










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.40 no.2 Bogotá July/Dec. 2014
NOTA CIENTÍFICA /SCIENTIFIC NOTE
Description of the female of the enigmatic Batrachophthalmum quimbaya (Diptera: Richardiidae)
Descripción de la hembra de la enigmática Batrachophthalmum quimbaya (Diptera: Richardiidae)
Lisiane Dilli WendtI,II and Rosaly Ale-RochaI,III
I Ph. D. Instituto Nacional de Pesquisas da Amazônia (INPA), Coordenação de Pesquisas em Biodiversidade, Av. André Araújo, 2936, Petrópolis, CEP 69011-970, Manaus, Amazonas, Brasil.
II Fellowship CAPES/FAPEAM, Brazil. lisidilli@yahoo.com.br. Corresponding author.
III Ph. D. Fellowship PQ/ CNPq, Brazil. alerocha@inpa.gov.br
Abstract
Batrachophthalmum hendel is a small genus of Richardiidae, with only two species: B. rufiventre, from Peru and B. quimbaya, from Colombia. The male of B. quimbaya has an extraordinary head, the width from one eye to another, can be almost three times the length of the body. The female of B. quimbaya was unknown. Herein, the female of B. quimbaya is described, which presents remarkable differences from the male, especially in the head. Description and illustration of the male and female terminalia of Batrachophthalmum are also provided.
Key words: Neotropical Region. Richardiinae. Sexual dimorphism. Stalked-eye.
Resumen
Batrachophthalmum hendel es un pequeño género de Richardiidae, con sólo dos especies: B. rufiventre, de Perú y B. quimbaya, de Colombia. El macho de B. quimbaya tiene una cabeza extraordinaria, el ancho de un ojo al otro, puede ser casi tres veces la longitud del cuerpo. La hembra de B. quimbaya era desconocida. Aquí, se describe la hembra de B. quimbaya, que exhibe diferencias notables con el macho, especialmente en la cabeza. También se proporcionan descripción y ilustración de la terminalia masculina y femenina de Batrachophthalmum.
Palabras clave: Región Neotropical. Richadiinae. Dimorfismo sexual. Ojos pedunculados.
Introduction
Richardiidae is a relatively small family of Tephritoidea (Diptera), with about 190 species (Steyskal 1968; Wendt and Ale-Rocha 2014, in press) distributed in 32 genera (Hancock 2010). The family is predominantly Neotropical, and currently 14 genera and 19 species are known to occur in Colombia (Steyskal 1968; Wendt and Ale-Rocha unpublished data).
Batrachophthalmum Hendel is a poorly known genus of Richardiidae, with two species: B. rufiventre Hendel, from Peru, described based on a single female (Hendel 1911); and B. quimbaya Carvalho, Wolff and Wendt, an enigmatic species from Colombia, recently described based on two males (Carvalho et al. 2011). The genus can be recognized mainly by all femora with spine-like anteroand posteroventral setae; setae on fore femur distinctly stronger than those on mid and hind femora; mid and hind legs longer and more slender than fore leg; vein R sinuous just before margin;crossvein r-m at distal three-fourths of cell dm; metasternum bare; and postcoxal metathoracic bridge completed (Carvalho et al. 2011).
Sexual dimorphism in the head is not frequently found in species of Richardiidae. These sex-related morphological differences have only been documented for species of Richardia Robineau-Desvoidy, especially those with antlerlike genal process (Enderlein 1912; Wendt and Ale-Rocha, in press) and stalked-eyes, and for species of Megalothoraca Hendel and Batrachophthamum (Carvalho et al. 2011). The male of B. quimbaya has an enigmatic stalked-eye: the width of the head, from one eye to another, can be almost three times the length of the body. To date, the female was unknown. In a recent visit to Colección Entomológica, Universidad de Antioquia, Medellín, Colombia (CEUA), we found one female specimen of B. quimbaya, which presents a remarkable feature in the head. Herein, we describe it, and describe and illustrate the male terminalia of the species for the first time.
