










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Entomología
versão impressa ISSN 0120-0488
Rev. Colomb. Entomol. vol.41 no.1 Bogotá jan./jun. 2015
Visitantes florales y polinizadores en poblaciones silvestres de agraz (Vaccinium meridionale) del bosque andino colombiano
Floral visitors and pollinators in wild population of Andean blueberry (Vaccinium meridionale) in Colombian Andean forest
MARIO SIMÓN PINILLA-GALLEGO1 y GUIOMAR NATES-PARRA2
1 Biólogo. Universidad Nacional de Colombia, sede Bogotá. Carrera 45 No 26-85, Bogotá, Colombia. simonpinilla@yahoo.com. Autor para correspondencia.
2 M. Sc. Universidad Nacional de Colombia, sede Bogotá. Carrera 45 No 26-85, Bogotá, Colombia. mgnatesp@unal.edu.co.
Resumen: El agraz (Vaccinium meridionale) es una planta nativa de Suramérica con alto potencial para el consumo nacional e internacional. El objetivo de este trabajo fue identificar los visitantes florales y potenciales polinizadores del agraz en dos zonas productoras de este fruto. El estudio se realizó en cuatro puntos de muestreo distribuidos en los municipios de Guachetá (Cundinamarca), Ráquira y San Miguel de Sema (Boyacá), durante la primera floración en 2012, con cuatro visitas en cada sitio. Se registraron los visitantes florales entre las 06:00 y 17:00 h, se tomaron datos de comportamiento de los visitantes y se obtuvo muestras de polen del cuerpo. Se calculó el Índice de Probabilidad de Polinización (PPI) de los principales visitantes, y el aporte al servicio de polinización con el Índice de Importancia de Polinización (PII). Se identificaron 11 especies de insectos visitantes, de las cuales Apis mellifera, Bombus hortulanus y B. rubicundus fueron las más frecuentes en todas las zonas de muestreo. Ambas especies de Bombus vibran en las flores para extraer polen, siendo B. hortulanus el visitante que más polen de agraz transporta. Apis mellifera visita las flores para recolectar néctar, aunque también puede transportar polen. El aporte a la polinización del agraz por parte de los principales visitantes varía en las diferentes zonas de muestreo, siendo mayor el de A. mellifera en Guacheta-Ráquira, y el de B. hortulanus en San Miguel de Sema.
Palabras clave: Abejas silvestres. Bombus hortulanus. Bombus rubicundus. Polinización. Polinizadores silvestres.
Abstract: Andean blueberry (Vaccinium meridionale) is a native plant from South America with a high potential for national and international use. The aim of this study was to identify the floral visitors and potential pollinators of the Andean blueberry in two growing areas. The study was done in four samplings areas distributed in Guachetá (Cundinamarca), Ráquira and San Miguel de Sema (Boyacá) during the first blooming season of 2012. The floral visitors were recorded between 06:00 and 17:00 h, visitors behavior was recorded and pollen samples obtained from their bodies. The Pollination Probability Index (PPI) of the main visitors was calculated, and also the contribution to the pollination service through the Pollination Importance Index (PII). A total of 11 insect species was identified visiting Andean blueberry, of which Apis mellifera, Bombus hortulanus and B. rubicundus were the most frequently in all sampling areas. Both species of Bombus vibrate flowers to remove pollen, been B. hortulanus the visitor that transported the most Andean blueberry pollen. Apis mellifera visits flowers to gather nectar, but it can also transport Andean blueberry pollen. The contribution of the main visitors to the Andean blueberry pollination varies between sampling areas, been highest the contribution of A. mellifera in Guacheta-Ráquira, and the B. hortulanus in San Miguel de Sema.
Key words: Wild bees. Bombus hortulanus. Bombus rubicundus. Pollination. Wild pollinators.
Introducción
Dentro del género Vaccinium (Ericaceae) se encuentran las plantas conocidas como "blueberries" y "cranberries", las cuales se cultivan principalmente en zonas templadas y tienen un alto valor económico (Judd et al. 2007). Sus flores suelen tener la corola acampanada, el androceo de 8 a 10 estambres, anteras poricidas y el polen permanece unido en té-tradas, el gineceo presenta un estigma, y el fruto es una falsa baya (Ligarreto 2009).
Algunas especies de Vaccinium son autofértiles y otras presentan autoincompatibilidad (Griffin y Blazich 2008), aunque en las primeras su producción mejora con la polinización cruzada, lo que resalta la importancia de los polinizado-res en ellas (Janick y Paull 2008). Los visitantes florales más comunes en cultivos de Vaccinium en Estados Unidos son Apis mellifera L., 1758 (Hymenoptera: Apidae), Bombus spp. Latreille, 1802 (Hymenoptera: Apidae), Xylocopa spp. La-treille, 1802 (Hymenoptera: Apidae) y Habropoda laboriosa Fabricius, 1804 (Hymenoptera: Apidae) (Dedej y Delaplane 2003; Ratti et al. 2008). De éstos, los más eficientes en cuanto a polinización son Bombus spp. y H. laboriosa, debido a que visitan las flores para recolectar polen y tienen la capacidad de usar el método de "polinización por zumbido", lo cual libera el polen de las anteras poricidas (Mackenzie 1994). Gracias a ello, una visita de Bombus spp. puede depositar 4,4 veces más polen en el estigma que una de A. mellifera en flores de V. angustifolium (Javorek et al. 2002). El servicio de la polinización de H. laboriosa en cultivos de V. ashei se ha estimado entre 18-21 dólares americanos por abeja (Cane 1997).
