










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.2 Bogotá July/Dec. 2014
ARTÍCULOS DE INVESTIGACIÓN / RESEARCH PAPERS
Defensive behavior of africanized honeybees (Hymenoptera: Apidae) in Dourados-Mato Grosso do Sul, Brazil
Comportamiento defensivo de las abejas africanizadas (Hymenoptera: Apidae) en Dourados-MS, Brasil
Márcia Regina FaitaI; Rita Maria Mattoso Colman CarvalhoII; Valter Vieira Alves-juniorI, II; José Chaud-nettoIII
IM. Sc., Ph. D. Programa de Pós-Graduação em Entomologia e Conservação da Biodiversidade, Universidade Federal da Grande Dourados. Rodovia Dourados/Itahum, Km 12, Caixa Postal 241, CEP: 79.804-970, Dourados-MS, Brasil. valteralves@ufgd.edu.br. Corresponding author
IIM. Sc. Faculdade de Ciências Biológicas e Ambientais, da Universidade Federal da Grande Dourados, Rodovia Dourados/Itahum, Km 12, Caixa Postal 241, CEP: 79.804-970, Dourados-MS, Brasil
IIIPh. D. Departamento de Biologia, Instituto de Biociências de Rio Claro, Universidade Estadual Paulista - UNESP, Avenida 24 A 1515, CEP 13506-900, Rio Claro-SP, Brasil
ABSTRACT
African bees were introduced in Brazil in 1956, in an attempt to improve honey production. The accidental hybridization between African and European breeds originated africanized bees, which are very well adapted to the local climate. That bee poly-hybrid has an initial production of honey 70 % more than Europeans. However, African and africanized bees were much more defensive than European subspecies, which required the development of appropriate management techniques. Beekeepers in southern Mato Grosso do Sul learned to work with africanized bees. The aim of this study was to evaluate the defensive behavior of Africanized bees in Dourados MS, using a ball of black leather and artificial enemy. There were is recorded, the time to deliver the first bite, the time it takes to enrage (attack the enemy with great intensity), the distance from the pursuit of the enemy after the first attack and the number of bites left in the beanbag. The results indicate a significant concentration of colonies of bees with different intensity and defensive behavior, but similar to that presented by africanized bees in the 60/70, particularly in the region of Ribeirão Preto (SP). It appears, in terms of their defensive behavior, bees in the southern region of the state, did not suffer influence by bees of European origin.
Key words: Defensive behavior; Apis mellifera; Beekeeping; Africanized Honeybees.
RESUMEN
Las abejas africanas fueron introducidas en Brasil en 1956, en un intento de mejorar la producción de miel. La hibridación accidental entre la raza africana y europea, originó abejas africanizadas que están muy bien adaptadas al entorno local. Esa abeja poli-híbrida presenta una producción inicial de miel 70 % más que la europea. Sin embargo, dado que las abejas africanas y africanizadas eran mucho más defensivas que las subespecies europeas, se requirió el desarrollo de técnicas de manejo apropiadas. Apicultores en el sur de Mato Grosso do Sul aprendieron a trabajar con las abejas africanizadas. El objetivo de este estudio fue evaluar el comportamiento defensivo de las abejas africanizadas en Dourados, utilizando una pelota de cuero negro como enemigo artificial. Para ello, se registraron, el tiempo para la primera picadura, el tiempo que toma para enfurecer (atacar al enemigo con una gran intensidad), la distancia de la persecución del enemigo después del primer ataque y el número de picaduras dejadas en la pelota. Los resultados indican una importante concentración de colonias de abejas con comportamiento defensivo y diversa intensidad, pero similar a la presentada por las abejas africanizadas en las décadas del 60/70, en particular en la región de Ribeirão Preto (SP ). Por lo tanto, en la actualidad las abejas en la región sur del estado, al parecer no fueron influenciadas por las de origen europeo, al menos en lo que respecta al comportamiento defensivo.
