Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.2 Bogotá July/Dec. 2014
ARTÍCULOS DE INVESTIGACIÓN / RESEARCH PAPERS
La estructura importa: abejas visitantes del café y estructura agroecológica principal (EAP) en cafetales
The structure matters: bees visitors of coffee flowers and agroecological main structure (MAS)
Juliana Cepeda-valenciaI; Diana Gómez PII; Clara NichollsIII
IBióloga, M. Sc. Candidata al Doctorado en Agroecología. Instituto de Estudios Ambientales (IDEA)-Universidad Nacional de Colombia. Investigadora Laboratorio de artrópodos del Centro Internacional de Física (CIF), Bogotá, Colombia. juliana.cepedav@gmail.com / jcepedav@unal.edu.co. Autor para correspondencia
IIEstudiante de Agronomía Universidad Nacional de Colombia y del Laboratorio de artrópodos del Centro Internacional de Física (CIF), Bogotá, Colombia. dagomezp@unal.edu.co
IIIAgrónoma, Ph. D. Entomology and Biological Control. Profesora Universidad de Antioquia (Colombia) y Profesora ocasional Berkeley University (USA). nicholls@berkeley.edu
RESUMEN
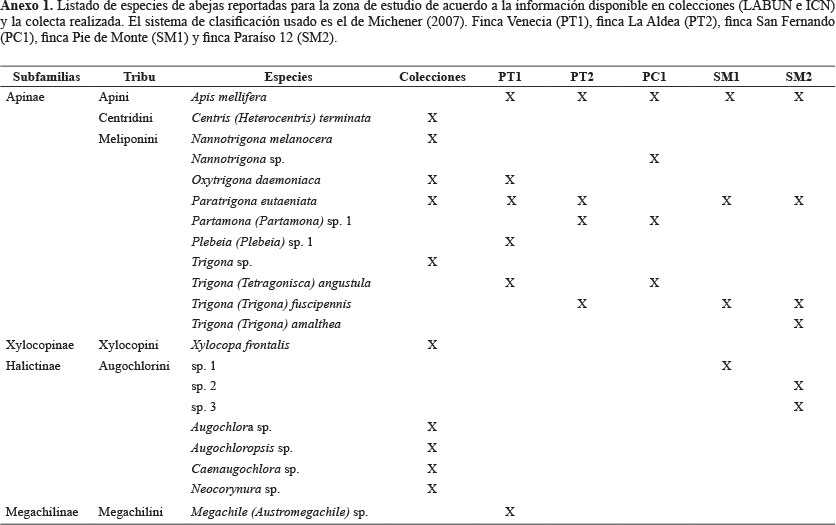
En cafetales, la polinización cruzada influye positivamente en la calidad de los frutos y su eficiencia depende de la riqueza de polinizadores. La Estructura Agroecológica Principal (EAP) es un nuevo concepto para caracterizar agroecosistemas, incluyendo variables locales relacionadas con el manejo agrícola, variables de paisaje y culturales. Para estudiar la relación entre EAP y polinización, se caracterizaron nueve sistemas (cinco cafetales, cuatro bosques) en Cundinamarca, evaluando diez variables locales, la distancia al bosque como variable de paisaje y las abejas visitantes del café. Las colecciones se hicieron durante las floraciones de 2011 y 2012 usando captura manual. Luego de su captura y montaje las abejas se identificaron y se extrajo y fijó el polen de contacto. Se encontraron 130 individuos de tres subfamilias: Halictinae (tres especies), Megachilinae (una especie) y Apinae (nueve especies). Trigona fue el género más rico. Apis mellifera y Paratrigona eutaeniata fueron los visitantes florales más abundantes. La riqueza de abejas presentó dependencia positiva con la riqueza vegetal (P = 0,031; R2 = 0,96) y la cercanía al bosque (P = 0,0075; R2 = 0,52) y negativa con la densidad de café (P = 0,045; R2 = 0,67). En el polen de contacto se identificaron árboles, hierbas y epífitas, concluyendo que las abejas visitan diferentes estratos vegetales en su búsqueda de recursos. También, se identificaron plantas propias del arreglo y otras propias de bosques y cercas vivas. Se concluyó que los cambios en la EAP afectan la composición y riqueza de las abejas y que mantener esta diversidad, implica conservar la diversidad del arreglo y a los ecosistemas cercanos.
Palabras clave: Agroecología; Apoidea; Biodiversidad; Manejo agrícola; Visitantes florales.
ABSTRACT
Cross pollination in coffee increases their quality and the efficiency of pollination has a positive trend with richness of pollinators. The Main Agroecological Structure (MAS) is a new concept to identify and characterize the agroecosystems. The method uses local variables related with agricultural management, landscape and cultural variables. As a first approach to study the relationship between MAS and pollination, we characterized nine systems (five coffee farms, four forests) located in Cundinamarca, using ten local variables, the distance to the forest as a landscape variable and bees (visitors of coffee flowers). The field work was done during the flowerings of 2011 y 2012. After the capture, bees were identified and the pollen loads were analyzed. We found 130 bees of the subfamilies: Halictinae (three species), Megachilidae (one species) and Apinae (nine species). Trigona was the richest genera. The most common visitors of coffee flowers were Apis mellifera and Paratrigona eutaeniata. The bees richness was positive related with plants richness (P = 0.031, R2 = 0.96) and the forest proximity (P = 0.0075, R2 = 0.52), Also, we found a negatively trend with the increase in coffee density (P = 0.045, R2 = 0.67). In the analysis of the pollen loads we identified trees, herbs and epiphytes species, this suggested that bees foraging in all vegetation strata. In this analysis we also found species related with the coffee system and others more related with forest and hedges. We concluded that a change in MAS affecting bee's composition and richness, and preserving richness of bees, implies protection of plant diversity inside the coffee farm and also the maintenance of nearby ecosystems.
Key words: Agroecology; Apoidea; Biodiversity; Agricultural management; Flower visitor.
Introducción
La polinización es un proceso clave tanto para los ecosistemas como para los agroecosistemas, cuya eficiencia es directamente afectada por la riqueza de polinizadores. Además, la deficiencia en la polinización produce efectos cascada que derivan en la reducción de frutos y semillas e incluso en la disminución de la densidad de las plantas en los ecosistemas (Anderson et al. 2011). Este proceso es también crítico para la producción de alimentos y el bienestar humano, relacionando directamente a los ecosistemas con los agroecosistemas (FAO 2008).
Está ampliamente documentada la existencia de una crisis mundial de polinizadores, manifestada en el descenso en la polinización de cultivos por abejas de la miel en América del Norte. En Europa el descenso se observa en los cultivos polinizados por abejorros y mariposas (Nicholls y Altieri 2012). En otros lugares, las comunidades de polinizadores nativos muestran respuestas diversas a los cambios ambientales pero, globalmente, se registra el declive de sus poblaciones (Ghazoul 2005; FAO 2008). Algunos factores que amenazan a los polinizadores incluyen la fragmentación y pérdida de ecosistemas y hábitats naturales por cambios en el uso del suelo, el cambio climático y la introducción de especies exóticas (FAO 2008). También, las prácticas relacionadas con la agricultura industrial pueden comprometer la eficiencia de la polinización, principalmente, el predominio del monocultivo que implica la homogenización de la diversidad vegetal y la concentración temporal de los recursos florales y, por otra, el uso de agroquímicos (Kremen et al. 2002; Ricketts 2004; Nicholls y Altieri 2012).