Ratti et al. (2008) encontraron una correlación positiva entre el número de visitas de Bombus spp. y el peso de los frutos en cultivos de Vaccinium corymbosum y V. macrocar-pon, y en algunos casos la polinización mediada exclusivamente por Bombus es suficiente para proveer una adecuada polinización en cultivos de arándanos de pequeña y mediana escala (Ratti 2006). A pesar de ello, en los cultivos a gran escala, las abejas silvestres suelen ser escasas por lo que para garantizar la polinización se instalan colmenas de A. mellife-ra. El gran número de visitas de esta especie suple su falta de especificidad en las visitas individuales (Dedej y Delaplane 2003).
En Colombia, el agraz o mortiño (V meridionale Swartz, 1788) presenta alto potencial para el consumo nacional y ha sido incluido en la lista de especies con mercado hacia Estados Unidos. La demanda y producción de esta fruta han subido sostenidamente en los últimos años (Torres et al. 2009). El interés en esta fruta se debe principalmente a su alto contenido de antocianinas y antioxidantes, a los cuales se le atribuyen numerosas propiedades terapeúticas (Gaviria et al. 2009). A pesar de esto, en Colombia no existen cultivos de agraz, sino poblaciones silvestres que son explotadas por las comunidades locales en zonas de bosque premontano y montano, en pequeñas parcelas y matorrales, principalmente en los departamentos de Boyacá, Antioquia y Cundinamarca (Muñoz et al. 2009). A pesar de la importancia de los polini-zadores en este tipo de plantas, para el agraz no se dispone de estudios sobre sus requerimientos de polinización ni sobre sus polinizadores, por lo que el objetivo de este estudio fue identificar sus visitantes florales y polinizadores potenciales en dos regiones productoras de Cundinamarca y Boyacá (Colombia).
Materiales y métodosÁrea de estudio. Se seleccionaron dos zonas, separadas entre sí aproximadamente 9 km y, en cada una, dos localidades de muestreo en donde se encontraran plantas de agraz en alta densidad. La primera zona (G-R), se ubicó dentro de la Reserva Forestal Protectora El Robledal, área de conservación de bosques andinos de roble (Quercus humboldtii), donde el agraz crece de forma silvestre en zonas de matorrales, potreros y cultivos abandonados. La primera localidad correspondió al municipio de Guachetá (Cundinamarca), vereda Ga-chetá Alto (G-Gachetá alto) (5°27'33"N 73°39'51"O) a 2.950 msnm, y la segunda localidad al municipio de Ráquira (Boyacá), vereda Valero (R-Valero) (5°28'0,9"N 73°39'24"O) a 3.029 msnm. Ambas localidades se encontraban separadas 1,39 km.
La segunda zona se encuentra en un menor estado de conservación, ya que gran parte del bosque de roble ha sido talado para dedicar estas áreas a la ganadería. Se ubicó en el municipio de San Miguel de Sema (Boyacá). En ella, la primera localidad correspondió a la vereda Centro (SM-Centro) (5°31'16"N 73°42'0,9"O) a 2.679 msnm, en la que el agraz crece en medio de un cultivo de pino parcialmente talado. La segunda localidad se ubicó en la vereda Peñas Blancas (SM-Peña Blanca) (5°32'10"N 73°43'62"O) a 2.734 msnm, donde el agraz crece en matorrales cerca de relictos de bosque de roble y potreros. Ambas localidades se encontraban separadas 2,3 km.
Observaciones en campo. Los muestreos se realizaron en febrero y marzo de 2012, durante el primer periodo de floración del agraz. Se realizaron cuatro visitas en cada una de las localidades, distribuidas a lo largo del periodo de floración.
Visitantes florales. Para la identificación de los visitantes florales se ajustó el método propuesto por Vaissiere et al. (2011). En cada visita se seleccionaron tres plantas de agraz distribuidas aleatoriamente en el sitio de estudio. En cada planta se delimitó un área de 50 cm2 con el fin de restringir los muestreos de visitantes a dicha área. También se determinó el número de flores que había dentro del área delimitada. Cada planta se tomó como una repetición. Se realizaron observaciones de 15 minutos durante cada hora entre las 06:00 y 17:00 h, en las que se contó el número de individuos de cada especie que visitaba las flores en el área delimitada. Algunos individuos de cada especie se capturaron y se identificaron en el laboratorio. Se registraron datos sobre el comportamiento de forrajeo de los visitantes: recurso colectado, duración de visita por flor y número de flores visitadas por minuto. Se midió la temperatura y humedad relativa del aire cada hora por medio de un higro-termómetro ubicado a 1 m de altura cerca de las plantas de agraz y protegido de la radiación solar directa, con el fin de determinar si la frecuencia de visitas estaba relacionada con alguna de estas variables.