Palabras clave: Comportamiento de defensa; Apis mellifera; Apicultura; Abejas africanizadas.
Introduction
Before the introduction of the African honeybees (Apis mellifera scutellata Lepeletier, 1836) to Brazil, the Brazilian honey production depended of two main collector species: the German bees (A. mellifera mellifera Linnaeus, 1758) and the Italian ones (A. mellifera ligustica Spinola, 1806) which were very calm, but presented incompatible productivity in relation to the blooms offered in the region of Piracicaba and Rio Claro (Stort 1971). In an attempt to improve honey production, in 1956 W. E. Kerr brought from South Africa and Tanzania 49 queens of A. m. scutellata which were introduced in test colonies maintained in the forest of Camaquan, at 14 km from Rio Claro - SP (Kerr 1967). According to that scientist, the introduction of a race of bees more adapted to the Brazilian environmental conditions would increase the honey production (Kerr 1967). So, the offspring of those queens would bee presumably better suited for the tropical and subtropical climate conditions found in some Brazilian states, which are similar to those of their native regions. Nevertheless, in March of 1957, twenty-six African bee swarms accidentally escaped from the experimental colonies and their virgin queens were mated by drones produced in resident European honeybee colonies of the region. The first poly-hybrid bees resulting from crossings among A. m. scutellata and the European subspecies A. mellifera ligustica, A. mellifera mellifera, A. mellifera caucasica (Stort and Gonçalves 1994; Pereira and Chaud-Netto 2005) and also A. mellifera iberica (Ruttner 1986) inherited some scutellata-like traits, as reproductive, foraging and defensive behaviors. Because of these characteristics they were called africanized honeybees (Diniz et al. 2003). Researchers all over the world were surprised and even amazed with the incredible relationships between these hybrid honeybees and the neotropical environment because they have high adaptability to variable ecological conditions, produce a great number of swarms over the year, and their workers are efficient collectors of nectar and pollen, competing for these floral resources with many species of native bees (Pereira and Chaud-Netto 2005; Traveset and Richardson 2006; Carbonari et al. 2009; Brizola- Bonacina 2009).
The first tests of productivity performed by Kerr with the africanized honeybees, in the region of Rio Claro - SP, showed an increase of over 70% in the honey production of the newcomers' bees in relation to the bees of European origin (Kerr 1967; Gonçalves et al. 1972). This result is one of the reasons for the preference of most beekeepers for africanized bees, mainly after they learned how to work with those bees and developed appropriate management techniques.
The protection of the colony against intruders and/or invaders is a very important behavior for colony survival (Free 1980). The African honeybee subspecies brought to Brazil, as well as the first africanized hybrids produced in the region of Rio Claro showed a more intense defensive behavior in relation to the German (A. m. mellifera) and Italian subspecies (A. m. ligustica), which had already been established into the country (Stort 1971).
In Brazil, any other information about the introduction of African queen bees was transmitted since 1956. On the other hand, 23.200 honeybee queens of European subspecies were spread in the South and Southeast regions of the country from 1963 to 1972, being most of them Italian and Caucasian (Gonçalves et al. 1972). Nowadays, more than 57 years after the African bee advent, their descendants no longer exhibit such a strong defensive behavior. This is mainly due to the proper management techniques, the continuous introduction of European queen bees in the most vicious populations and the selection of queen bees from gentle africanized colonies, performed consciously by most of Brazilian beekeepers from South and Southeast states. However, the defensive behavior of africanized bees remains highly distinguished. Collins et al. (1994) stated that it is possible to distinguish africanized bees (more aggressive) from European bees (less aggressive), based on their morphological characters associated with behavioral responses by using pheromones for tests of defensiveness.