En una revisión global sobre el impacto de la polinización mediada por animales en cultivos, Klein et al. (2007) encontraron que en el 85% de los cultivos revisados la polinización influía positivamente en la producción. Incluso en cultivos que no dependen de este tipo de polinización, la calidad de la cosecha se incrementa y la producción agrícola se estabiliza en el tiempo (Ricketts et al. 2008). Además, se ha encontrado que la diversidad de insectos polinizadores en los cultivos depende de dos variables: la diversidad floral que implica la diversificación espacio-temporal de recursos (Nicholls y Altieri 2012) y los espacios naturales cercanos al cultivo (Klein et al. 2003a; Kremen et al. 2004; Ricketts 2004), que además de servir como posibles lugares de forrajeo son también lugares de anidación.
Entre los polinizadores, las abejas son consideradas de gran importancia ya que intervienen en la reproducción de más del 50% de las plantas (Nates 2005) y además son importantes polinizadores de las plantas cultivadas. En el estudio de Ricketts et al. (2008) de los 22 cultivos citados las abejas se consideraron polinizadores claves de 20, entre ellos del café tanto de Coffea arabica L. como de Coffea canephora Pierre. También se ha encontrado que la riqueza de polinizadores mejora la eficiencia de la polinización. Por ejemplo, Klein et al. (2003a; 2003b) mostraron que la riqueza de abejas en cafetales de Indonesia estaba positivamente correlacionada con el porcentaje de frutos cuajados, es decir, disminuía el número de abortos por grupo de flores.
En particular, en el sector cafetero de Colombia, se ha dado poca importancia a los estudios relacionados con la polinización, ya que en las variedades de C. arabica que se emplean en el país la capacidad de autopolinización es alta y ésta no se ha considerado un factor limitante para la producción (Arcila 2007), a pesar que tanto para esta especie como para C. canephora, se ha encontrado que la riqueza de abejas visitantes florales, incrementa el porcentaje de frutos cuajados y el peso de los mismos (Klein et al. 2003a; 2003b). Tampoco se ha profundizado en el potencial de los cafetales colombianos como espacios para la conservación de esta fauna, pese a que mundialmente se ha registrado el valor biológico de estos agroecosistemas (Perfecto et al. 1996; Moguel y Toledo 1999; Perfecto et al. 2009), especialmente aquellos con sombrío diversificado, ya que prestan servicios ecosistémicos importantes.
Recientemente, Jaramillo (2012) al examinar la influencia de la polinización abierta y en particular de las abejas como agentes polinizadores en cafetales de Antioquia, encontró que la polinización cruzada ayuda a disminuir el número de abortos y que, además, influye positivamente en el tamaño de los frutos (mayor diámetro y mayor peso seco). También, se encontró en ellos mayor concentración de azúcares (grados brix), lo que puede mejorar la calidad de taza o sabor. Por tanto, Jaramillo concluyó que la polinización cruzada cumple un papel relevante en la producción y calidad de los frutos de café, pese al alto grado de autopolinización.
Por otra parte, la estructura agroecológica principal (EAP) es un concepto reciente propuesto por León 2014 para evaluar los cambios en la diversidad planeada dentro de los agroecosistemas, midiendo tanto su configuración o arreglo espacial, como el grado de conectividad interno entre sus distintos sectores, parches, corredores de vegetación y sistemas productivos, como externo con los hábitats naturales adyacentes. En teoría, el mantenimiento de esta estructura permitiría el movimiento y el intercambio de distintas especies animales y vegetales y potenciaría las funciones ecológicas y, por tanto, la oferta de servicios ecosistémicos. Sin embargo, los nexos entre el mantenimiento de la EAP y la funcionalidad ecológica no son simples, dada la complejidad de las relaciones ecológicas incluso en agroecosistemas (Vandermeer et al. 2010) y que no todos los grupos biológicos responden de la misma forma a los cambios en la estructura de la vegetación (Perfecto et al. 2003) o al manejo agrícola (Cepeda et al. 2009).
Los cafetales han servido como modelo de estudio para la intensificación agrícola, tanto en relación a los cambios en la diversidad biológica como en la preservación de funciones ecológicas (Moguel y Toledo 1999; Perfecto et al. 2003; Klein et al. 2003a; 2003b; Cepeda et al. 2009; Perfecto et al. 2009; Vandermeer et al. 2010). Aquí se retoman estos agroecosistemas para responder las siguientes preguntas:
(i) ¿Pueden los cambios en la EAP afectar la diversidad de abejas visitantes del café?
(ii) ¿Qué componentes particulares de la EAP en los sistemas cafeteros evaluados se relacionan con la riqueza de abejas?
(iii) ¿Qué otras plantas dentro de los agroecosistemas cafeteros pueden servir a las abejas como recurso?
Materiales y métodos
Área de estudio. El estudio se llevó a cabo durante 2011 y 2012, en nueve localidades de Quipile y Pulí en el departamento de Cundinamarca (Tabla 1). La precipitación media en la región es de 2.800 mm anuales, con dos estaciones de lluvia, abril-mayo y octubre-noviembre (Mesa et al. 1997). Esta estacionalidad afecta las floraciones del café debido a la disponibilidad de agua y energía, lo que hace que las floraciones plenas del café se produzcan generalmente antes de la estación lluviosa.
Caracterización de los lugares de muestreo. Cada localidad seleccionada fue clasificada en un tipo de sistema que se identificó (Tabla 1) como bosque (B), policultivos tradicionales (PT), policultivos comerciales (PC) y sistemas de sombrío mono-específico (SM) de acuerdo a la propuesta de Moguel y Toledo (1999). Inicialmente se hizo una cuantificación de la EAP siguiendo la metodología de León 2014 pero esta no mostró grandes diferencias entre los agroecosistemas evaluados, que presentaron índices de EAP entre 65 - 88, es decir estructuras de moderada a altamente desarrolladas que están bien conectadas con la estructura ecológica del paisaje y en su interior poseen conectores altamente diversificados, manejan arvenses, con policutivos como uso predominante del suelo y que además están soportadas por agricultores de alta conciencia del valor de la biodiversidad y de los efectos ambientales de los sistemas productivos. Este resultado y la imposibilidad de medir la variabilidad en los bosques estudiados con esta metodología, llevó a un acercamiento diferente para cuantificar la EAP como se indica a continuación.