Transporte de polen. Para evaluar el transporte de polen por parte de los visitantes florales se ajustó la metodología usada por Howlett et al. (2011). En cada muestreo se capturaron algunos visitantes florales por medio de una red entomológica. Los insectos se inmovilizaron con frío, y se les tomó muestras de polen de contacto de la cabeza, tórax y patas por medio de gelatina glicerinada teñida con fucsina. En el caso de las abejas que tenían polen en las corbículas, las cargas se retiraron y se almacenaron en viales marcados. Después, los insectos se liberaron. Cada individuo se consideró como una repetición. Las muestras se llevaron al Laboratorio de Meli-sopalinología (Dpto. Biología) de la Universidad Nacional de Colombia sede Bogotá, donde se hizo el correspondiente montaje de las muestras de gelatina en láminas porta-objetos para su posterior observación al microscopio óptico. Se contaron 300 granos de polen en cada lámina; en los casos en los que hubo menos de este número, se contaron todos los granos presentes. En los conteos se diferenció entre las tétradas de agraz y otros tipos de polen, los cuales se clasificaron por morfotipos. Se determinó la frecuencia de aparición del polen de agraz en cada una de las muestras.
Análisis de datos. Se calculó el coeficiente de correlación no paramétrico de Spearman entre el número de visitas de A. mellifera y Bombus hortulanus Friese, 1904 (Hymenoptera: Apidae), así como entre las visitas de A. mellifera y B. rubi-cundus Smith, 1854 (Hymenoptera: Apidae), en las dos zonas de muestreo, utilizando un nivel de significancia de P < 0,05.
Para los Resultados de comportamiento (tiempo de visita, número de flores visitadas por minuto), polen de contacto e índices, sólo se tuvieron en cuenta los datos de A. mellifera, B. hortulanus y B. rubicundus debido a que para los demás visitantes se contó con un número muy bajo de repeticiones (1-4 repeticiones).
Se usó la prueba de Shapiro-Wilk para determinar la distribución normal y la de Bartlett para evaluar la homogeneidad de varianzas. Para los datos que no siguieron una distribución normal (variable respuesta: número de granos de polen de agraz en cada parte del cuerpo. Factor: especie de visitante) se usó la prueba no paramétrica de Kruskall-Wallis para identificar diferencias estadísticas, utilizando un nivel de significancia de P < 0,05. En caso de encontrarse diferencias significativas se empleó la prueba de Mann-Whitne y Wilcoxon para determinar cuáles tratamientos fueron diferentes.
Para los datos que siguieron una distribución normal (variable: tiempo de las visitas en las flores y número de flores visitadas por minuto. Factor: especie de visitante) se aplicó la prueba paramétrica de análisis de varianza (ANAVA) de una vía para identificar diferencias estadísticas, utilizando un nivel de significancia de P < 0,05. En caso de encontrarse diferencias estadísticamente significativas se utilizó la prueba Diferencia Mínima Significativa (DMS) para determinar cuál tratamiento fue diferente.
Se realizó una regresión múltiple entre las variables ambientales (temperatura y humedad relativa) y el número de visitas de A. mellifera y B. hortulanus en cada una de las localidades. Para todos los análisis se utilizó el software de libre distribución R 2.12 (R Development CoreTeam 2008).
Para los tres visitantes más frecuentes, se calculó el Índice de Probabilidad de Polinización (PPI, "Pollination Probabi-lity Index") propuesto por Ne'man et al. (1999), de acuerdo con la siguiente fórmula:
PPI = PCP x PBP
donde:
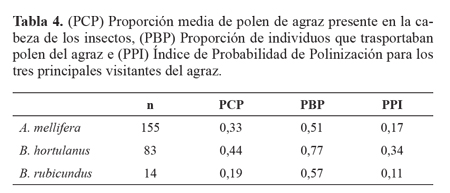
PCP ("Proportion of Conspecific Pollen") es la proporción media de polen de agraz presente en la cabeza de cada especie de insecto, debido a que es la única parte del cuerpo de los visitantes que entra en contacto con los órganos reproductivos de la flor. PBP ("Proportion of Bees carrying Pollen"), es la proporción de individuos de una especie que trasportan polen de agraz. El PPI toma valores entre 0 y 1, y refleja la probabilidad de que un visitante floral sea un polinizador eficiente de una planta en particular (Ne'man et al. 1999).
