










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Entomología
versión impresa ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.2 Bogotá jul./dic. 2014
ARTÍCULOS DE INVESTIGACIÓN / RESEARCH PAPERS
Curvas de crecimiento poblacional de adultos de Hippodamia convergens y Olla v-nigrum (Coleoptera: Coccinellidae)
Population growth curves of adults of Hippodamia convergens and Olla v-nigrum (Coleoptera: Coccinellidae)
Oscar Alejandro Martínez-jaimeI; José Antonio Díaz-garcíaII; Manuel Darío Salas-araizaIII
IM. Sc. Departamento de Agronomía, División Ciencias de la Vida, Campus Irapuato-Salamanca, Universidad de Guanajuato. Ex-Hacienda "El Copal", Km. 9, carretera Irapuato-Silao; A.P. 311; C.P. 36500; Irapuato, Guanajuato, México. oscarja@ugto.mx. Autor para correspondencia
IIPos Doc. Departamento de Estadística y Cálculo, Universidad Autónoma Agraria Antonio Narro; C.P. 25315; Buenavista, Saltillo, Coahuila, México. jadiaz@uaaan.mx
IIIPh. D. Departamento de Agronomía, División Ciencias de la Vida, Campus Irapuato-Salamanca. Universidad de Guanajuato. Ex-Hacienda "El Copal", Km. 9, carretera Irapuato-Silao; A.P. 311; C.P. 36500; Irapuato, Guanajuato, México. salasm@ugto.mx
RESUMEN
A través de las técnicas de regresión múltiple multivariada se estimaron las curvas de crecimiento poblacional de dos especies depredadoras para la región de Irapuato, Guanajuato, México, mediante la forma funcional de polinomios de tercer grado definidos por:
Y1(T, PP) = 1,058e+02 - 1,300e+01*T - 7,817e-01*PP + 4,058e-01*T2 + 1,037e-02*PP2 + 3,922e-02*T*PP - 5,360e-04*T*PP2
Y2(T, PP) = 1,246e+02 - 1,761e+01*T - 8,349e-01*PP + 6,359e-01*T2 + 6,767e-02*PP2 - 1,166e-02*T*PP - 2,865e-04*T*PP2
donde T es el promedio de la temperatura media mensual, PP es el promedio de la precipitación acumulada mensual y Y1 y Y2 son, respectivamente, los números de adultos de Hippodamia convergens y Olla v-nigrum. En el análisis de varianza multivariado de la regresión múltiple multivariada, se utilizó el criterio de Roy cuyo valor de su estadístico de prueba fue de 80,379 con una probabilidad para su correspondiente aproximación F de 0,0001295, en consecuencia los polinomios estimados para las curvas de crecimiento poblacional fueron adecuados; asimismo se calcularon las medidas de asociación multivariadas de Wilks (ALH = 0,987978) y de Roy (θ = 0,9877118), las cuales permitieron recomendar las curvas de crecimiento estimadas como herramientas de predicción. Los modelos de dichas curvas de crecimiento polinómicas fueron comparados entre sí, mediante el estadístico W = 1978,3, que arrojó un valor de probabilidad P = 0**, concluyendo que las curvas de crecimiento fueron diferentes. Los puntos críticos de temperatura y precipitación coincidieron en ambas especies con el punto (13,7 °C; 2,0 mm), en el cual se obtuvieron los números máximos de individuos, resultando 3,4 para H. convergens y 2,2 para O. v-nigrum.
Palabras clave: Múltiple multivariada; Depredadores; Temperatura; Precipitación.
ABSTRACT
ABSTRACT: Population growth curves in two predator species in the region of Irapuato, Guanajuato, Mexico, were estimated using multivariate multiple regression techniques through third-degree polynomials functions defined by:
Y1(T,PP) = 1.058e+02 - 1.300e+01*T - 7.817e-01*PP + 4.058e-01*T2 + 1.037e-02*PP2 + 3.922e-02*T*PP - 5.360e-04*T*PP2
Y2(T,PP) = 1.246e+02 - 1.761e+01*T - 8.349e-01*PP + 6.359e-01*T2 + 6.767e-02*PP2 - 1.166e-02*T*PP - 2.865e-04*T*PP2
where T is the average monthly standard temperature, PP is the average monthly accumulated rainfall and Y1 and Y2 are the numbers of Hippodamia convergens and Olla v-nigrum adults, respectively. The Roy criterion was used in the multivariate analysis of variance of the multivariate multiple regression, which statistical test value was 80.379 with a probability for its corresponding approximation F of 0.0001295, consequently, the estimated polynomials for the population growth curves were adequate. Multivariate association measures Wilks (ALH = 0.987978) and Roy (θ = 0.9877118) were calculated, allowing estimation of growth curves as prediction tools. These models of polynomial growth curves were compared to each other through the statistic W = 1978.3, which yielded a probability value P = 0**, concluding that the growth curves were different. The critical point for temperature and rainfall in both species coincided with the point (13.7 °C, 2.0 mm), in which the maximum numbers of individuals were obtained, resulting 3.4 to H. convergens and 2.2 for O. v-nigrum.