In sul of State of Mato Grosso do Sul (Dourados/MS)-Brazil, beekeepers always worked with africanized honeybees and since those insects arrived to the region they have no information about the introduction of any other bee subspecies. Thus, more than 57 years after the arrival of the African honeybees, it is more probable to find bees that did not undergo any influence of European races in this region, at least regarding to their defensive behavior. This can be inferred only for the states of South and Southeast. According to reports of most beekeepers of those regions, they have observed and experienced varying degrees of response for this behavior in daily activities with bees, which indicates a large phenotypic diversity.
Thus, this study aimed to develop a current appreciation for the defensive behavior of africanized honeybees in the region of Dourados (MS) in Brazil.
Material and methods
The bees used in this study came from Carbonari Beekeeping located near the city of Dourados-MS, Coqueiro Farm, in the "Mata do Azulão", MS Highway 162, Km 22 - (22°12'S 54°54'W, 430 masl - GPS).
The tests were performed according to Stort (1971; 1974) in ten homogeneous hives, containing the same number of combs covered with bees, and approximately the same quantity of brood and food (honey plus pollen). The colonies were developed from swarms captured in the nature, which maintained their original queens. All tests were performed in the afternoon, following a period of three days or more, (when necessary). Two colonies were assessed daily, separated minimum 25m each other and by an area of partially dense vegetation. For each colony the "defensiveness" test comprised five replications: in each bioassay an artificial enemy, a black leather ball 2 cm in diameter filled with cotton was used to provoke the bees (the black color irritates the bees). The target was shaken in front of each hive for 60 seconds after the first sting, and the following behavioral variables were evaluated:
Time (in seconds) to occur the first sting in a black leather ball.
Time (in seconds) taken for the bees to become furious after the first attack to the leather ball.
Observer chasing distance (in meters): distance which the bees followed the observer after the ending of each 60-seconds test, while he was walking away.
Number of stings left in the black leather ball during the test.
The behavioral variables recorded for each bioassay were compared in pairs by Spearman´s Correlation Test (rs) with significance level of 0.5%, with the statistical program PAST version 1.91 was used (Hammer et al. 2009).
Results and discussion
The results of bioassays regarding the defensive behavior variables of africanized honeybee workers (A. mellifera L.) in the region sul of state are displayed in Table 1.
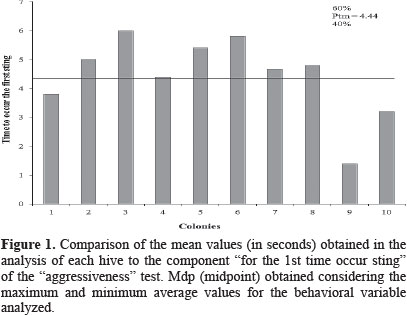
The time to occur the first sting in the black leather ball ranged from 1.4 ± 0.54 s. (hive 9) to 6.0 ± 3.60 s. (hive 3) (Table 1). As can be seen in figure 1, taking into account the mean value response time to the physical stimulus used (Ptm = 4. 44 s.), in 40% of the cases (colonies 1, 4, 9 and 10) the bees' attack was faster, evidencing an intense defensiveness. For the same variable Stort (1971; 1975a) recorded a mean time of 3.15 seconds for African honeybee colonies and 12.86 seconds for africanized ones. The results of this research (Table 1) are very similar to those recorded by that author for African bees. So, the africanized honeybees from the region of Dourados/MS may be considered as defensive as the African bees used by Stort. The same author considered this trait as a product of a four-gene interaction without dominance among them.