Se calculó el Índice de Importancia de Polinización (PII, "Pollination Importance Index") modificado por Escaravage y Wagner (2004), para los tres visitantes más frecuentes. Para calcular este índice inicialmente se obtuvo el PIV ("Pollina-tion Importance Value") de acuerdo con la siguiente fórmula:
PIV = PE x VR x C x PCC
donde:
PE o eficiencia básica de polinización (basic "Pollination Efficiency") se obtiene a partir de una escala arbitraria que evalúa el comportamiento de los visitantes florales; para este caso se utilizaron los siguientes parámetros: número de flores visitadas en un minuto (calificado entre 0 y 0,3), presencia/ ausencia en todos los puntos de muestreo (calificado entre 0 y 0,3) y si el visitante realiza o no vibración en las flores (calificado entre 0 y 0,4).VR o tasa de visitas ("Visitation Rate") es el porcentaje total de las visitas de cada especie con respecto al total de visitas registradas de todos los insectos para cada zona. C o constancia ("Constancy"), expresada como la proporción promedio de polen de agraz en la muestras de polen de la cabeza de cada especie, nótese que este valor es equivalente al valor PCP del índice PPI. pCc o capacidad de trasporte de polen ("Pollen Carrying Capacity"), es la proporción de polen de agraz trasportado por cada especie de visitante del total de polen de agraz llevado por todos los visitantes (sólo en la cabeza).
Finalmente, se calculó el PII para las tres especie de acuerdo con la siguiente fórmula:
PII = (PIV/APIV) x 100
ResultadosVisitantes florales. Se identificaron 11 especies de insectos visitantes del agraz, de las cuales seis fueron abejas (Apidae y Halictidae) (Tabla 1). De éstos, los más frecuentes en las cuatro localidades fueron A. mellifera y B. hortulanus (reinas, machos y obreras). Apis mellifera fue el más frecuente en todas las localidades y mostró su mayor frecuencia de visitas en G-R (84 ± 2%), mientras que B. hortulanus mostró su mayor frecuencia de visitas en la zona de SM (33 ± 6%). Las visitas de los demás insectos fueron poco frecuentes y sólo se encontraron en tres o menos localidades (Tabla 1).
Horarios de forrajeo. La temperatura y la humedad relativa estuvieron correlacionadas negativamente (Pearson, -0,80), siendo la temperatura la variable que más influyó sobre la actividad de A. mellifera, tanto en G-R (F = 24,15; df = 1, 1; P = 0,03) como en SM (F = 39,20; df = 1, 1; P = 4,8e-5). Para el caso de B. hortulanus, ni la temperatura ni la humedad relativa tuvieron efecto significativo (en SM F = 0,63; df = 1, 1; P = 0,41. En G-R F = 0,937; df = 1, 1; P = 0,33) sobre la actividad de este visitante en ninguna de las localidades (Fig. 1).
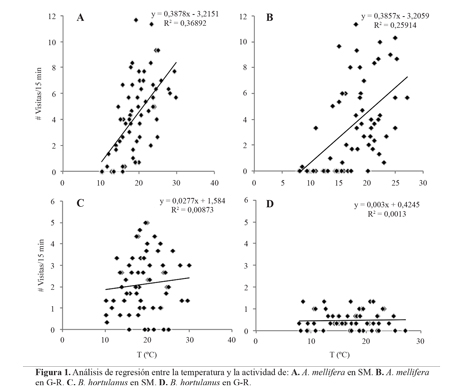
Las visitas de A. mellifera sobre el agraz inician entre las 07:00 y 08:00 h, muestran un pico de actividad cerca de las 12:00 h, y finalizan entre las 16:00 y 17:00 h (Figs. 2A-D). Bombus hortulanus inicia su actividad de forrajeo a las 06:00 h, y a diferencia de A. mellifera, no muestra picos de actividad marcados; sus visitas cesan después de las 17:00 h (Figs. 2A-D). El número de visitas de B. hortulanus no estuvo correlacionado con el número de visitas de A. mellifera en ninguna de las localidades (GR r = -0,066, P = 0,46; SM r = 0,173, P = 0,23). Bombus rubicundus no estuvo presente en SM-Peña Blanca (Fig. 2B), y en SM-Centro solo se observaron visitas entre las 06:00 y 08:00 h y entre las16:00 y 17:00 h (Fig. 2A); en este caso las visitas de B. rubicundus mostraron una correlación negativa con las visitas de A. mellifera (r = -0,390, P = 0,006). En G-R la actividad de B. rubicundus fue esporádica a lo largo de todo el día, entre las 06:00 y 17:00 h, aunque en R-Valero su actividad fue más constante en horas de la mañana (Fig. 2D), y no mostro correlación con las visitas de A. mellifera (r = -0,089, P = 0,32).