Key words: Multivariate multiple regression; Predators; Temperature; Precipitation.
Introducción
Los coccinélidos se usan ampliamente en el manejo integrado de plagas, así como en programas aumentativos de control biológico, tal es el caso de Olla v-nigrum (Mulsant, 1866) que depreda al psílido de los cítricos Diaphorina citri (Kuwayama, 1908) (Hemiptera: Psyllidae) y debido a su marcada respuesta numérica es un elemento potencial de control biológico (Michaud 2001). En México, en la región del Bajío Guanajuatense, las poblaciones de la catarinita roja Hippodamia convergens (Guérin-Méneville, 1842) (Coleoptera: Coccinellidae) emigran de las zonas bajas hacia las sierras aledañas donde forman grandes agregados en la base de los árboles, posteriormente retornan a los valles en febrero (Salazar y Salas 1994), igual ocurre en el estado de California, EEUU (Lee 1980); ambas especies de coccinélidos ajustan su ciclo de vida a la disponibilidad de áfidos para su alimentación (Nicholls 2008). Algunas otras especies como Adalia bipunctata (Linneo, 1758) (Coleoptera: Coccinellidae) se adaptan a diferentes temperaturas, ya que incrementan el tamaño del cuerpo y reducen la sobrevivencia de las pupas cuando las temperaturas son bajas, siendo más susceptibles a temperaturas mayores (Sorensen et al. 2013).
La fluctuación de las poblaciones insectiles, sus interacciones tróficas y la ecología de su comportamiento, se han descrito a través de la creación de modelos ecológicos (Hilker y Westerwoff 2007). Aunque un modelo ecológico refleja las variaciones observadas en la población, no se percibirán claramente sin el uso de instrumentos analíticos que permitan estimar y/o predecir esas variaciones, considerando además las interacciones depredador-presa o huésped-parásito, que con frecuencia forman parte de los programas de manejo integrado de plagas. Lima et al. (2009) indicaron que una aproximación a los sistemas ecológicos se logra con la simplificación matemática; es decir, la creación de un modelo matemático con una estructura simple como prototipo de un sistema agroecológico, que facilite no solamente su utilización práctica, sino también su interpretación y validación.
Venette et al. (2010) afirmaron que para construir modelos que estimen con buena aproximación a las poblaciones de insectos, es necesario considerar información específica sobre los factores externos e internos que inciden en su fluctuación a través del tiempo. En cuanto a los primeros, Hodgson et al. (2011) señalaron que las variables climáticas influyen en la duración de los ciclos de vida y supervivencia de los individuos, lo que resulta en un incremento o reducción de sus poblaciones.
Se han realizado estudios que utilizan diferentes tipos de modelado en algunas especies de insectos, buscando aproximar el número de individuos considerando factores abióticos como variables independientes, como el de Acevedo et al. (2010), que con información de dos años, obtuvieron una ecuación para pronosticar la dinámica poblacional de adultos de Epilachna varivestis (Mulsant, 1850) (Coleoptera: Coccinellidae) en términos del tiempo, la edad fisiológica y la temperatura, logrando variaciones no mayores al 7% al validar dicho modelo; de igual forma, Ahumada et al. (2004) generaron un modelo de predicción del crecimiento poblacional de Culex quinquefasciatus (Say, 1826) (Diptera: Culicidae), incorporando variables como la temperatura, la precipitación y la altura sobre el nivel del mar, para comprender cómo estos agentes influyen sobre los cambios poblacionales de este díptero.
