig Sus scrofa Linnaeus. Ecology 46 (5): 592-602. [ Links ]
PÉREZ, S. P.; DUQUE, P.; WOLFF, M. 2005. Successional behavior and occurrence matrix of carrion-associated arthropods in the urban area of Medellín, Colombia. Journal of Forensic Sciences 50 (2): 1-7. [ Links ]
SCHOENLY, K. G. 1981. Demographic bit trap. Environmental Entomology 10: 615-617. [ Links ]
SCHOENLY, K. G.; REID, W. 1987. Dynamics of heterotrophic succession in carrion arthropod assemblages: discrete series or a continuum of change? Oecologia 73 (2): 192-202. [ Links ]
SCHOENLY, K. G.; REID, W. 1989. Dynamics of heterotrophic succession in carrion revised (a reply to Boulton and Lake). Oecologia 79 (1): 140-142. [ Links ]
SEGURA, N. A.; USAQUÉN, W.; SÁNCHEZ, M.; NARVÁEZ-SÁNCHEZ, R.; CHUAIRE, L.; CAMACHO, G. P.; RAMÍREZ, L. A.; CARREÑO, M. I.; BELLO, F. J. 2005. Curvas de crecimiento y desarrollo de los primeros insectos colonizadores (Diptera: Calliphoridae) sobre cadáveres de cerdo (Sus scrofa) en Bogotá, Colombia. Revista de Investigación 5 (1): 129-140. [ Links ]
SEGURA, N. A.; USAQUÉN, W.; SÁNCHEZ, M. C.; CHUAIRE, L.; BELLO, F. 2009. Succession pattern of cadaverous entomofauna in a semi-rural area of Bogotá, Colombia. Forensic Science International 187 (1): 66-72. [ Links ]
SOUZA, C. R.; VON ZUBEN, C. J. 2012. Diversity and synanthropy of Calliphoridae (Diptera) in the region of Río Claro, SP, Brazil. Neotropical Entomology 41: 243-248. [ Links ]
SMITH, K. G. V. 1986. A manual of forensic entomology. London, Ithaca, British Museum of Natural History, Cornell Univ. Press. Londres, Reino Unido. 205 p. [ Links ]
VÉLEZ, M. C.; WOLFF, M. 2008. Rearing five species of Diptera (Calliphoridae) of forensic importance in Colombia in semicontrolled field conditions. Papéis Avulsos de Zoología 48 (6): 41-47. [ Links ]
WOLFF, M.; URIBE, A.; ORTIZ, A.; DUQUE, P. A. 2001. A preliminary study of forensic entomology in Medellín, Colombia. Forensic Science International 120 (1-2): 53-59. [ Links ]
Recibido: 17-feb-2014
Aceptado 22-oct-2014