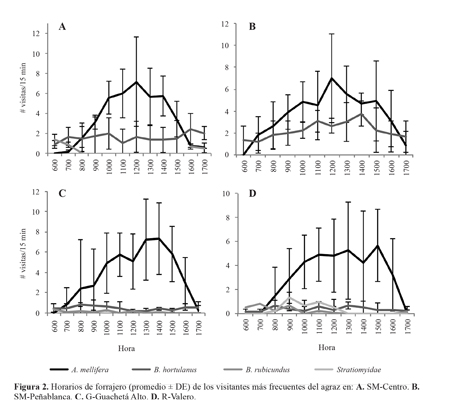
Tanto B. hortulanus como B. rubicundus pueden obtener néctar y polen de las flores de agraz. El polen lo obtienen gracias al comportamiento de vibración, que lo libera de las anteras poricidas. El tiempo de visita en las flores de B. hor-tulanus (1,9 ± 1,1 s) es significativamente mayor (F = 51,01; df = 2, 2; P = 0,037) que el de B. rubicundus (1,5 ± 0,7 s). Las dos especies visitan 15 ± 5 flores por minuto (F = 16,12; df = 2, 2; P = 0,65) (Tabla 2).
Transporte de polen. Solo de los tres principales visitantes (A. mellifera, B. hortulanus y B. rubicundus) se obtuvo un número de muestras suficientes, debido a que los demás fueron muy poco frecuentes, y en las escasas muestras colectadas se encontró muy poco polen.
En las muestras de polen de contacto se encontraron 42 tipos polínicos diferentes al agraz. Para A. mellifera se contabilizaron 38 morfotipos, para B. hortulanus 37, y para B. rubicundus 28; sin embargo, cada individuo transportó un máximo de 12 tipos polínicos. Apis mellifera y B. hortulanus comparten el 83% de los tipos polínicos que transportan en su cuerpo, mientras que A. mellifera y B. rubicundus comparten el 57%. Entre B. hortulanus y B. rubicundus comparten el 54% de los tipos polínicos.
B. hortulanus es el visitante que transporta un mayor número de granos de polen de agraz (tétradas) en la cabeza (F = 8,10; df = 2, 1; P = 0,00039). En el tórax y las patas de B. hortulanus y A. mellifera se observaron más granos de polen de agraz que para B. rubicundus (tórax F = 3,71; df = 2, 1; P = 0,026; patas F = 5,54; df = 2, 1; P = 0,0045), el cual transporta pocos granos de polen en su cuerpo (Tabla 3). Para A. mellifera y B. hortulanus, el porcentaje de polen de agraz en el cuerpo puede variar entre el 0 y 100%, mientras que para B. rubicundus varía entre 0 y 67%. Las muestras de cargas de polen corbicular que se obtuvieron muestran que A. mellifera lleva un mayor porcentaje de cargas monoflorales de agraz (Tabla 3).
Probabilidad de polinización del agraz por los insectos visitantes. Bombus hortulanus fue el visitante que transportó un mayor porcentaje de polen de agraz en la cabeza (PCP = 44%), así como un mayor porcentaje de individuos transportando polen de agraz (PBP = 77%), lo que hace que sea el visitante que mayor probabilidad tiene de ser un polinizador eficiente del agraz (PPI = 34%). Apis mellifera y B. rubicun-dus transportan un porcentaje de polen de agraz menor (PCP = 33 y 19% respectivamente), y el PBP fue similar para ambas especies (51 y 57% respectivamente), lo que hace que su valor PPI se reduzca (17 y 11% respectivamente) (Tabla 4).
El índice PII se calculó de forma independiente para las dos zonas de muestreo (Tabla 5). En ambas las especies de Bombus obtuvieron mayores valores de PE que A. mellifera debido, principalmente, a que presentaron el comportamiento de vibración en las flores. En las dos zonas el visitante más frecuente fue A. mellifera, seguido por B. hortulanus y B. rubicundus (Tabla 5).Tanto para la constancia (C) (equivalente al valor PCP del índice PPI) como para el valor PCC, B. hortulanus mostró los valores más altos, seguido por A. me-llifera y B. rubicundus. Finalmente, el PII muestra que en SM B. hortulanus realizó la mayor contribuyó a la polinización del agraz (69,5%), mientras que en G-R, A. mellifera tuvo la mayor contribución con el 64,1%. El aporte de B. rubicundus fue mínimo en ambas zonas (Tabla 5).
DiscusiónActividad de los principales visitantes. Los visitantes más frecuentes del agraz fueron A. mellifera, B. hortulanus y B. rubicundus, lo cual coincide con lo hallado en cultivos de arándanos de Estados Unidos, en los que A. mellifera y Bombus spp. son los visitantes más comunes (Ratti et al. 2008). También existen reportes sobre Syrphidae y Lepidoptera visitando Vac-cinium, pero al igual que en este estudio, son poco frecuentes y no transportan polen de Vaccinium (Davis et al. 2003).
La actividad constante de B. hortulanus desde la mañana hasta la tarde probablemente se deba a que los abejorros de Bombus tienen una mayor capacidad de termorregulación que A. mellifera, lo que les permite volar a temperaturas más bajas (Heinrich1980; 2004; Goulson 2010). Esto explicaría que la actividad de forrajeo de A. mellifera sobre el agraz comience más tarde, y que su pico de actividad sea cerca del mediodía, hora de mayor temperatura. Además, esto coincide con los Resultados del análisis de regresión, los cuales muestran que la actividad de A. mellifera está relacionada con la temperatura del aire en todas las localidades, mientras que la actividad de B. hortulanus no se relaciona con la temperatura en ninguno de los casos.