Para la caracterización de la EAP, se ubicaron tres parcelas en cada uno de los nueve sistemas que fueron georreferenciadas. Las parcelas fueron de 5 m de radio en los cafetales (sistemas más homogéneos) y de 12 m de radio en los bosques (sistemas altamente heterogéneos). En cada parcela se midieron las siguientes variables asociadas a la estructura de la vegetación: 1) riqueza de árboles (total por parcela), 2) densidad de árboles, número de árboles en cada parcela / área de la parcela, 3) altura promedio de árboles, 4) diámetro a la altura del pecho de los árboles (DAP promedio), 5) densidad del café, número de cafetos en cada parcela / área de la parcela, 6) DAP (promedio) de los cafetos, 7) altura promedio de los cafetos, 8) riqueza de arvenses, 9) porcentaje de cobertura del dosel usando un densiómetro esférico y 10) la riqueza de plantas como el resultado de la suma entre la riqueza de árboles y arvenses.
Para la identificación de las especies tanto de sombrío como arvenses, se tomaron muestras botánicas en los sistemas estudiados. La identificación se hizo con ayuda de la guía de Gómez y Rivera (1995), la colección en línea del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN, 2004) y el diccionario de nombres comunes de plantas de plantas de Colombia (Bernal et al. 2011).
Además, en entrevistas semiestructuradas con los caficultores se evaluó el uso de agroquímicos, su frecuencia de aplicación y otras variables culturales de los sistemas. La evaluación sobre el uso de agroquímicos en cada finca se centró en dos indicadores, el número de agroquímicos usados y la frecuencia anual de aplicación de estos productos. El uso de agroquímicos se evaluó de 1 a 9, siendo 1 un manejo completamente orgánico, 3 el uso de insumos químicos para fertilización (bajo), 6 el uso de fertilizantes junto con otro agroquímico para control de hongos o insectos (moderado) y 9 un manejo completamente convencional que implica el uso de agroquímicos para fertilización, control de insectos, hongos y arvenses (alto). En los sistemas categorizados como bosques (B1, B2, B3 y B4) se consideró que estas variables tomarían el valor de 0. Así, se midieron un total de 12 variables locales en cada sistema para su caracterización y posterior correlación con la riqueza de abejas evaluada (Tabla 2). Como variable de paisaje se midió la distancia al bosque más cercano con ayuda del GPS - Garmin. Un resumen de las variables medidas se presenta en la tabla 2.
Colección de abejas, extracción e identificación de polen de contacto. Aunque el café presenta flores todo el año, hay dos periodos de floración plena, marzo-abril y septiembre-octubre (Arcila 2007) las cuales son cortas y se restringen a aproximadamente una semana, tiempo durante el cual se realizaron las observaciones. Cabe resaltar que el 2011 fue particularmente lluvioso, durante el año se presentó el fenómeno de La Niña así que es posible anticipar su influencia sobre los datos.
Las abejas se colectaron durante las floraciones de julio 2011 y marzo 2012 en los cinco cafetales mediante captura manual, entre 8-11 a.m. en su horario de mayor actividad (Klein et al. 2003a, 2003b; Ricketts 2004).
Para cada cafetal se hicieron dos réplicas de dos horas cada una, para un total de cuatro horas de muestreo efectivo. Cada muestra consistió en una persona recorriendo aleatoriamente el cafetal hasta encontrar un cafeto con flores atractivas para las abejas. En él, observó la actividad de las abejas y las colectó en frascos individuales. Esta actividad fue repetida hasta concluir las dos horas de muestreo. La unidad de muestreo fue temporal y no responde a un área específica o número de arbustos pero, en general, se trató de recorrer todo el cafetal durante el muestreo. Esta decisión se tomó porque la floración del café es corta (una semana), así mismo, las flores de café permanecen abiertas tres días y solo durante el primer día son atractivas para las abejas (Jaramillo 2012). Por tanto, con esta estrategia se buscó maximizar las probabilidades de encontrar actividad de las abejas al interior de los cafetales.
Las abejas fueron montadas en seco e identificadas con ayuda de claves especializadas y de especialistas. Para establecer qué otras plantas dentro del cafetal fueron visitadas como fuentes potenciales de alimento, luego de su captura y montaje, se extrajo y fijó el polen adherido a su cuerpo, diferenciando entre aquel del abdomen y las corbículas (polen de carga) y el polen de contacto ubicado principalmente en la cabeza y dorso. Este último se tiñó y montó en placas con la técnica de gelatina de fuscina (Dafni 1992) para su conteo en microscopio e identificación botánica con ayuda de las láminas de la palinoteca del Laboratorio de Abejas de la Universidad Nacional de Colombia (LABUN) y de las claves de Roubik y Moreno (1991) y Herrera y Giraldo (1996).
Análisis estadístico. En general, los análisis se hicieron con el programa estadístico R, usando la plataforma InfoStat 2012 (Di Rienzo et al. 2012) como interfase.
Caracterización de los lugares de muestreo. Dado el gran número de variables locales (12) utilizadas para la caracterización de los sistemas, fue necesario escoger entre aquellas que podrían estar correlacionadas entre sí, para reducir otros análisis estadísticos. Así, primero se eliminaron las variables de "riqueza de árboles" y "riqueza de arvenses" que se encontrarían naturalmente correlacionadas con la "riqueza de plantas", que es la suma de ambas. Con las diez variables restantes se corrió un análisis de componentes principales (ACP) para identificar si las variables usadas eran buenos descriptores de la intensificación agrícola. También se buscó reconocer cuáles variables estarían afectando más la varianza. Para reducir la dimensionalidad del análisis, se usaron los valores de "eigen" del primer componente del ACP (Di Rienzo et al. 2012).
Con las variables seleccionadas en el ACP se corrió un análisis de varianza no paramétrica de Kruskall- Wallis donde las categorías (B, PT, PC y SM) se consideraron tratamientos. Además, de este análisis se efectuaron pruebas de comparación entre pares de tratamientos usando las diferencias medias de rango para las variables evaluadas.
También, se agruparon los lugares de muestreo de acuerdo a las variables locales con ayuda de un análisis de conglomerados usando Bray-Curtis como índice de distancia y el enlace-promedio como método de agrupamiento. Para el conglomerado se descartaron las variables relacionadas con el café, ya que estas tienen valor de 0 en los bosques y podrían afectar la capacidad discriminativa del análisis.
Diversidad de abejas visitantes del café. Se comparó la diversidad alfa de abejas visitantes del café entre dos tratamientos, policultivos (P) y cafetal de sombrío mono específico (SM), utilizando la función "compare diversity" del programa PAST 2.14 (Hammer et al. 2001) y la diversidad beta calculando el índice de Jaccard con ayuda del mismo programa. Además, se realizó una regresión lineal de la riqueza de abejas visitantes con las variables locales usadas dentro del análisis Kruskall- Wallis para evaluar su significancia. En estos análisis se incluyó a los sistemas de bosque (B1, B2, B3 y B4) como un único sistema y aunque la riqueza de abejas no se midió directamente, para estas comparaciones se considera como el total del especies de abejas registradas en la zona de estudio de acuerdo a las colecciones del LABUN y del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN 2004). El valor de cada variable dentro de las regresiones fue la media.