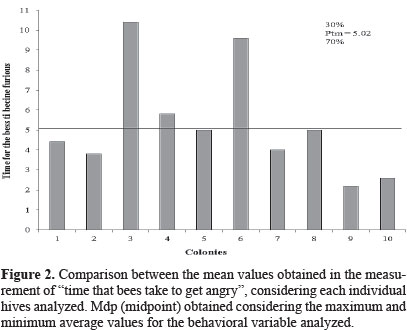
Using the technique developed by Stort (1971; 1974) to assess the defensive behavior of africanized honeybees during the rainy season in Mossoró - RN, Nascimento et al. (2008) observed bee attacks against an artificial enemy, in three periods of the day. Those authors reported a lower number of stings on the enemy between 7 and 9 am and observed that bee responses were faster between 15 and 17 hours They found no significant correlation between relative humidity and bee attacks, contradicting the results obtained by other authors (Brandeburgo et al. 1982; Brandeburgo 1990; Brandeburgo and Gonçalves 1990), who pointed out that defensive behavior of africanized honeybees is influenced by climatic conditions, mainly by relative humidity and temperature. In A. mellifera cells, which cover the venom reservoir, produce isopentyl acetate, an important alarm pheromone that is used to "tag" the enemy (Free 1980). This substance and other venom components are very volatile, being disseminated by the air soon after the enemy is stung, so that other honeybee workers are alerted to defend the colony by attacking vigorously the marked target. Thus, consequently, it is assessed the time taken for the bees to become furious and attack the artificial enemy in large scale. The maximum and minimum mean values recorded for this behavioral trait were 10.33 ± 4.50 s. (colony 3) and 2.2 ± 0.44 s. (colony 9), respectively (Table 1), and 70% of the colonies analyzed presented high defensiveness, taking into account the mean point obtained (Ptm = 5.02 s.; Fig. 2). Results of Stort (1971; 1976) for the same variable averaged 9.04 seconds for African honeybees and 23.46 seconds for africanized ones. Stort suggested that this behavior would be controlled by two pairs of genes that would be recessive in the africanized honeybees whenever are crossed with Italian bees (Stort 1971, 1976; Stort and Gonçalves 1991).
When an animal is stung by bees the cells which cover the venom reservoir of the stings left in its body liberate isopentyl acetate, and the animal will be attacked by other worker bees attracted by that pheromone. On the other hand, those bees, which already lost their stings, continue to attack the enemy using their mandibles. By this action, another volatile pheromone, produced in the mandibular glands (2 - heptanone), is released. It is also efficiently used for marking the enemy and rapidly attracts a great number of new bees involved in the colony defense. Consequently, the victim will exhale a strong alarm pheromone and when it tries to escape will be pursued by dozens of bees. Based on these observations, it was measured the distance that these bees chased the observer after the artificial enemy was exposed to the attack unleashed by the bees.
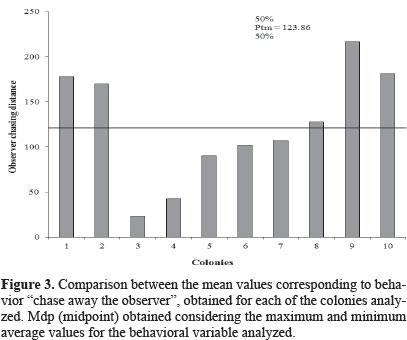
Results of this research revealed that the average observer chasing distance (Table 1) ranged from 23.33 ± 5.85 m (colony 3) to 216.6 ± 7.95 m (colony 9). In 50 % of the observations (Fig. 3) the workers showed higher defensive behavior (Ptm = 123. 86 m). The values recorded by Stort (1971; 1980) for this variable averaged 160.20 m for African honeybee colonies and 38.80 m for africanized ones. The results in Table 1 indicate that africanized bees of southern part of the state are very similar to the African bees evaluated by Stort (1971) which display a high defensive behavior. According to this author, this trait would be determined by the interaction of three pairs of genes with complete dominance. The great variation observed by Stort was due to this interaction.
Regarding to the potential defensiveness of these africanized honeybees, in the next phase of the experiment we recorded the number of stings left in the black leather ball after it was attacked by the worker bees. The averaged number of stings left in the artificial enemy varied from 13.2 ± 0.83 (colony 2) to 28.4 ± 2.50 (colony 6) (Table 1, Fig. 4).