Se han construido modelos utilizando la técnica de la regresión con el fin de estimar poblaciones insectiles, cuyos predictores son precisamente variables climáticas. Por ejemplo, utilizando datos de tres años y mediante un análisis de regresión múltiple, Murphy et al. (2012) determinaron que la distribución espacial de Circulifer tenellus (Baker, 1896) (Hemiptera: Cicadellidae) depende principalmente de las variables temperatura media, precipitación y velocidad del viento. En otra investigación, Villa y Villa (1996) a través de un modelo de regresión múltiple, concluyeron que la temperatura y la precipitación explican el 89% de la variación en el número de Dendroctonus mexicanus (Hopkins, 1905) (Coleoptera: Scolytidae), utilizando capturas durante un año. Empleando registros disponibles de año y medio, Cuéllar et al. (2012) encontraron una ecuación de regresión múltiple con un coeficiente de determinación R2 = 0,64, para explicar la relación entre la abundancia de individuos del mismo escolítido con respecto a la temperatura y la precipitación pluvial. Por su parte, Gopar y Losada (2004), en dos años de conteos, estimaron la densidad poblacional de cuatro especies de curculiónidos en alfalfa, a través de modelos de regresión múltiple en función de la temperatura, la precipitación y el fotoperiodo, obteniendo los mayores coeficientes de determinación para las especies Pantomorus auripes (Hustache, 1947) (R2 = 0,56) y Aramigus tessellatus (Say, 1824) (R2 = 0,67).
Existen pocos trabajos sobre modelos que aproximen la abundancia de adultos de coccinélidos para la región de El Bajío, por lo que analizar las variaciones poblacionales de H. convergens y O. v-nigrum a través del tiempo, será un aporte más a considerar en el manejo integrado de áfidos (insectos-presa); por lo anterior, los objetivos de este trabajo fueron: (1) proponer y estimar las funciones de crecimiento poblacional de estas dos especies en términos de la temperatura y la precipitación, (2) realizar la comparación entre si de las curvas de crecimiento para las dos especies, y (3) estimar los valores de la temperatura y precipitación, para los que se alcanza el número máximo de individuos adultos.
Materiales y métodos
Periodo y área de estudio. Entre enero de 1993 a diciembre de 2002 se desarrolló el presente trabajo en el campo experimental de la División de Ciencias de la Vida (DICIVA) de la Universidad de Guanajuato (UG), ubicado en la Ex-Hacienda "El Copal", en Irapuato, Guanajuato, México; situada a una altura de 1.730 msnm, 20°40'18"N 101°20'48"O, de acuerdo con INEGI (2013); según la clasificación realizada por García (2004), el clima para esta región es del tipo BS(hw)(h)(e) semicálido subhúmedo con lluvias en verano, con una temperatura media anual de 19,6 °C y una precipitación de 575 mm anuales.
Toma de datos. Se utilizaron 12 trampas de luz negra de 15 watts de acuerdo con el diseño propuesto por Cantelo (1990); colocando una trampa cada mes en el borde de las parcelas de maíz del citado campo experimental. Los adultos de las especies H. convergens y O. v-nigrum se recolectaron mensualmente durante los diez años que incluyó este ensayo, los cuales se mataron con acetato de etilo contenido en el recipiente de captura de la trampa, la muestra se vació en una bolsa de plástico con cierre hermético para proceder a registrar el número de insectos en el Laboratorio de Entomología de la DICIVA-UG. La información climática mensual (temperatura media y precipitación pluvial acumulada) correspondiente al periodo analizado, se obtuvo de la estación meteorológica "El Copal" ubicada en el mismo campo.
Análisis de datos. Las variables dependientes fueron las 12 observaciones obtenidas con los promedios mensuales de individuos adultos de cada una de las dos especies de coleópteros registradas durante 10 años y las variables independientes fueron los promedios mensuales de temperatura media (T) en oC y de precipitación pluvial acumulada (PP) en mm, de 1993 a 2002. Primero, se realizó una prueba de independencia entre las variables dependientes para discernir si los datos serían analizados a través de técnicas univariadas o multivariadas. Enseguida, usando técnicas de regresión múltiple multivariada, se estimaron las curvas de crecimiento poblacional en forma simultánea para cada una de las dos especies de coccinélidos, ajustándolas con polinomios de tercer grado. Luego, se calcularon dos medidas de asociación multivariada, la θ de Roy y la ALH de Lawley-Hotelling. Además, se realizó la comparación de las dos curvas de crecimiento poblacionales y finalmente, mediante técnicas estándar de programación matemática, se determinaron los puntos críticos de temperatura y precipitación (T,PP) para cada curva de crecimiento, que permitieron aproximar el número máximo de individuos de cada especie.