Resultados
Caracterización de los lugares de muestreo. El Análisis de Componentes Principales (ACP) desarrollado con las diez variables locales, fue significativo, con el 82,7% de la varianza explicada en los dos primeros componentes. En el primer eje que explica el 68,5% de la varianza, se evidencia claramente un gradiente de intensificación, con los bosques ubicados hacia la izquierda (lado negativo) y los cafetales hacia la derecha (lado positivo). Dentro de los cafetales se observan dos grupos diferenciados. Los policultivos, tanto tradicionales como comerciales, más cercanos a los bosques y los sistemas de sombrío monoespecífico con el máximo valor de intensificación (Fig. 1).
La selección de las variables que explicarían la mayor variación dentro del estudio, se basó en los valores de eigen para el primer eje del ACP (Tabla 3). Así, se seleccionaron los valores absolutos iguales o mayores de 0,30, esto permitió descartar dos variables: la densidad de árboles y el DAP de los árboles. Con las ocho variables restantes se desarrolló el análisis no paramétrico de Kruskall-Wallis.
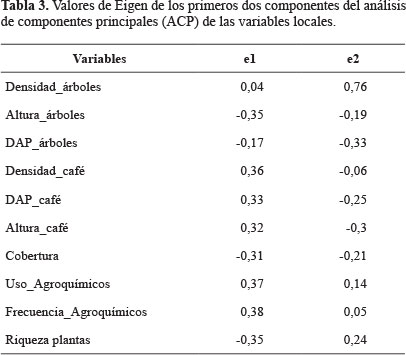
El análisis de Kruskall- Wallis mostró diferencias significativas entre los tratamientos (tipos de sistemas) para las ocho variables evaluadas (Tabla 4), Además, las pruebas de comparación entre pares de tratamientos usando las diferencias medias de rango, mostraron que no existen diferencias significativas entre las medias de las variables locales para los tratamientos PT y PC, con excepción de la densidad del café. Los parámetros más significativos, con los menores valores de p, fueron la densidad de café, la altura del café, uso de agroquímicos y frecuencia de uso de agroquímicos. Estos parámetros constituyen variables importantes para determinar el grado de intensificación y, por tanto, los cambios de la EAP en los cafetales estudiados de acuerdo a nuestro análisis.
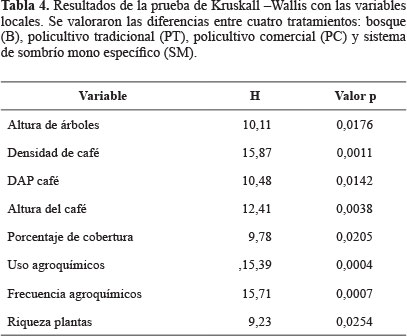
El análisis de similitud, donde se usaron las variables relacionadas con la estructura de la vegetación, mostró que los lugares de estudio se agrupan en dos conglomerados principales (Fig. 2): uno agrupa a los bosques (B1, B2, B3 y B4) y policultivos (PT1, PT2 y PC1) y el otro a los dos cultivos de sombrío mono-específico (SM1 y SM2). El conglomerado (Fig. 2) indica que en cuanto a la estructura de la vegetación policultivos y bosques son semejantes. También, se evidencia que la mayor pérdida de estructura (EAP) o el mayor grado de intensificación se relacionan con los cafetales de sombrío mono-específico (SM), los cuales son los que presentan la mayor distancia con respecto a los bosques con un índice de Bray-Curtis de 0,47.
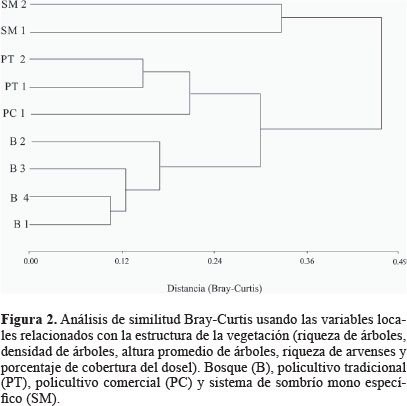
Estos resultados sugieren que para las comparaciones de diversidad de abejas, sería mejor usar solo dos tipos de cafetales: policultivos (P) y SM, eliminado la división entre policultivo comercial y tradicional, ya que estos sistemas se agruparon a una distancia menor de 0,23 (Fig. 2) y sus medias no mostraron diferencias significativas dentro del análisis de Kruskall-Wallis (Tabla 4).
Diversidad de abejas visitantes del café. Entre las abejas visitantes del café (Anexo 1), se encontraron 130 individuos de 13 especies catalogadas en tres familias: Halictidae, Megachilidae y Apidae, esta última con la mayor riqueza (nueve especies). El género más diverso fue Trigona (Jurine, 1807) con tres especies Trigona (Tetragonisca) angustula (Latreille, 1825) (Hymenoptera: Apidae), T. (Trigona) fuscipennis (Jurine, 1807) (Hymenoptera: Apidae) y T. (Trigona) amalthea (Oliver 1789) (Hymenoptera: Apidae). Por otra parte, las especies con mayor número de visitas al café (mayor número de capturas y observaciones) fueron Apis mellifera L., 1758 (Hymenoptera: Apidae) (62%) y Paratrigona eutaeniata (Camargo y Moure, 1994) (Hymenoptera: Apidae) (16%).
No se encontraron diferencias significativas en la diversidad de abejas visitantes del café entre P y SM, aunque los valores de los índices de diversidad de P son ligeramente mayores (Tabla 5). No obstante, la composición de especies en cada tratamiento fue diferente y el índice de Jaccard mostró similitud entre tratamientos de solo 2,3%.
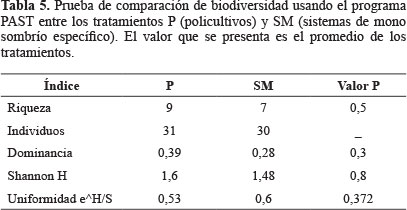
Con las variables estadísticamente significativas en el análisis de Kruskall- Wallis se hicieron regresiones lineales para indentificar cuales podrían afectar la riqueza de abejas, dado que se ha encontrado que esta tiene una relación directa con la eficiencia de la polinización en cafetales (Klein et al. 2003a; 2003b). De estas regresiones, no fueron significativas la altura promedio de los árboles (P = 0,3316 > 0,05; R2 = 0,85), el DAP promedio del café (P = 0,0763 > 0,05; R2 = 0,39), la altura promedio del café (P = 0,0792 > 0,05, R2 = 0,58), el uso de agroquímicos (P = 0,128 > 0,05, R2 = 0,48) y su frecuencia de uso (P = 0,0825 > 0,05, R2 = 0,57).