A pesar de que B. rubicundus también tiene un alta capacidad de termorregulación, mostró un patrón de actividad diferente, con visitas restringidas a horas de la mañana y de la tarde en SM, y visitas intermitentes en G-R. Esto puede deberse a que los abejorros del género Bombus presentan plasticidad en la selección de plantas para forrajeo, la cual puede ser afectada por la disponibilidad de los recursos y por la presencia de otras especies (Goulson 2010; Rubio 2011). El comportamiento de forrajeo de B. rubicundus en SM-Centro, correlacionado negativamente con el número de visitas de A. mellifera, podría considerarse como una estrategia para evitar la competencia con este visitante, más frecuente y con mayor actividad al mediodía.
Comportamiento de los principales visitantes. Apis me-llifera visita el agraz para obtener néctar, sin embargo, es posible encontrar polen de esta planta en baja cantidad en todo su cuerpo, además de cargas corbiculares monoflorales de agraz. Estos Resultados sugieren que A. mellifera puede adquirir indirectamente polen de la flor, que luego es almacenado paulatinamente en las corbículas. Este comportamiento también se ha observado en cultivos de V. macrocarpon (Phi-llips 2011).B. hortulanus y B. rubicundus pueden obtener activamente néctar y polen de las flores de agraz, este último gracias a su capacidad de vibración. El hecho de que las visitas de Bombus sean más cortas que las de A. mellifera también se ha observado en flores de V. agustifolium y V. corymbosum (Stubbs y Drummond1997), lo que les permite visitar un mayor número de flores por minuto (Dalyet al. 2013). Esto parece obedecer, en general, a que Bombus tiene la probóscide más larga que A. mellifera (Brodie 1996), lo que les puede facilitar adquirir los recursos de plantas que tiene corolas estrechas, como es el caso del agraz. Estas diferencias en el tiempo de visita también pueden estar determinadas por la morfología de la flor, ya que se ha observado que la tasa de visitas de A. mellifera es menor en flores de V. corymbosum con diámetro de la corola corto (Courcelles et al. 2013). Estas características de los abejorros Bombus los convierten en buenos polinizadores y, por ello, son apreciados en cultivos como el agraz (Batra 1997; Ratti et al. 2008).
En el agraz, cuyas flores son pequeñas y de corola acampanada y estrecha, la única parte del cuerpo de los visitantes que entra en contacto con el estigma y las anteras es la cabeza. Por esta razón, los datos de polen de contacto de la cabeza son de mayor importancia y, en este estudio, indican que B. hortulanus sería el visitante con más posibilidades de transferir polen de una flor a otra, hecho sustentado por el mayor índice PPI. El que B. rubicundus, que también vibra en las flores, lleve un número de granos de polen de agraz significativamente menor puede deberse a su patrón de visitas intermitentes y ocasionales sobre el agraz.
Aporte a la polinización del agraz por los principales visitantes. Los Resultados del PII indican que B. hortulanus contribuye con el mayor porcentaje de la polinización del agraz en SM, lo que muestra que su comportamiento especializado para la extracción de polen de este tipo de plantas compensa su menor número de visitas en comparación con A. mellife-ra. Sin embargo, en G-R, donde el número de visitas de B. hortulanus fue menor que en SM, el aporte a la polinización estimado para esta especie sobre el agraz se redujo notoriamente. Por lo tanto, no sólo es importante que un visitante sea buen polinizador en términos de una sola visita, sino que también debe brindar un número suficiente de visitas para que su aporte al servicio de polinización sea significativo. Varios estudios en zonas templadas demuestran que una población de tamaño adecuado de Bombus puede contribuir en gran medida con la polinización de los cultivos de Vaccinium, mejorando su nivel de polinización y producción (Cane et al. 1985; Mackenzie 1994; Brevis 2001; Ratti et al. 2008).
Por otro lado, en G-R, donde A. mellifera muestra su mayor frecuencia de visitas, el mayor porcentaje de la polinización estaría mediada por esta especie, lo que sugiere que su falta de especificidad al visitar las flores del agraz estaría compensada por la alta frecuencia de visitas. Esto coincide con lo observado en cultivos de arándanos, en los que el servicio de polinización de A. mellifera depende de la densidad de individuos, por lo que suelen ser los polinizadores más usados en cultivos donde la densidad de otras abejas es baja (Dedej y Delaplane 2003).
El agraz es una planta nativa de Suramérica (Ligarreto 2009), por lo que antes de la llegada de A. mellifera a América su polinización debió estar mediada principalmente por Bombus, quizá más abundante que en la actualidad, ya que tanto en Colombia como a nivel mundial las poblaciones de Bombus tienden a disminuir (Nates-Parra 2006; Goulson et al. 2008; Williams y Osborne 2009). En este estudio estimamos que Bombus brinda el mayor porcentaje de la polinización del agraz en SM y, por tanto, en la eventual exclusión de A. mellifera, la población de Bombus podría mantener un adecuado servicio de polinización para el agraz.