Resultados y discusión
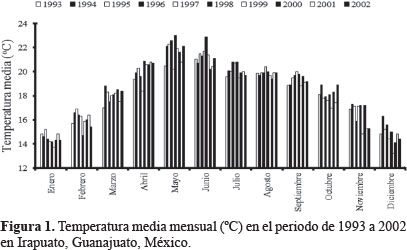
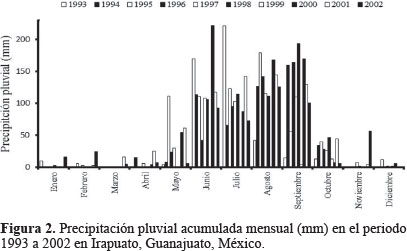
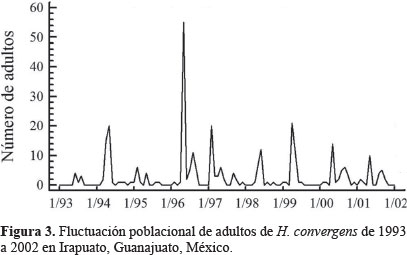
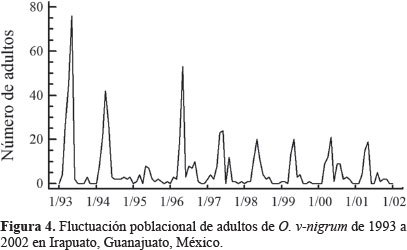
Dinámica poblacional. Olla v-nigrum presentó el mismo patrón de fluctuación que H. convergens, excepto en 1993 y 1994 cuando las poblaciones de la primera se dispararon marcadamente y las de la catarinita roja disminuyeron, al respecto Schellhorn et al. (2014) mencionaron que los enemigos naturales que viven en sistemas agrícolas, cambian constantemente por emigración e inmigración; estos flujos son esenciales para localizar al sexo opuesto, el alimento, los sitios para reproducirse y evitar la mortalidad por las prácticas agrícolas; por otro lado, la variación entre especies se presenta incluso con las que son dominantes en el mismo sitio y en diferentes años (Hodek y Honek, 1996), las poblaciones de O. v-nigrum fueron mayores que las de la catarinita roja en los primeros años de este trabajo, es posible que la temperatura de desarrollo haya influido en el incremento de las poblaciones; Hodek y Honek (1996) también observaron que cuando la temperatura para el desarrollo larvario fue de 15 °C, la longevidad del adulto fue 172 días y cuando fue de 25 °C los adultos vivieron 51 días aproximadamente, coincidiendo con los resultados obtenidos en la presente investigación, donde las temperaturas del 93 y 94 fueron bajas comparadas con los siguientes años, obteniéndose las mayores poblaciones O. v-nigrum. Rice y Wilde (1991) al comparar las poblaciones de H. convergens en trigo con diferentes sistemas de cultivo, encontraron que las poblaciones no fueron consistentes año con año, tal como ocurrió en este estudio, posiblemente la presencia de las presas influya en la aparición de esta especie, además de las condiciones de los sitios de hibernación que probablemente no fueron las más favorables para mantener las poblaciones de este depredador. Los individuos de H. convergens aumentaron su número en 1996 particularmente de marzo a mayo, cuando la temperatura se incrementó (Fig. 1); posiblemente la disponibilidad de alimento en ese año fue adecuada para que la población aumentara y disminuyó cuando las lluvias se presentaron (Fig. 2), afectando negativamente la presencia de insectos que sirven de alimento como son los áfidos; al respecto, Tarango (2008) señaló que la existencia de presas es esencial para mantener las poblaciones de esta especie en cierto hábitat; Salas et al. (1999) observaron que la incidencia más alta de áfidos en el cultivo de trigo coincide con el incremento de adultos de H. convergens para el Bajío Guanajuatense, confirmando además los resultados obtenidos por Nicholls (2008), donde la disponibilidad de pulgones determina la abundancia de estas catarinitas depredadoras, Tarango y Chávez (1997) refirieron que cuando los áfidos escasean en árboles, los adultos migran hacia arbustos de porte bajo, por lo que es posible que la falta de alimento en ciertos años haga que sus poblaciones disminuyan como ocurrió en los años 1993, 1994 y 1996; además los coccinélidos adultos son muy sensibles a las condiciones de humedad ambiental para poder emprender el vuelo migratorio, si la humedad relativa es de 0% o arriba del 77% no ocurre el vuelo (Hodek y Honek 1996). De la misma manera O. v-nigrum mostró mayor cantidad de ejemplares en 1993, 1994 y 1996 coincidiendo con los mismos meses que la catarinita roja (Figs. 3 y 4).