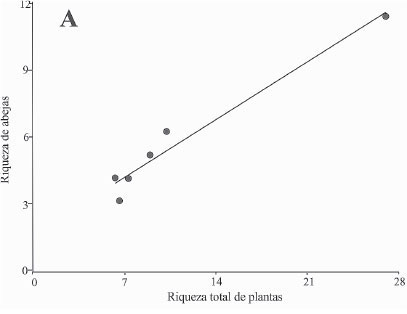
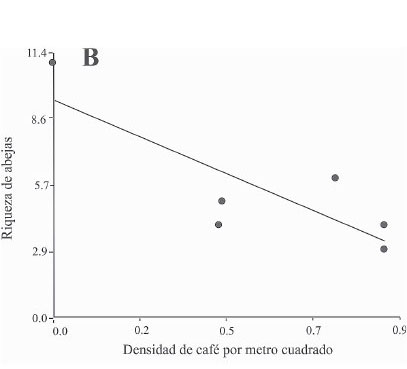
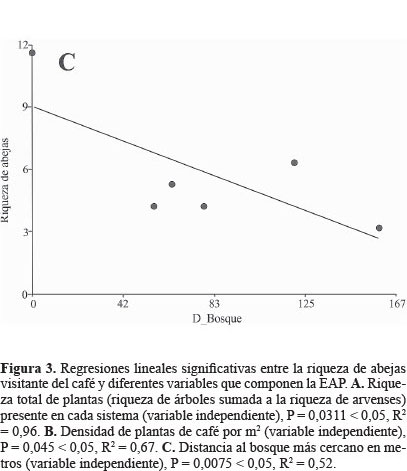
Por otra parte en la figura 3 se observan las regresiones significativas entre la riqueza de abejas visitantes y las variables de la EAP, particularmente la riqueza vegetal total (P = 0,0311 < 0,05, R2 = 0,96), la densidad de cafetos por metro cuadrado (P = 0,045 < 0,05; R2 = 0,67) y la distancia al bosque como variable asociada al paisaje (P = 0,0075 < 0,05; R2 = 0,52).
Polen de contacto. Se procesó el polen de contacto de 90 individuos de todas las especies capturadas en el muestreo. Se identificaron 11 especies de plantas, entre árboles y arbustos: Coffea arabiga, Psidium guajava, Eugenia sp., Heliocarpus americanus, Tabebuia rosea, Cecropia sp., Heisteria acuminata, Braccharis pedunculata y entre las hierbas: Balsamina ipantengs, Bidens sp. y Taraxacum officinale, así como una miscelánea de asteráceas, rubiáceas y bromelias en el polen menor.
Discusión
Caracterización de los lugares de muestreo. Muchas de las variables utilizadas para la caracterización de los cafetales han sido previamente utilizadas para este tipo de trabajo, principalmente aquellas relacionadas con la estructura de la vegetación (Moguel y Toledo 1999; Mas y Dietsch 2003), dado que parte de la literatura que trabaja el tema de café y biodiversidad no diferencia entre estructura vegetal y manejo agrícola, pues la mayor parte de los policultivos tienden a ser orgánicos, mientras los monocultivos generalmente usan mayor cantidad de agroquímicos. No obstante, este análisis muestra la importancia de incluir otras variables como la densidad del café, su altura, al igual que el uso y frecuencia de agroquímicos que pueden ser diferentes en sistemas que presenten una estructura vegetal semejante. Además, se ha mostrado que aunque la estructura vegetal se conserve, el cambio en el tipo de manejo puede implicar transformaciones de la diversidad asociada (Cepeda et al. 2009).
También es importante resaltar como actualmente la caracterización de los sistemas debe ir más allá de la finca o el cultivo y tener en cuenta el contexto paisajístico, pues tanto desde la conservación biológica como desde la agroecología se construye evidencia que sugiere que el manejo agrícola debe llevarse a cabo a nivel de paisaje para que sea exitoso (Perfecto et al. 2009).
Diversidad de abejas visitantes del café. El género Trigona, el más diverso dentro del estudio, es común en fragmentos de bosques tropicales (Siqueira et al. 2012) y en los Andes, donde este trabajo fue llevado a cabo. Algunas de sus especies se consideran muy plásticas, con amplia distribución, gracias a su capacidad para adaptarse a diferentes ambientes y forrajear diferentes fuentes de polen (González y Engel 2004; Velez-Ruiz et al. 2013). Los trabajos de Ricketts (2004) para Costa Rica y Jaramillo (2012) en Colombia reportan a Trigona spp. entre los visitantes florales más importantes. Además, Klein et al. (2003a; 2003b) consideran a las especies de este género como polinizadores claves del café tanto de C. canephora como de C. arabica.
A. mellifera fue la especie más observada, es dominante en cafetales y se considera importante para su polinización. Se ha registrado su dominancia entre un 41% - 53,8% del total de observaciones (Ricketts 2004; Jaramillo 2012). No se encontraron diferencias significativas entre el número promedio de A. mellifera observadas en P (18,7 ± 11) y SM (12 ± 11,3). Tampoco la distancia al bosque afectó el número de observaciones de esta especie (P = 0,1630 y R2 = 0,13), tendencia que coincide con los datos de Ricketts (2004) quien encontró que en un gradiente de distancia al bosque las visitas de A. mellifera fueron constantes, incrementando un poco en los lugares más distantes al bosque.
No obstante la dominancia de A. mellifera, P. eutaeniata mostró gran constancia floral (obs. pers.) y fue el segundo visitante floral más común (Anexo 1). Debido a su reducido tamaño fue difícil su captura, por lo que se considera que su abundancia estaría por debajo del valor real. Es la primera vez que se registra a P. eutaeniata como visitante de las flores del café, aunque existía un registro de su nidificación en cafetales (Fernández et al. 2010). Esta especie es además una de las más utilizadas para la meliponicultura en Colombia (Nates et al. 2009), esto la hace interesante para profundizar en estudios que involucren la conservación, la meliponicultura y la posible potenciación de la polinización en el manejo agrícola de cafetales.
P. eutaeniata se consideraba restringida a la vertiente occidental de la Cordillera Oriental de Colombia (Nates et al. 1999; Camargo y Pedro 2012), pero recientemente se registró en el Cauca y se propuso una distribución más amplia de la especie para toda la zona andina del país (Fernández et al. 2010), es decir que estaría ubicada en todas las zonas de producción de café del país. Sin embargo, Jaramillo (2012) no registra a esta o no ninguna otra especie del género Paratrigona como visitante de las flores del café o residente de los cafetales de Antioquia. Por otra parte, Rosso (2003) reporta representantes del género Paratrigona pero no a esta especie en cafetales del Valle del Cauca.
En los P se encontró mayor riqueza de meliponinos (8 especies) en contraste con SM (3 especies) y solo se compartieron dos especies de esta tribu P. eutaeniata y T. fuscipennis. Esta tendencia se relaciona con que los meliponinos prefieren troncos de árboles vivos para anidar (Siqueira et al. 2012) y dado que la especie vegetal dominante además del café en los sistemas SM es el plátano, en estos agroecosistemas se reducen las posibilidades de anidación para este tipo de abejas.