Desafortunadamente, las especies autóctonas no sólo están siendo afectadas por la modificación del paisaje por deforestación y expansión de la frontera agrícola (Goulson et al. 2008), sino también por la competencia con A. mellifera. Se ha visto que el solapamiento de nichos entre A. mellifera y Bombus puede llevar a reducciones tanto en la tasa de forrajeo de Bombus (Thomson 2006), la producción de sexuados en las colonias (Thomson 2004) y el tamaño de las obreras (Goulson y Sparrow 2009). La coincidencia en los tipos polínicos encontrados entre A. mellifera y B. hortulanus sugiere que ambas especies están visitando grupos muy similares de plantas, por lo que es probable que haya competencia entre ellas. La menor coincidencia entre los tipos polínicos transportados por A. mellifera y B. rubicundus, sumada al patrón de forrajeo de ésta y la correlación negativa entre las visitas de ambas especies podría indicar que B. rubicundus forrajea en plantas diferentes a las que visita A. mellifera para evitar la competencia. Si esto es así, el agraz estaría perdiendo el aporte importante de un polinizador potencial, pero se requieren estudios más detallados para comprobar esta hipótesis. Esto resalta la importancia de trabajar en la conservación de las especies nativas de Bombus, debido a que pueden estar contribuyendo en gran medida a la polinización del agraz, así como de otras plantas cultivadas y silvestres.
ConclusionesEste es el primer reporte sobre visitantes y polinizadores del agraz en Colombia, encontrando que en las zonas de estudio A. mellifera y B. hortulanus son los que más contribuyen a la polinización de esta planta, gracias a que transportan polen en su cabeza y son visitantes frecuentes. Bombus rubicundus tiene características que le permitirían ser un buen polini-zador, pero su baja frecuencia de visitas reduce su impacto sobre la polinización de esta planta. El identificar especies nativas de Bombus como polinizadores de un frutal promisorio resalta la importancia de trabajar en la conservación de éstas especies, así como su habitad natural para mantener un adecuado servicio de polinización.
AgradecimientosA los productores Misael Pachón, Carlos Moreno y Gloria Sierra. A los integrantes del Laboratorio de Investigación en Abejas de la Universidad Nacional (LABUN) por el apoyo durante los muestreos. Al departamento Administrativo de Ciencia, Tecnología e Innovación (COLCIENCIAS) y a la Universidad Nacional de Colombia por la financiación del proyecto "Valoración del servicio de Polinización en tres frutales promisorios para Colombia" (código 1101-521-28758). A los evaluadores anónimos por sus aportes.
Literatura citadaBATRA, S. 1997. Solitary bees for Vaccinium pollination. Acta Horticulturae 446 (1): 71-76. [ Links ]
BREVIS, P. A. 2001. Factors controlling fruit set of rabbiteye blue-berry (Vaccinium ashei Read). Tesis de doctorado, Universidad de Concepcion, Chile. 112 p. [ Links ]
BRODIE, L. 1996. Bumblebee foraging preference: differences bet-ween species and individuals. Tesis de pregrado.University of Aberdeen. Escocia. 36 p. [ Links ]
CANE, J. H.; EICKWORT, G. C.; WESLEY, R.; SPIELHOLZ, J. 1985. Pollination ecology of Vaccinium stamineum (Ericaceae: Vaccinioideae). American Journal of Botany 72 (1): 135-142. [ Links ]
CANE, J. H. 1997. Lifetime monetary value of individual pollinators: the bee Habropoda laboriosa at rabbiteye blueberry (Vaccinium ashei). Acta Horticulturae 446 (1): 67-70. [ Links ]
COURCELLES, D. M. M.; BUTTON, L.; ELLE, E. 2013. Bee visit rates vary with floral morphology among high bush blueberry cultivars (Vaccinium corymbosum L.). Journal of Applied Entomology 137 (9): 693-701. [ Links ]
DALY, K.; PACHECO, M.; POPLACK, A.; MAXON, M.; JOHNSON, C.; WHITCOMB, A.; KOPEC, K.; CYPEL B.; WARD L. 2013. Comparing Apis mellifera and Bombus spp. Pollination efficiencies on Willamette Valley blueberry farms. Oregon Undergraduate Research Journal 4 (1): 22-33. [ Links ]
DAVIS, A. N.; HOLLOWAY, P. S.; KRUSE, J. J. 2003. Insect visitors and potential pollinators of lingonberries, Vaccinium vitisidaea subsp. minus, in subarctic Alaska. Acta Horticulturae 626 (1): 433-438. [ Links ]