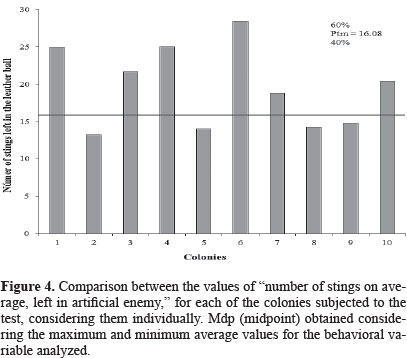
Taking into account the mean value of stings left in the leather ball (Ptm = 16.08), we can observe that 60% of all the colonies showed high defensiveness. Stort (1975b) recorded the following averages for the same behavioral component: 61.15 stings for African bees and 48.13 stings for africanized ones. These values are quite higher in relation to those recorded for the colonies of sul of State, and this result indicates that these bees presented a less intense reaction in relation to the artificial enemy used in the behavioral bioassays performed. This peculiar response against the target probably is due to particular genotypic interactions found in the Africanized honeybees of this region.
Using an elegant and efficient selection program, which would become a classical referenced work in behavior genetics, Rothenbuhler (1964a) obtained two inbred strains of European honeybees which differed in relation to nest cleaning behavior. One of them, termed Brown line, was resistant to Bacillus larvae White, a sporulating bacterium that causes American foulbrood, a terrible disease, which cannot be controlled by chemical products. The second honeybee line, named Van Scoy, was susceptible to the same bacterium. Resistant worker bees are very efficient to remove dead larvae and pupae infected by B. larvae from the brood nest and were called "hygienic honeybees". Conversely, susceptible worker bees remove dead brood very slowly or not at all and consequently were termed "non hygienic honeybees" (Rothenbuhler 1964b). In conjunction with the nest cleaning behavior, the same author also discovered that Van Scoy line had gentle honeybee workers, which almost never sting the observer during colony inspections. On the other hand, the resistant Brown worker bees frequently sting the experimenter (Rothenbuhler 1964a). Regarding to the number of stings left in the observer, Rothenbühler (1964b) analyzed 29 backcrosses of F1 to Brown line queens and stated that the stinging behavior would be controlled by two or more genetic loci and that the tendency to sting was recessive. Stort (1975b) reached the same conclusion and pointed out that the defensive behavior component is due to the activity of two pairs of genes without dominance between them, which "add" their expressiveness and consequently give rise to different phenotypes. According to Free (1980), hives with different genetic pool can vary widely in behavioral expressiveness.
Considering the results obtained in this research (Table 1), except for the number of stings left in the black leather ball, colonies 9 and 10 were the most defensive, while the honeybee workers from colony 3 were less defensive when compared with the other. Colony number 6 showed a less intense defensiveness in relation to the time to occur the first sting in the leather ball and time taken for the bees to become furious. Nevertheless, the bees from that same colony showed great capacity to pursue the observer, and left the largest number of stings in the leather ball. The results of the Spearman's correlation test revealed significant correlation between the following traits (Table 2): association between components of defensive behavior compared: Time to occur the first sting and Time to become furious (rs = 0.7642; P= 0.017), Time to become furious and Observer chasing distance (rs = -0.7637; P= 0.001), Time to occur the first sting and Observer chasing distance (rs = -0.7480; P= 0.017). The dependency relationships among these three traits were already expected because the shorter the time to occur the first sting in the leather ball, the shorter the time for the bees to become furious and hence the greater the observer chasing distance. A quick answer to the stimulus represented by the artificial enemy results in a more intense defensive behavior and consequently a faster time for the bees to become angry. So, a higher amount of alarm pheromone is released on the victim and, finally, a greater observer persecution distance is recorded. These relationships explain the negative correlations results obtained when the data related to the "observer chasing distance" were compared with those concerning to the variables: "time to occur the first sting" and "time to become furious". The other comparisons among traits of the defensive behavior showed non-significant results (Table 2).