Por otra parte, todas las especies de la familia Halictidae fueron exclusivas de los SM. Esta relación también fue encontrada por Klein et al. (2003) en cafetales de Indonesia, donde la intensidad de luz aumentaba la presencia de especies solitarias, en su mayoría identificadas como halíctidos. Los SM presentaron en promedio cobertura del 20%, mientras para los P este valor fue de 67%. El trabajo de Klein et al. (2003a) también sugiere que los halíctidos prefirieron para su nidificación agroecosistemas que ofrecen áreas abiertas para anidar en el suelo y mayor número de plantas herbáceas florecidas. Nuestros datos corroboran que los SM también pueden ser hábitats favorables para estas abejas en este sentido, pues encontramos mayor riqueza promedio de arvenses dentro de los SM (7,5 ± 3) que en los P (4,9 ± 2), debido a la reducción en el número de arvenses que provoca el sombrío (Soto-Pinto et al. 2002).
El análisis de regresión entre las variables de la EAP con la riqueza de abejas (Fig. 3), apoya la idea sobre la importancia de la diversidad floral y de los espacios naturales cercanos al cultivo como variables determinantes en la diversidad de insectos polinizadores (Klein et al. 2003a; Kremen et al. 2004; Nicholls y Altieri 2012). En general, la diversidad floral al interior de los cultivos es clave para el mantenimiento de funciones ecológicas. En una reciente revisión de Letourneau et al. (2011) encontraron evidencia decisiva sobre como la diversificación de los agroecosistemas promueve la supresión de la herbivoría a través de diferentes mecanismos.
Además, se encuentra que la densidad de café por metro cuadrado afectó la riqueza de estos insectos negativamente (Fig. 3B). Esta tendencia también se ha encontrado en hormigas (De la Mora et al. 2013). Esta variable se postula como un hilo conductor que afecta la diversidad de diferentes grupos de artrópodos y puede transformar la funcionalidad ecológica de los cafetales, tal vez porque es un buen predictor del grado de intensificación agrícola en los mismos.
Los valores del R2 de las variables locales estadísticamente significativas, riqueza vegetal total y densidad de café, fueron mayores a 0,5 lo que implica que estas variables explican la mayor parte de la variación de la riqueza de abejas (Figs. 2A-2B). En contraste la distancia al bosque solo explicó el 0,52 de la variación es decir que existe casi un 50% de variación no explicada (Fig. 2C). Este resultado indica que las variables locales son mejores predictores de la variación en la riqueza de abejas, por tanto tendrían relación estrecha con la conservación de su riqueza. Estos resultados son consistentes con De la Mora et al. (2013) quienes, para hormigas en cafetales, encontraron que en general los factores locales afectaban de forma más directa y fuerte la diversidad, que los del paisaje. No obstante, la regresión estadísticamente significativa con la distancia al bosque (Fig. 2C) indica que el contexto del paisaje no puede descartarse como parte de las acciones para conservar esta fauna.
Esto es un argumento a favor de la EAP, ya que se evidencia cómo el mantenimiento de un arreglo productivo diversificado, con conectividad interna brindada principalmente por la estructura de la vegetación y la conexión con hábitats naturales adyacentes, crean efectos sinérgicos que propician la riqueza de abejas dentro del cafetal y posiblemente potencian su conservación y función ecológica como polinizadores.
De las tres variables cuya regresión lineal fue significativa, la distancia al bosque obtuvo la pendiente con un valor absoluto más alto (13,74), o sea que al incrementar la distancia al bosque en un metro se perderían aproximadamente 14 especies de abejas, lo cual confirma la importancia de los bosques para la conservación de estos polinizadores (Ricketts 2004). Los valores absolutos de la pendiente para las regresiones de la riqueza de plantas y la densidad de café fueron de 2,7 y 0,1 respectivamente.
Entre las variables que no fueron significativas dentro de la regresión lineal con la riqueza de abejas, llama la atención la no significancia tanto del uso, como de la frecuencia de agroquímicos, explicable porque los sistemas evaluados en general tienen un uso moderado de agroquímicos, entre 3 y 6, con frecuencia de aplicación de baja (2) a moderada (4). Es decir que se aplican mensual a trimestralmente, uno o dos tipos diferentes de agroquímicos, principalmente fertilizantes NPK y herbicidas (Roundup®), que pueden afectar esta fauna (Nicholls y Altieri 2012), pero no de forma tan directa como lo haría un insecticida. Aunque no hubo significancia estadística, pese al uso moderado, es importante resaltar que en ambos casos se dio una correlación negativa con la riqueza de abejas observada.
Polen de contacto. La gran diversidad vegetal encontrada dentro del polen de contacto evidencia que las abejas posiblemente visitan los diferentes estratos vegetales (árboles, hierbas y epífitas). Lo anterior sustenta por qué la relación de la riqueza de abejas es alta con la riqueza vegetal total (Fig. 3).
De los árboles identificados T. rosea (ocobo) y P. guajava (guayaba) son reportados por Echeverry (1984) como de importancia apícola. Particularmente T. rosea se considera un árbol melífero, visitado frecuentemente por abejas sin aguijón y reportado como sustrato común para la construcción de sus nidos (Arce et al. 2001). Por otra parte, P. guajava es fuente importante de alimento (polen y néctar) para las abejas, ya que presenta flores durante todo el año (op.cit.). Entre las arvenses encontradas B. ipantengs y todas especies de asteráceas, entre las que se destacan Bidens sp. y T. officinale, son consideradas por Rosso (2003) y Vivas et al. (2008) como importantes y necesarias para el sostenimiento de las poblaciones de abejas.
Todas estas plantas contribuyen a conservar a las abejas dentro de los cafetales, ya que aunque el café es una fuente importante de alimento, tiene periodos de floración restringidos y sus flores duran en promedio abiertas solo tres días (Arcila 2007). Además, Jaramillo (2012) reporta que son atractivas para los visitantes florales solo durante el primer día de apertura. Así, otras plantas dentro del arreglo productivo sirven de fuente de alimento alternativo cuando las plantas económicamente importantes no se encuentran en floración (Girón 1995) y son útiles para el sostenimiento de las poblaciones de abejas.
Las plantas mencionadas pertenecen con mayor frecuencia al arreglo del sistema productivo. Sin embargo, se encontraron otras especies más relacionadas con barbechos y utilizadas como cercas vivas en las fincas cafeteras como: Cecropia sp. (yarumo) y H. americanus (balso). También se observaron árboles encontrados en los bosques en la caracterización realizada durante este trabajo como: Eugenia sp., H. acuminata, B. pedunculata. Por tanto, se considera que los cafetales, los bosques y barbechos son aprovechados por las abejas para su alimentación y por ello su conservación es importante.
Conclusiones
Retomando las tres preguntas planteadas en la introducción y con ayuda de la evidencia encontrada a través del trabajo se concluye que:
Los cambios en la EAP afectan la diversidad de abejas visitantes del café. Las principales transformaciones se relacionan con cambios en la composición y la riqueza de estas especies.