Material and methods
All studied material was obtained from the Colección Entomológica, Universidad de Antioquia, Medellín, Colombia (CEUA). The external morphological terminology follows Cumming and Wood (2009), except female and male terminalia, we followed Norrbom and Kim (1988) and White et al. (1999), respectively. Measurements of the head are presented as proportions, which were calculated always with the head in frontal view, as follows: (I) head width-to-height ratio: the greatest width of the head, including the eyes, divided by the greatest height of the head, measured from the vertex to the ventral limit of face; and (II) frons-to-head ratio: the greatest width of frons divided by the greatest width of head.
For the study and drawing of the terminalia, the abdomen was detached and treated according to the protocol of Cumming (1992). The dissected abdomens were stored in plastic microvials containing glycerin and pinned with their corresponding specimen.
Batrachophthalmum quimbaya
(Figs. 1A-1F, 2A-2K and 3A-3I)
Batrachophthalmum quimbaya Carvalho, Wolff and Wendt, 2011: 44 (Figs. 6-10, habitus, head, wing, thorax). Holotype and paratype males, CEUA. Type locality: Colombia, Risaralda. Pereira, Santuario de Flora y Fauna Otún/ Quimbaya.


Diagnosis and comments. The female of B. quimbaya is easily differentiated from the female of Batrachophthalmum rufiventre, by having ocellar seta reduced; orbital seta present; lateral vertical seta reduced, almost indistinguishable from setulae around it; one dorsocentral seta; wing with wide and discontinuous median band; abdomen and legs brown, except mid tarsomeres and apical half of hind tibia yellow and hind tarsomeres whitish. By contrast, B. rufiventre has ocellar and lateral vertical setae developed; orbital seta absent; two dorsocentral setae; wing with central, narrow and continuous median band (see Carvalho et al. 2011, fig. 1); abdomen and legs brownish yellow, except coxae brown.
The most remarkable difference between the male and female of B. quimbaya is the absence of stalked-eyes in the female. In males, the width-to-height ratio of the head is 10.0, and the eye is reduced, occupying only 4% of it; in the female, by contrast, this ratio is 1.57 and the eye width occupying about 60% of the head, in frontal view. Moreover, the female has lateral vertical seta reduced, almost indistinguishable from the setulae around it; clypeus yellow; and sternites 3-6 with small anterior apodemes (Figs. 2H-2J). However, males and females are identical in the general coloration of the body (especially the legs), thoracic chaetotaxy, wing pattern of spots and wing venation.
Description female. Measurements (in mm): body length 6.7; abdomen 2.6; wing 7.4. Head (Figs. 1C-1E): Frons pale brown, weakly covered with whitish pollinosity, except two triangulate glossy brown mark, from below of orbital seta extending to occiput. Ocellar triangle brown. Parafacial and anterior half of gena brownish yellow. Posterior half of gena, lunule and face brown. Clypeus yellow; developed and protruded, in lateral view, exceeding the face level (Fig. 1D). Postcranium, postgena and prementum brown. Palpus brownish yellow and triangulate (obtuse scalene shape) (Fig. 1D). Scape pale brown, as long as pedicel. Pedicel and first flagellomere dark brown. First flagellomere 2.5 times longer than wide (Fig. 1D). Arista dark brown, except sub-base yellow; with pubescence relatively dense and long (Fig. 1D). Head