Curvas de crecimiento poblacionales. Se verificó si las variables respuesta (número de adultos de ambas especies) eran dependientes o independientes, mediante la obtención de la matriz de correlación y la aplicación de la prueba de independencia planteada por Rencher (2002), cuyo valor del estadístico de prueba fue U = 18,39166 con una probabilidad P = 1,798438e-05, evidenciando el rechazo de la hipótesis de independencia, lo que obligó a analizar la información a través de técnicas multivariadas, en lugar de los métodos univariados.
Además, al estimar las curvas de crecimiento poblacional, se planteó como hipótesis que los dos modelos deben tener la misma forma funcional, se procedió entonces a la estimación del mejor modelo de regresión de manera univariada para cada variable dependiente (para cada especie), basándose en los valores de la prueba de F de los análisis de varianza de la regresión y los coeficientes de determinación parciales (R2). Una vez definido el modelo de mayor grado necesario para tener el mejor ajuste, representado por el polinomio de tercer grado: Y(T,PP) = b0 + b1*T + b2*PP + b3*T2 + b4*PP2 + b5*T*PP + b6*T*PP2 , se procedió a estimar los dos polinomios que modelaron las curvas de crecimiento poblacional para la cantidad de adultos de las dos especies bajo estudio, en función de los promedios mensuales de temperatura media (T) y precipitación pluvial acumulada (PP) durante diez años a través de las técnicas de regresión múltiple multivariada, las cuales también aplicaron Murphy et al. (2012) y Villa y Villa (1996) pero para el caso univariado, demostrando su eficiencia al estimar densidades poblacionales de insectos adultos.
La prueba utilizada en el análisis de varianza multivariado (MANOVA) de la regresión múltiple multivariada, fue el criterio de Roy, cuyo valor del estadístico de prueba, la aproximación de F y su probabilidad P se presentan en la Tabla 1, lo que indicó que hay evidencia suficiente en la muestra para rechazar la hipótesis nula, concluyendo que los polinomios estimados para las curvas de crecimiento poblacional son adecuados, de forma similar a lo obtenido por Cuéllar et al. (2012) quienes, basándose en el coeficiente de determinación, lograron estimar en forma adecuada el número de adultos de D. mexicanus, pero mediante técnicas de regresión múltiple univariada.
Utilizando los algoritmos sugeridos por Rencher (2002), se calcularon las medidas de asociación multivariadas è de Roy y ALH de Lawley-Hotelling, cuyos valores permiten concluir que ambos modelos pueden usarse como herramientas de estimación (Tabla 2), estas medidas son análogos multivariados de la medida de bondad de ajuste R2 utilizada en el caso univariado. Al comparar las medidas de asociación obtenidas en este estudio (caso multivariado), con los coeficientes de determinación obtenidos por Gopar y Losada (2004) en cuatro especies de coleópteros curculiónidos (caso univariado), se concluyó que los modelos para estimar las poblaciones de H. convergens y O. v-nigrum tienen mayor grado de precisión, Rosa et al. (2011) indicaron que modelar un sistema biológico es muy difícil debido a la gran cantidad de factores que intervienen, pero es posible hacer una generalización de los resultados obtenidos, en esta investigación se aprecia que el número de individuos capturados es variable, pero su presencia en los diferentes años es constante (Figs. 3 y 4), lo que posiblemente ayudó al ajuste obtenido en los modelos, obsérvese que en algunos años los individuos de O. v-nigrum superaron a los de H. convergens. Además, para una perspectiva univariada para el lector, en la tabla 3 se presentan los polinomios de tercer grado estimados, los valores de F para los análisis de varianza (ANOVA) y los correspondientes valores de R2 y el valor de la probabilidad P para cada especie.