Dentro de las variables de la EAP medidas en los sistemas cafeteros evaluados que se relacionaron con la riqueza de abejas y fueron estadísticamente significativas se destacan la riqueza de plantas total, la densidad del café y la distancia al bosque (Fig. 2). Para finalizar se encontró que las abejas usan como recursos tanto árboles como arvenses dentro del arreglo (E.j. ocobo, la guayaba y B. ipantengs), pero también a otras especies propias de los bosques y cercas vivas (E.j. Cecropia sp.). Por tanto para mantener la diversidad de abejas en los cafetales, se debe mantener la diversidad del arreglo y además pensar en la conservación de los parches de bosques cercanos.
Agradecimientos
Agradecemos a Cecilia Cantor y Andrés García por su ayuda en campo y a todos los caficultores que participaron de este proyecto. A Danny Vélez por su apoyo en la identificación de las abejas. A Tomás León y Alexander Sabogal por su lectura crítica y aportes al manuscrito. A la profesora Giomar Nates y los miembros del LABUN por el acceso a la palinoteca y su colaboración desinteresada. Este trabajo se llevó a cabo gracias al apoyo del Comité de Cafeteros de Cundinamarca, al Laboratorio de Artrópodos del Grupo de biotecnología del Centro Internacional de Física (CIF) y a la financiación brindada por el Instituto de Estudios Ambientales de la Universidad Nacional de Colombia por medio del proyecto de iniciativa universitaria. Código QUIPU 202010018127, CDP No. 337 del 22 de agosto de 2012.
Literatura citada
ANDERSON, S. H.; KELLY, D.; LADLEY, J. J.; MOLLOY, S.; TERRY, J. 2011. Cascading effects of bird functional extinction reduce pollination and plant density. Science 331 (6020): 1068-1071. [ Links ]
ARCE, G.; SÁNCHEZ, L. A.; SLAA, J.; SÁNCHEZ-VINDAS, P. E.; ORTIZ, A.; VAN VEEN, J.; SOMMEIJER, M. J. 2001. Árboles melíferos nativos de mesoamérica. Herbario Juvenal Valerio Rodríguez; Heredia, Costa Rica; 208 p. [ Links ]
ARCILA, J. 2007. Crecimiento y desarrollo de la planta de café. pp. 21-60. En: Ospina, H. F.; Marín, S. M. (Eds.). Sistemas de producción de café en Colombia. Cenicafé, Chinchiná [ Links ].
BERNAL, R.; GALEANO, G.; RODRÍGUEZ, A.; SARMIENTO, H.; GUTIÉRREZ, M. 2011. Diccionario de nombres comunes de las plantas de Colombia. Versión en línea. Instituto de ciencias naturales, Universidad Nacional de Colombia, Bogotá. Disponible en: http://www.biovirtual.unal.edu.co/diccionario/ (Fecha revisión: 10 octubre 2012). [ Links ]
CAMARGO, J. M. F.; PEDRO, S. R. M. 2012. Meliponini Lepeletier, 1836. In: Moure, J. S.; Urban, D.; Melo, G. A. R. (Orgs.). Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region. Disponible en: http://www.moure.cria.org.br/catalogue. (Fecha revisión: 2 marzo 2013). [ Links ]
CEPEDA-VALENCIA, J; SABOGAL, A; LEÓN, T. 2009. Aproximación a la diversidad de arañas y coleópteros en cultivos de café bajo manejo convencional y orgánico: Revista Brasilera de Agroecología 4 (2): 2063-2066. [ Links ]
DAFNI, A. 1992. Pollination ecology: a practical approach. IRL Press Ltd; 72-74 p. [ Links ]
DE LA MORA, A.; MURNEN, C. J.; PHILPOTT, S. M. 2013. Local and landscape drivers of biodiversity of four groups of ants in coffee landscapes. Biodiversity and Conservation, DOI 10.1007/s10531-013-0454-z. [ Links ]
DI RIENZO, J. A.; CASANOVES, F.; BALZARINI, M. G.; GONZÁLEZ, L.; TABLADA, M.; ROBLEDO, C. W. 2012. InfoStat versión 2012. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Disponible en: http://www.infostat.com.ar (Fecha revisión: 10 octubre 2012). [ Links ]
ECHEVERRY, R. 1984. Flora apícola colombiana. Biblioteca científica de la Presidencia de la República de Colombia. Primera edición. 238 p. [ Links ]
FAO. 2008. A contribution to the international initiative for the conservation and sustainable use of pollinators. FAO. 1-10p. [ Links ]
FERNÁNDEZ, D. C.; ZAMBRANO G.; GONZÁLEZ, V. H. 2010. Comportamiento de nidificación, notas taxonómicas y distribución potencial de Paratrigona eutaeniata (Hymenoptera: Apidae, Meliponini). Revista Colombiana de Entomología 36 (2): 325-332. [ Links ]
GHAZOUL, J. 2005. Buzziness as usual? Questioning the global pollination crisis. Trends Ecology Evolution 20: 367-373. [ Links ]
GIRÓN, M. 1995. Análisis palinológico de la miel y la carga de polen colectada por Apis mellifera en el suroeste de Antioquia, Colombia. Boletín del Museo de Entomología de la Universidad del Valle 3 (2): 35-54. [ Links ]
GÓMEZ, A.; RIVERA, J. H. 1995. Descripción de arvenses en plantaciones de café. CENICAFE. Colombia. [ Links ]
GONZÁLEZ, V. H.; ENGEL, M. S. 2004. The tropical Andean bee fauna (Insecta: Hymenoptera: Apoidea), with examples from Colombia. Entomologische Abhandlungen 62 (1): 65-75. [ Links ]
HAMMER, O.; HARPER, D. A. T.; RYAN, P. D. 2001. PAST: paleontological statistics software package for education and data analysis. (Paleontología Electrónica) Versión 2.12. Disponible en: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (Fecha revisión: 20 marzo 2013). [ Links ]
HERRERA, L. F.; GIRALDO, L. E. U. 1996. Atlas de polen de plantas útiles y cultivadas de la Amazonia Colombiana. Tropenbos. Colombia. [ Links ]
ICN (Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia). 2004 y continuamente actualizado. Colecciones en Línea. Disponible en: http://www.biovirtual.unal.edu.co (Fecha revisión: 10 octubre 2012). [ Links ]
JARAMILLO, A. 2012. Efecto de las abejas silvestres en la polinización del café (Coffea Arabica: Rubiaceae) en tres sistemas de producción en el departamento de Antioquia. Tesis para optar al título de Magister en entomología. Universidad Nacional sede Medellín. [ Links ]