width-to-height ratio 1.57 (Fig. 1C). Frons-to-head ratio 0.42 (Fig. 1C). Ocellar and postocellar setae present, very weak, distinctly weaker than orbital seta. Orbital seta shorter than medial vertical seta; inserted slightly above the level of ocellar seta insertion. Genal and postgenal setae absent. Lateral vertical seta very reduced, almost indistinguishable from setulae around it. Thorax (Figs. 1A-1B and 1F): Entirely dark brown, except inconspicuous median brown stripe on scutum. Scutum 1.5 times as long as wide (Fig. 1F), with yellowish setulae. Postpronotal seta absent. Anterior notopleural seta thicker than posterior seta. Intra-alar distinctly smaller than postalar seta. Scutellar setae 2, basal seta distinctly smaller than apical seta. Katepisternal seta absent. Proepimeral seta weak. Halter yellowish on basal half and dark brown on apical half. Legs (Fig. 1A): Fore coxa, trochanter, femur and tibia brown; fore tarsomeres brownish yellow. Mid and hind coxae dark brown. Mid and hind trochanters and femora, mid tibia and basal half of hind tibia brown; mid tarsomeres and apical half of hind tibia yellow. Hind tarsomeres whitish. Fore femur with spine-like setae distinctly stronger and longer than those on mid and hind femora; with two and three spine-like setae on anteroand posteroventral surfaces, respectively, on apical third. Mid and hind femora long, 2.0 times as long as fore femur; with weak spine-like anteroand posteroventral setae on apical fourth. Hind femur without preapical dorsal seta. Wing: Identical to male: mostly yellowish, with three brown marks: one faint sub-basal spot, one wide irregular and discontinuous median band, with two hyaline areas, one subcircular on apical portion of cell dm and another more elongate on basal portion of cell r , and one irregular apical spot covering apices of cells r ,punctuated (Figs. 2B and 2E). Setae on cercus: two sub-basal setae, longer and thinner than preapical setae; two very short postmedian setae; two long preapical setae (Figs. 2B and 2E). Two spherical and strongly sclerotized spermathecae (Fig. 2C; only one was photographed).
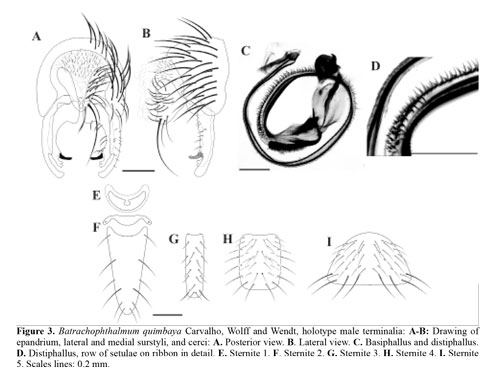
Description of male sternites (Figs. 3E-3I) and terminalia (Figs. 3A-3D): Sternite 1 present, slightly more sclerotized centrally (Fig. 3E). Sternites 2-6 with setulae very weak, except some apical stronger setulae on sternites 4-5 (Figs. 3F-3I). Sternite 2 elongate, divided into two plates (Fig. 3F). Sternites 3-5 without apodemes (Figs. 3G-3I). Sternite 3 distinctly longer than wide, rectangular (Fig. 3G). Sternite 4 slightly longer than wide, subquadrangular (Fig. 3H). Sternite 5 wider than long (Fig. 3I). Epandrium slightly wider than high, densely covered with long setae (Figs. 3A3B). Cerci developed and fused only in the lower edge (Figs. 3A-3B); in posterior view, heart-shaped (Fig. 3A); slightly sclerotized; densely ciliated. Lateral surstylus slightly longer than medial surtylus; with postmedial projection, and basal two-thirds wider than apical third; slightly ciliated (Figs. 3A3B). Medial surstylus wider at base; bilobed at apex (Figs. 3A-3B). Prensiseta preapically inserted, developed and long, rounded at apex (Figs. 3A-3B). Distiphallus, on basal half, with fringe of dense and reduced setulae (Fig. 3C); on median to preapical region with rows of conspicuous spine-like setulae (Figs. 3C-3D); on preapical region with dense fringe of very thin setulae (Fig. 3C); apex strongly sclerotized (Fig. 3C).