Para comparar los modelos de las curvas de crecimiento poblacionales polinómicos, se eliminó el parámetro b0 de cada polinomio, ya que cuando se comparan poblaciones de diferentes especies, tan sólo el b0 puede dar como resultado que los modelos sean diferentes, pues este parámetro representa el efecto del número inicial de adultos presentes de cada especie, para esta comparación se utilizó la prueba de hipótesis sugerida por Graybill (1976), mediante el estadístico W = 1978,3 con probabilidad P = 0,0, concluyendo que las curvas de crecimiento polinómicas poblacionales son diferentes, ésto de alguna manera confirma la diferencia en los hábitos de alimentación de estas especies, O. v-nigrum es de hábitos arbóreos donde las condiciones son más estables y hay inóculos de las presas para alimentarse, en este trabajo las poblaciones de esta especie se incrementaron gradualmente a diferencia de H. convergens que mostraron una decremento poblacional al inicio y posteriormente se elevaron, ésta es una especie que se desarrolla en hábitats menos estables, tales como cultivos bajos o matorrales, los cuales desaparecen anualmente, o los incendios defolian las plantas y matan las presas, sobreviven entonces alimentándose de néctar y polen. Al respecto, Tarango et al. (2001) indicaron que O. v-nigrum es afectada cuando las poblaciones de áfidos son muy bajas en las nogaleras, por lo que es conveniente inocular las huertas para incrementar las poblaciones y tener resultados satisfactorios en el control; de la misma forma, esta catarinita es muy escasa en huertas de cítricos cuando la presencia de Diaphorina citri (Kuwayama, 1908) (Heteroptera: Psyllidae) es muy baja, pero se incrementa notablemente cuando sus poblaciones se elevan (Michaud 2001), lo que hace suponer en la presente investigación, que la variación observada en los diferentes años estuvo relacionada con la presencia o ausencia de áfidos y, de manera indirecta el incremento de temperatura y precipitación pluvial elevaron las poblaciones de áfidos, sobre todo en árboles.
Por último, en la tabla 4 se muestran los puntos críticos encontrados (T, PP), los cuales establecen las condiciones climatológicas de temperatura y precipitación bajo las cuales se presenta la máxima densidad poblacional de adultos para cada una de las dos especies de coccinélidos, esto se aprecia gráficamente en las curvas de crecimiento estimadas para cada una de las dos especies estudiadas (Figs. 5 y 6).
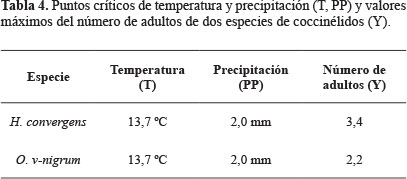
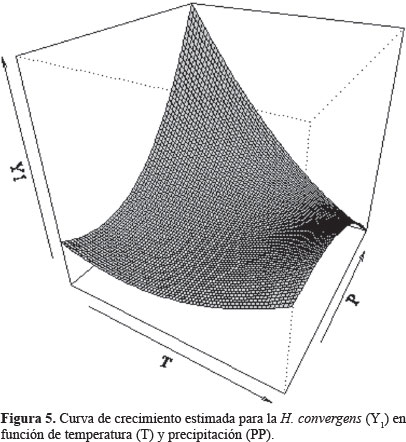
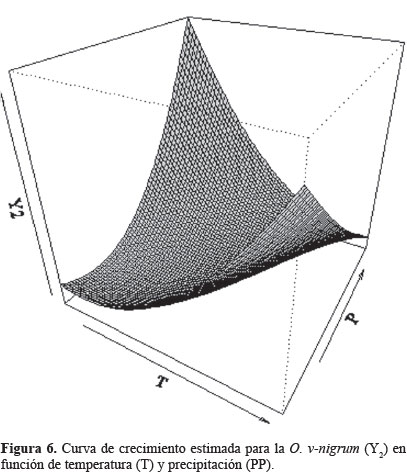
Los resultados obtenidos permiten aproximar la cantidad de estas catarinitas, que como insectos benéficos podrán incluirse en la planeación de programas más precisos de manejo integral de plagas, esta técnica la usaron eficientemente Northfield et al. (2008) con poblaciones de Frankliniella spp (Karny, 1910) (Thysanoptera: Thripidae) y la chinche Orius (Wolff, 1811) (Hemiptera: Anthocoridae) para modelar las posibles fuentes de incrementos poblacionales. Asimismo, estos resultados podrán incluirse en las alertas fitosanitarias establecidas en el estado de Guanajuato, considerando las condiciones de temperatura y precipitación para las que los enemigos naturales de los áfidos alcanzan su mayor incidencia y evitar en lo posible, la aplicación de insecticidas. La información también puede usarse como referencia en investigaciones posteriores, tendientes a determinar el umbral económico de los pulgones, basándose en la presencia de estos coccinélidos.