Therefore, the results of the present research showed that africanized honeybees from the region sul of State of Mato Grosso do Sul, have an effective defensive behavior, with intensity similar to that observed in the decades of 60/70, when many accidents with animals and humans took place. In comparison with the results recorded by Stort (1971; 1975b) for africanized honeybees of Ribeirão Preto, the main difference observed in this study refers to the number of stings left in the leather ball, which showed a lower mean value in this study (Table 1). The results recorded for the other components of defensive behavior were similar to those previously obtained by Stort.
Thus, we can conclude that the africanized honeybees of Dourados-MS over the years have maintained a tendency to keep the defensive behavior originally presented by the first in Africanized bees that arrived to the region. In recent years, the decrease of recorded accidents was due to the development of beekeeping techniques most suitable for handling the colonies, the ability of the local beekeepers and their increasing knowledge on bee biology.
The africanized bee colonies produce swarms at least twice a year and each cluster contains a new queen who will transmit to their descendants at least a part of the genes responsible for the defensive behavior of the colony. Furthermore, these queens produce drones carrying genes for defensiveness. Thus, the behavioral characteristics observed today would be more widespread in the regional population. If these naturally produced queens are maintained in the population, the genes responsible for this behavior will also be kept.
Based on the results of this study we can also conclude that the defensive behavior of africanized bees in the region sul of state is diversified in intensity. Furthermore, the results also indicate a significant concentration of colonies containing bees with defensive behavior of intensity similar to that presented by africanized honeybees analyzed between 1960 and 1970. Thus, considering that the bees in sul of state maintained defensive characteristics very similar to those recorded in that period, particularly in the region of Ribeirão Preto, one can say that the actual bees from sul of State of Mato Grosso do Sul apparently were not influenced by bees of European origin, at least in regards the defensive behavior.
Literature cited
BRANDEBURGO, M. A. M. 1990. Aggressive behavior of bees. Ciencia y Cultura 42: 1025-1034. [ Links ]
BRANDEBURGO, M. A. M.; GONÇALVES, L. S. 1990. Environmental influence on the aggressive (defence) behaviour and colony development of Africanized bees (Apis mellifera). Ciencia y Cultura 42: 759-771. [ Links ]
BRANDEBURGO, M. A. M.; GONÇALVES, L. S.; KERR, W. E. 1982. Effects of Brazilian climatic conditions upon the aggressiveness of Africanized colonies of honeybees. In: Jaisson, P. (Org.) Social insects in the Tropics, vol. 1, 1st ed. Paris - France: Université Paris - Nord, p. 255-280. [ Links ]
BRIZOLA-BONACINA, A. K. 2009. Presença de Apis mellifera L. em uma região de cerrado em Dourados (MS) e sua relação com a fauna de abelhas nativas. Tese (Doutorado) - Universidade Estadual Paulista, Instituto de Biociências de Rio Claro, SP. [ Links ]
CARBONARI, V.; POLATTO, L. P.; ALVES-JUNIOR, V. V. 2009. Evaluation of the impact on Pyrostegia venusta (Bignoniaceae) flowers due to nectar robbery by Apis mellifera (Hymenoptera, Apidae). Sociobiology 54: 373-382. [ Links ]
COLLINS, M. A.; DALY, H. V.; RINDERER, T. E.; HARBO, J. R.; HOELMER, K. 1994. Correlations between morphology and colony defense in Apis mellifera L. Journal of Apicultural Research 33: 3-10. [ Links ]