Examined material: CoLoMBIA. Risaralda. Santuario Fauna y Flora Otún-Quimbaya. 1800 m. Bosque Nativo. Van Someren-Ryon Trap with fish. 1PM. 07.v.2001. Group Entomology col. CEUA16283 [internal collection code] (1♀, CEUA). Compared to holotype and paratype males. CoLoMBIA. Risaralda Pereira. Santuario de Flora y Fauna Otún-Quimbaya. VSR trap [Van Someren-Ryon Trap]. Bosque [Forest]. v.2001. Entomology Group col. CEUA 12796 (CEUA).
Discussion
Even though B. quimbaya females differ from their male counterparts, it is not difficult to associate both sexes, since they are similar in the general coloration of the body, especially the contrasting coloration of the hind tibia and tarsomeres relative to the rest of the legs, and the pattern of spots on the wing.
Within Diptera, stalked-eyes are present in eight families: Diopsidae, Drosophilidae, Micropezidae, Periscelidadae, (Figs. 1A-1B): Tergites and sternites dark brown. Sternite 1 present, bare and slightly developed (Fig. 2F). Sternites 2-6 with setulae very weak, except some apical setulae stronger on sternites 3-6 (Figs. 2G-2K). Sternite 2 elongate, divided into two plates (Fig. 2G). Sternites 3-5 longer than wide, subrectangular, with two basal and vestigial apodemes (Figs. 2G-2J). Sternite 6 wider than long, without apodemes (Fig. 2K). Terminalia: Oviscape brown, 0.95 mm long (Fig. 2A). Taenia light brown, distinctly less sclerotized than oviscape; 0.5 times as long as oviscape (Fig. 2A). Eversible membrane brownish. Sternite and tergite 8 densely ciliated (Fig. 2E); sternite 8 striated, 0.75 mm long, slightly shorter than tergite 8 (Fig. 2D). Tip of cerci rounded. Cerci slightly (Wilkinson and Dodson 1997). Among the superfamily Tephritoidea, stalked-eyes seem to have evolved in one or more independent evolutionary events within each family (Wilkinson and Dodson 1997). In Richardiidae, stalked-eyes are found in Richardia, Megalothoraca and Batrachophthalmum and, according to preliminary phylogenetic analysis in preparation, they have evolved independently within each genus.
In Tephritoidea, modifications of the head, including antler genal projections, are commonly limited to males, which suggests that they play a role in sexual selection (Wilkinson and Dodson 1997). At least in acalyptrates, these modifications are generally not directly used for mating, but seem to be associated with male-male agonistic interactions (McAlpine 1982). For example, a number of ulidiid and platystomatid males have their eyes on stalks, which may be used to defend oviposition sites from sexual rivals (Wilkinson and Dodson 1997). In addition, males of Phytalmia Gerstaecker (Tephritidae), which have the most elaborated genal projections among acalyptrates, try to maintain exclusive possession of oviposition sites and resources on the tree surface to which females will be attracted (Moulds 1977; Dodson 1997). However, the role of stalked-eyes in the sexual behavior of Richardiidae remains poorly understood. The parallel development of antlers and stalked eyes in several groups may be ascribed to similar opportunities to defend resource and/or territory. Consequently, knowledge on these groups may help us understand the behavioral evolution of stalk-eyed richardiids.
Acknowledgements
Our sincere thanks to Marta Wolff (CEUA) for lending the specimens. We also thank Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for supporting the Project “Incremento do conhecimento da diversidade, formação e fixação de especialistas em Sistemática de Diptera (Insecta) na Amazônia”. We are also grateful to CAPES, FAPEAM and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the scholarships provided. Automontage equipment was made available to us through the project “Amazonas: Diversidade de insetos ao longo de suas fronteiras” (PRONEX/FAPEAM Proc. 1437/2007/NTO318/07) headed by Dr. José Albertino Rafael.