Conclusiones
Se estimaron las densidades de adultos de H. convergens (Y1) y O. v-nigrum (Y2) en términos de la temperatura y precipitación, a través de la obtención de curvas de crecimiento poblacionales, utilizando la técnica de regresión no lineal múltiple multivariada y proponiendo como modelos funciones polinómicas de tercer grado definidas respectivamente por:
Y1(T,PP) = 1,058e+02 - 1,300e+01*T - 7,817e-01*PP + 4,058e-01*T2 + 1,037e-02*PP2 + 3,922e-02*T*PP - 5,360e-04*T*PP2
Y2(T,PP) = 1,246e+02 - 1,761e+01*T - 8,349e-01*PP + 6,359e-01*T2 + 6,767e-02*PP2 - 1,166e-02*T*PP - 2,865e-04*T*PP2
El rechazo de la prueba de igualdad entre las curvas de crecimiento de ambas especies, posiblemente se debió a que los hábitos de ingesta en un medio efímero en alimento para la primera especie y más estable para la segunda, influyeron en la diferencia.
A partir de estas curvas de crecimiento, se calcularon los valores de 13,7 ºC de temperatura y 2,0 mm de precipitación, para los cuales se presentó el máximo número de individuos adultos de ambas especies.
Literatura citada
ACEVEDO P., A. I.; PINTO, V. M.; GARZA G., R.; RAMÍREZ A., S.; VERA G., J. 2010. Estimación del crecimiento poblacional de Epilachna varivestis Mulsant utilizando un modelo de simulación. Southwestern Entomologist 35: 557-567. [ Links ]
AHUMADA J., A.; LAPOINTE, D.; SAMUEL M., D. 2004. Modeling the population dynamics of Culex quinquefasciatus (Diptera: Culicidae), along an elevational gradient in Hawaii. Journal of Medical Entomology 41 (6): 1157-1170. [ Links ]
CANTELO W., W. 1990. Comparative efficacy of a blacklight trap and a Robinson trap in trapping moths. Southwestern Entomologist 15: 159-162. [ Links ]
CUÉLLAR R., G.; EQUIHUA M., A.; ESTRADA V., E.; MÉNDEZ M., T.; VILLA C., J.; ROMERO N., J. 2012. Fluctuación poblacional de Dendroctonus mexicanus Hopkins (Coleoptera: Curculionidae: Scolytinae) atraídos a trampas en el noreste de México y su correlación con variables climáticas. Boletín del Museo de Entomología de la Universidad del Valle 13 (2): 12-19. [ Links ]
GARCÍA, E. 2004. Modificaciones al sistema de clasificación climática de Koppen: para adaptarlo a las condiciones de la República Mexicana. Instituto de Geografía. UNAM. México, D.F. 90 p. [ Links ]
GOPAR, A.; LOSADA J., V. 2004. Estudio sobre la fluctuación poblacional de gorgojos (Coleoptera: Curculionidae) adultos que afectan a la alfalfa (Medicago sativa, L.). INTA EEA Anquil, Publicación Técnica 57: 1-20. [ Links ]
GRAYBILL, F. A. 1976. Theory and application of the linear model. Duxbury Press. Boston, Mass. EEUU. 704 p. [ Links ]
HILKER, F. M.; WESTERHOFF, F. H. 2007. Preventing extinction and outbreaks in chaotic populations. American Naturalist 170: 232-241. [ Links ]
HODEK, I.; HONEK, A. 1996. Ecology of Coccinellidae. Kluwer Academic Publishers. Dordrecht. 464 p. [ Links ]
HODGSON, J. A.; THOMAS C., D.; OLIVER T., H.; ANDERSON B., J.; BRERETON T., M.; CRONE E., E. 2011. Predicting insect phenology across space and time. Global Change Biology 17: 1289-1300. [ Links ]
INEGI. 2013. Instituto Nacional de Estadística y Geografía. Biblioteca Digital Colección InfoINEGI. México. Disponible en: http://www3.inegi.org.mx. (Fecha revisión: 14 junio 2013). [ Links ]
LEE JR., R. E. 1980. Physiological adaptations of Coccinellidae to supranivean and subnivean hibernacula. Journal Insect Physiology 26: 135-138. [ Links ]
LIMA E., A. B. F.; FERREIRA P., C.; GODOY W., A. C. 2009. Ecological modeling and pest population management: a possible and necessary connection in a changing world. Neotropical Entomology 38 (6): 699-707. [ Links ]