DINIZ, N. M.; SOARES, A. E. E.; SHEPPARD, W. S.; DEL LAMA, M. A. 2003. Estructura genética de las poblaciones de abejas procedentes de Brasil y Uruguay. Genetics and Molecular Biology 26: 47-52. [ Links ]
FREE, J. B. 1980. A organização social das abelhas (Apis). Coleção Temas de Biologia, vol. 13, E.P.U. - EDUSP, São Paulo, 79 p. [ Links ]
GONÇALVES, L. S.; KERR, W. E.; CHAUD-NETTO, J.; STORT, A. C. 1972. Some comments on the "Final Report of the Committee on the African Honeybee". National Research Council - N.A.S. Mimeographic paper. USA: Cornell University, 35 p. [ Links ]
HAMMER, O.; HARPER, D. A. T.; RYAN, P. D. 2009. PAST version 1.91: Paleontological Statistical Software package for education and data analysis. Paleontologia Electronica 4: 9 p. [ Links ]
KERR, W. E. 1967. A solução é criar uma raça nova. Guia Rural, Edição Coopercotia 2: 20-22. [ Links ]
NASCIMENTO, F. J.; MARACAJÁ, P. B.; DINIZ-FILHO, E. T.; MAIA, F. J.; NASCIMENTO, R. M. 2008. Agressividade de abelhas africanizadas (Apis mellifera) associada à hora do dia e à umidade relativa do ar em Mossoró - RN. Acta Veterinaria Brasilica 2: 80-84. [ Links ]
PEREIRA, A. M.; CHAUD-NETTO, J. 2005. Africanized honeybees: Biological characteristics, urban Nesting behavior and accidents caused in Brazilian cities (Hymenoptera: Apidae). Sociobiology 46: 535- 550. [ Links ]
ROTHENBUHLER, W. C. 1964a. Behaviour genetics of nest cleaning in honey bees. IV. Responses of F1 and backcross generations to disease-killed brood. American Zoologist (a) 4: 111-123. [ Links ]
ROTHENBUHLER, W. C. 1964b. Behaviour genetics of nest cleaning in honey bees. I. Response of four inbred lines to disease-killed brood (b). Animal Behaviour 12: 578-583. [ Links ]
RUTTNER, F. 1986. Geographical variability and classification. In: Rinderer, T. E. (Ed.). Bee genetics and breeding. Orlando: Academic Press, 23-56. [ Links ]
STORT, A. C. 1971. Estudo genético da agressividade de Apis mellifera. Araraquara: Faculdade de Filosofia Ciências e Letras de Araraquara. 166 p. (Ph. D. Thesis). [ Links ]
STORT, A. C. 1974. Genetic study of the aggressiveness in two subspecies of Apis mellifera in Brazil. I. Some tests to measure aggressiveness. Journal of Apicultural Research 13: 33-38. [ Links ]
STORT, A. C. 1975a. Genetic study of the aggressiveness of two subspecies of Apis mellifera in Brazil. II. Time at which the first sting reached the leather ball (a). Journal of Apicultural Research 14: 171-175. [ Links ]
STORT, A. C. 1975b. Genetic study of the aggressiveness of two subspecies of Apis mellifera in Brazil. (b) V. Number of stings in the leather ball. Journal of the Kansas Entomological Society 48: 381-387. [ Links ]
STORT, A. C. 1976. Genetic study of the aggressiveness of two subspecies of Apis mellifera in Brazil. III. Time taken for the colony to become aggressive. Ciência e Cultura 28: 1182-1185. [ Links ]
STORT, A. C. 1980. Genetic study of the aggressiveness of two subspecies of Apis mellifera in Brazil. VI. Observer persecution behavior. Brazilian Journal of Genetics 3: 285-294. [ Links ]
STORT, A. C.; GONÇALVES, L. S. 1991. Genetics of defensive behavior. In: SPIVAK, M.; FLETCHER D. J. C.; BREED, M. D. (Org.). The African Honeybee, 1st ed. Boulder - Colorado, USA. Westview Press, p. 329-356. [ Links ]
STORT, A. C.; GONÇALVES, L. S. 1994. A africanização das abelhas Apis mellifera nas Américas. pp. 33-47. In: Barraviera, B. (Ed.). Venenos Animais: Uma visão integrada. Rio de Janeiro, Editora de Publicações Científicas Ltda. [ Links ],
TRAVESET, A.; RICHARDSON, D. M. 2006. Biological invasions as disruptors of plant reproductive mutualisms. Trends in Ecology and Evolution 21: 208-216. [ Links ]
Received: 14-Feb-2014
Accepted: 26-Oct-2014