Literature cited
CARVALHO, C. J. B. DE; WOLFF, M.; WENDT, L. D. 2011. A new stalk-eyed species of Batrachophthalmum Hendel (Diptera, Richardiidae) from Colombia. Zootaxa 2932: 41-46. [ Links ]
CUMMING, J. M. 1992. Lactic acid as an agent for macerating Diptera specimens. Fly Times 8: 7. [ Links ]
CUMMING, J. M.; WOOD, D. M. 2009. Morphology and terminology. pp. 9-50. In: Brown, B. V.; Borkent, A.; Cumming,J. M.; Wood, D. M.; Woodley, N. E.; Zumbado, M. A. (Eds.). Manual of Central American Diptera.Vol. 1. NRC Research Press. Ottawa. Canada. 714 p. [ Links ]
DODSON, G. N. 1997. Resource defense mating system in antlered flies, Phytalmia spp. (Diptera: Tephritidae). Annals of the Entomological Society of America 90: 496-504. [ Links ]
ENDERLEIN, G. 1912. Die Richardiiden des Stettiner Museums.Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin 1912: 99-113. [ Links ]
HANCOCK, E. G. 2010. Family Richardiidae (Richardiid flies). pp.871-879. In: Brown, B. V.; Borkent, A.; Cumming, J. M.; Wood,D. M.; Woodley, N. E.; Zumbado, M. A. (Eds.). Manual of Central American Diptera.Vol. 2. NRC Research Press. Ottawa. Canada. p. 715-1442. [ Links ]
HENDEL, F. 1911. Die arten der dipteren-subfamilie Richardiinae. Deutsche Entomologische Zeitschrift 1911: 181-212, 239-270; 367-396. [ Links ]
MCALPINE, D. K. 1982. The acalyptrate Diptera with special reference to the Platystomatidae. p. 659-673. In: Gressitt, J. L. (Ed.). Biogeography and Ecology of New Guinea, Dr. W. Junk Publishers, The Hague. 983 p. [ Links ]
MOULDS, M. S. 1977. Field observations on behaviour of a North Queensland species of Phytalmia (Diptera, Tephritidae). Journal of the Australian Entomological Society 16: 347-352. [ Links ]
NORRBOM, A. L.; KIM, K. C. 1988. Revision of the schausi group of Anastrepha Schiner (Diptera: Tephritidae), with a discussion of the terminology of the female terminalia in the Tephritoidea. Annals of the Entomological Society of America 81: 164-173. [ Links ]
STEYSKAL, G. C. 1968. Famly Richardiidae. pp. 1-20. In: Vanzoline, E. P.; Papavero, N. (Eds.). A catalogue of the Diptera of the Americas South of the United States. Departamento de Zoologia, Secretaria da Agricultura, São Paulo, Brazil. [ Links ]
WENDT, L. D.; ALE-ROCHA, R. 2014. Taxonomic revision of wasp-like richardiids, Neoidiotypa Osten-Sacken (Diptera, Tephritoidea, Richardiidae). Zootaxa 3857: 527-550. http://dx.doi.org./10.11646/zootaxa.3857.4.4 [ Links ]
WENDT, L. D.; ALE-ROCHA, R. in press. Antlered flies richardiids: new species of Richardia Robineau-Desvoidy (Tephritoidea, Richardiidae) with antler-like genal process. Entomological Science. [ Links ]
WHITE, I. M.; NORRBOM, A. L.; HEADRICK, D. H.; CARROLL, L. E. 1999. Glossary. pp. 881-924. In: Aluja, M.; Norrbom, A. L. (Eds.). Fruit flies (Tephritidae): Phylogeny and evolution of behavior. CRC Press, Boca Raton. Washington. USA. 944 p. [ Links ]
WILKINSON, G.; DODSON, G. N. 1997. Function and evolution of antlers and eye-stalks in flies. pp. 310-328. In: Choe, J. C.; Crespi, B. J. (Eds.). The evolution of mating systems in insects and Arachnids. Cambridge University Press. Cambridge. United Kingdom. 440 p. [ Links ]
Received: 26-Aug-2014
Accepted: 22-Oct-2014