MICHAUD, J. P. 2001. Numerical response of Olla v-nigrum (Coleoptera: Coccinellidae) to infestations of Asian citrus psyllid (Hemiptera: Psyllidae) in Florida. Florida Entomologist 84 (4): 608- 612. [ Links ]
MURPHY, A. F.; RONDON, S. I.; JENSEN, A. S. 2012. Population dynamics of the beet leafhopper (Hemiptera: Cicadellidae) in the Columbia Basin as influenced by abiotic variables. Environmental Entomology 41: 768-775. [ Links ]
NICHOLLS E., C. I. 2008. Control biológico de insectos: un enfoque agroecológico. Editorial Universidad de Antioquia. Medellin, Colombia. 278 p. [ Links ]
NORTHFIELD T., D.; PAINI D., R.; FUNDERBURK J., E.; REITZ S., R. 2008. Annual cycles of Frankliniella spp (Thysanoptera: Thripidae) thrips abundance on north Florida uncultivated reproductive hosts: predicting possible sources of pest outbreaks. Annals of the Entomological Society of America 101 (4): 769-778. [ Links ]
RENCHER A., C. 2002. Methods of multivariate analysis. Wiley, New York. EEUU. 738 p. [ Links ]
RICE M., E.; WILDE G., E. 1991. Aphid predators associated with conventional and conservation-tillage winter wheat. Journal of the Kansas Entomological Society 64: 245-250. [ Links ]
ROSA G., S; COSTA M., I. S.; CORRENTE J., E.; SILVEIRA A., L. V.; GODOY C., W. A. 2011. Population dynamics, life stage and ecological modeling in Chrysomya albiceps (Wiedemann) (Diptera: Calliphoridae). Neotropical Entomology 40 (2): 181-189. [ Links ]
SALAS A., M. D.; SALAZAR S., E.; MARTÍNEZ S., A. M. 1999. Fluctuación poblacional de los áfidos del trigo y sus enemigos naturales en El Bajío, México. Revista Manejo Integrado de Plagas 52: 58-64. [ Links ]
SALAZAR S., E.; SALAS A., M. D. 1994. Análisis preliminar de la entomofauna en las sierras aledañas al Bajío Guanajuatense. Acta Universitaria 4 (2): 18-24. [ Links ]
SCHELLHORN N., A.; BIANCHI F. J. J., A.; HSU C., L. 2014. Movement of entomophagous arthropods in agricultural landscapes: links to pest suppression. Annual Review of Entomology 59: 559-581. [ Links ]
SORENSEN C., H.; TOFT, S.; KRISTENSEN T., N. 2013. Cold-acclimation increases the predatory efficiency of the aphidophagous coccinellid Adalia bipunctata. Biological Control 65 (1): 87-94. [ Links ]
TARANGO R., S. H.; CHAVEZ S., N. 1997. Control natural de áfidos amarillos (Homoptera: Aphididae) en nogal pecanero. Vedalia 4: 3-7. [ Links ]
TARANGO R., S. H.; QUIÑONES P., F.; CHAVEZ S., N. 2001. Control biológico de pulgones del nogal con catarinitas. SAGARPA. INIFAP. Campo Experimental Delicias. Desplegable para Productores No. 26. Chihuahua, México. 4 p. [ Links ]
TARANGO R., S. H. 2008. Bioecología de catarinitas afidófagas (Coleoptera: Coccinellidae) en árboles frutales. pp. 99-112. En: Salas A., M. D.; Salazar S., E. (Eds.). Entomófagos en el control de plagas agrícolas en México. Universidad de Guanajuato. Guanajuato, México. [ Links ]
VENETTE R., C.; KRITICOS D., J.; MAGAREY R., D.; KOCH F., H.; BAKER R., H. A.; WORNER S., P.; GOMEZ R., N. N.; MCKENNEY D., W.; DOBESBERGER E., J.; YEMSHANOV, D.; DE BARRO P., J.; HUTCHISON W., D.; FOWLER, G.; KALARIS T., M.; PEDLAR, J. 2010. Pest risk maps for invasive alien species: a roadmap for improvement. BioScience 60 (5): 349-362. [ Links ]
VILLA C., J.; VILLA C., J. 1996. La ubicación de trampas y factores climáticos afectan el monitoreo de descortezadores en el sur del estado de Jalisco. Revista Ciencia Forestal en México 21 (79): 87-100. [ Links ]
Recibido: 14-feb-2014
Aceptado: 3-nov-2014