KLEIN, A.-M.; STEFFAN-DEWENTER, I.; TSCHARNTKE, T. 2003a. Pollination of Coffea canephora in relation to local and regional agroforestry management. Journal of Applied Ecology 40:837-845. [ Links ]
KLEIN, A.-M.; STEFFAN-DEWENTER, I.; TSCHARNTKE, T. 2003b. Bee pollination and fruit set of Coffea arabica and C. canephora (Rubiaceae). American Journal of Botany 90 (1): 153-157. [ Links ]
KLEIN, A. M.; VAISSIERE, B. E.; CANE, J. H.; STEFFAN-DEWENTER, I.; CUNNINGHAM, S. A.; KREMEN, C., TSCHARNTKE, T. 2007. Importance of pollinators in changing landscapes for world crops. Proceedings of Royal Society of Biological Sciences 274: 303-313. [ Links ]
KREMEN, C.; WILLIAMS N. M.; THORP, R. W. 2002. Crop pollination from native bees at risk from agricultural intensification. Proceedings of the National Academy of Sciences of the United States of America 99: 16812-16816. [ Links ]
KREMEN, C.; WILLIAMS, N. M.; BUGG, R. L.; FAY, J. P.; THORP, R. W. 2004. The area requirements of an ecosystem service: crop pollination by native bee communities in California. Ecology Letters 7: 1109-1119. [ Links ]
LEÓN, T. 2014. Perspectiva ambiental de la agroecología: La ciencia de los agroecosistemas. IDEAS 23. Instituto de Estudios Ambientales (IDEA), Universidad Nacional de Colombia. Bogotá.151-211 p. [ Links ]
LETOURNEAU, D. K.; ARMBRECHT, I.; RIVERA, B. S.; MONTOYA-LERMA, J.; JIMÉNEZ, E.; DAZA, M. C.; ESCOBAR, S.; GALINDO, V.; GUTIÉRREZ, C.; DUQUE LÓPEZ, S.; LÓPEZ MEJÍA, J.; ACOSTA RANGEL, A. M.; HERRERA RANGEL, J.; RIVERA, L.; SAAVEDRA, C. A.; TORRES, A. M.; REYES TRUJILLO, A. 2011. Does plant diversity benefit agroecosystems? A synthetic review. Ecological Applications 21 (1): 9-21. [ Links ]
MAS, A.; DIETSCH, T. 2003. An index of management intensity for coffee agroecosystems to evaluate butterfly species richness. Ecological Applications 13 (5): 1491-1501. [ Links ]
MESA, O.; POVEDA, G.; CARVAJAL, F. 1997. Introducción al clima de Colombia. Universidad Nacional de Colombia. Medellín. 80-100 p. [ Links ]
MOGUEL, P.; TOLEDO, V. 1999. Biodiversity conservation in traditional coffee systems in Mexico. Conservation Biology 13 (1): 11-21. [ Links ]
NATES, G. 2005. Abejas silvestres y polinización. Manejo Integrado de Plagas y Agroecología (Costa Rica) 75: 7-20. [ Links ]
NATES-PARRA, G.; GONZÁLEZ, V. H.; OSPINA-TORRES, R. 1999. Descripción de los machos y anotaciones sobre la biología de Paratrigona anduzei y P. eutaeniata (Hymenoptera: Apidae: Meliponini) en Colombia. Caldasia 21 (2): 174-183. [ Links ]
NATES-PARRA, G.; ROSSO L., J.; CEPEDA, M.; LUGO, J. S. 2009. Características de la Meliponicultura en Colombia. pp. 17-23. En: Yurrita, C. L. (Ed.). Memorias VI Congreso Mesoamericano de sobre abejas nativas. Universidad de San Carlos de Guatemala - Centro de Estudios Conservacionistas-CECON. [ Links ]
NICHOLLS, C.; ALTIERI, M. 2012. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agronomy for Sustainable Development 33: 257-274. [ Links ]
PERFECTO, I.; RICE, R.; GREENBERG, R.; VAN der VOORT, M. E. 1996. Shade Coffee: A disappearing refuge for biodiversity. BioScience 46 (8): 598-608. [ Links ]
PERFECTO, I.; MAS, A.; DIETSCH, T.; VANDERMEER, J. 2003. Conservation of biodiversity in coffee agroecosystems: a tri-taxa comparison in southern Mexico. Biodiversity and Conservation 12 (6): 1239-1252. [ Links ]
PERFECTO, I.; VANDERMEER, J.; WRIGHT, A. 2009. Nature´s matrix: linking agriculture, conservation and food sovereignty. Ed. Earthscan. Londres. [ Links ]
RICKETTS, T. H. 2004. Tropical forest fragments enhance pollinator activity in nearby coffee crops. Conservation Biology 18 (5): 1262-1271. [ Links ]
RICKETTS, T.; REGETZ, J.; STEFFAN-DEWENTER, I.; CUNNINGHAM, S. A.; KREMEN, C.; BOGDANSKI, A.; GEMMILL-HERREN, B.; GREENLEAF, S. S.; KLEIN, A. M.; MAYIELD, M. M.; MORANDIN, L. A.; OCHIENG, A.; BLANDE, F. V. 2008. Landscape effects on crop pollination services: are there general patterns?. Ecology Letters 11 (5): 499-515. [ Links ]
ROSSO, J. M. 2003. Diagnóstico para el aprovechamiento y manejo integrado de abejas silvestres en agroecosistemas andinos en el Valle del Cauca. Tesis para optar al título de Zootecnista. Universidad Nacional de Colombia sede Bogotá [ Links ].
ROUBIK, D. W.; MORENO, P. J. E. 1991. Pollen and spores of Barro Colorado island. Monographs in Systematic Botany. Panamá [ Links ].
SIQUEIRA, E. N. L.; BARTELLI, B. F.; NASCIMENTO, A. R. T.; NOGUEIRA-FERREIRA, F. H. 2012. diversity and nesting substrates of stingless bees (Hymenoptera, Meliponina) in a forest remnant. Psyche: A Journal of Entomology 2012: 1-9. [ Links ]
SOTO-PINTO, L.; PERFECTO, I.; CABALLERO-NIETO, J. 2002. Shade over coffee: its effects on berry borer, leaf rust and spontaneous herbs in Chiapas, Mexico. Agroforestry Systems 55 (1): 37-45. [ Links ]
VANDERMEER, J.; PERFECTO, I.; PHILPOTT, S. M. 2010. Ecological complexity and pest control in organic coffee production: uncovering an autonomous ecosystem service. Bioscience 60 (7): 527-537. [ Links ]
VELEZ-RUIZ, R. I.; GONZALEZ, V. H.; ENGEL, M. S. 2013. Observations on the urban ecology of the Neotropical stingless bee Tetragonisca angustula (Hymenoptera: Apidae: Meliponini). Journal of Melittology (15): 1-8. [ Links ]
VIVAS, N.; MACA, J.; PARDO, M. 2008. Caracterización cualitativa del polen recolectado por Apis mellifera L. en tres apiarios del Municipio de Popayán. Biotecnología en el sector Agropecuario y Agroindustrial 6 (2): 94-98. [ Links ]
Recibido: 29-may-2014
Aceptado: 25-ago-2014