DEDEJ, D.; DELAPLANE, K. S. 2003. Honeybee (Hymenoptera: Apidae) pollination of rabbit eye blueberry Vaccinium ashei var. 'Climax' is pollinator density-dependent. Journal of Economic Entomology 96 (4): 1215-1220. [ Links ]
ESCARAVAGE, N.; WAGNER, J. 2004. Pollination effectiveness and pollen dispersal in a Rhododendron ferrugineum (Erica-ceae) population. Plant Biology 6 (5): 606-615. [ Links ]
GAVIRIA, C. A.; OCHOA, C. I.; SANCHEZ, N. Y; MEDINA, C. I.; LOBO, M.; GALEANO, P L.; MOSQUERA, A. J.; TAMACO, A.; LOPERA, Y E.; ROJANO, B. A. 2009. Propiedades antioxidantes de los frutos de agraz o mortiño (Vaccinium meridionale Swartz). pp. 93-109. En: Ligarreto, G. A. (Eds.). Perspectiva del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en zona altoandina de Colombia. Universidad Nacional de Colombia. Facultad de Agronomía. Bogotá. Colombia. 134 p. [ Links ]
GOULSON, D.; LYE, G. C.; DARVILL, B. 2008. Decline and conservation of bumblebees. Annual Review of Entomology 53 (1): 191-208. [ Links ]
GOULSON, D.; SPARROW, K. R. 2009. Evidence for competition between honey bees and bumblebees; effects on bumblebee worker size. Journal of Insect Conservation 13 (1): 177-181. [ Links ]
GOULSON, D. 2010. Bumblebees, behavior, ecology and conservation. Oxford University Press, 2a edición. Nueva York. EEUU. 300 p. [ Links ]
GRIFFIN, J. J.; BLAZICH, F. A. 2008. Vaccinium L. pp. 1154-1159. En: Bonner, F. T.; Karrfalt, R. P. (Eds.). Woody plant seed manual. U.S. Department Agricultural and Forest Service. Washington. EE.UU. 12-23 p. [ Links ]
HEINRICH, B. 1980. Mechanisms of body-temperature regulation in honeybees, Apis mellifera. Journal of Experimental Biology 85 (1): 61-72. [ Links ]
HEINRICH, B. 2004. Bumblebee economics. Harvard University. Cambridge, EEUU. 245 p. [ Links ]
HOWLETT, B. G.; WALKER, M. K.; RADER, R.; BUTLER, R. C.; NEWSTROM-LLOYD I.; TEULON, D. A. J. 2011. Can insect body pollen counts be used to estimate pollen deposition on pakchoi stigmas. New Zealand Plant Protection 64 (1): 25-31. [ Links ]
JANICK, J.; PAULL, R. E. 2008. The encyclopedia of fruit and nuts. CAB International. Oxfordshire. Reino Unido. 954 p. [ Links ]
JAVOREK, S. K.; MACKENZIE, K. E.; VANDER, S. P. 2002. Comparative pollination effectiveness among bees (Hymenop-tera: Apoidea) on Low bush blueberry (Ericaceae: Vaccinium angustifolium). Annals of the Entomological Society of America 95 (1): 345-351. [ Links ]
JUDD, W. S.; CAMPBELL, C. S.; KELLOGG, E. A.; STEVENS, P. F.; DONOGHUE, M. J. 2007. Plant systematics: A phylogenetic approach, Sunderland, 3a edición. EE.UU. 490 p. [ Links ]
LIGARRETO, G. A. 2009. Descripción del género Vaccinium, estudio del caso: agraz o mortiño (Vaccinium meridionale Swartz). pp. 13-27. En: Ligarreto, G. A. (Eds.). Perspectiva del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en zona altoandina de Colombia. Universidad Nacional de Colombia. Facultad de Agronomía. Bogotá. Colombia. 134 p. [ Links ]
MACKENZIE, K. E. 1994. The foraging behaviour of honeybees (Apis mellifera) and bumblebees (Bombus spp.) on cranberry. Apidologie 25 (1): 375-383. [ Links ]
MUÑOZ, J. D.; MARTÍNEZ, L. F.; LIGARRETO, G. A. 2009. Caracterización de los ambientes agroecológicos del agraz o mortiño (Vaccinium meridionale Swartz) en zonas altoandinas de Colombia. pp. 29-55. En: Ligarreto, G. A. (Eds.). Perspectiva del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en zona altoandina de Colombia. Universidad Nacional de Colombia. Facultad de Agronomía. Bogotá, Colombia. 134 p. [ Links ]
NATES-PARRA, G. 2006. Abejas corbiculadas de Colombia: Hymenoptera: Apidae. Universidad Nacional de Colombia. Facultad de Ciencias. Bogotá, Colombia. 155 p. [ Links ]
NE'MAN, G., G.; DAFNI, A.; POTTS, S. G. 1999. A new pollination probability index (PPI) for pollen load analysis as a measure for pollination effectiveness of bees. Journal of Apicultural Research 38 1): 19-23. [ Links ]
PHILLIPS, K. 2011. A comparison of bumblebees (Bombus spp.) and honey bees (Apis mellifera) for the pollination of Ore-gon cranberries (Ericaceae: Vaccinium macrocarpon). Tesis de Maestria. Oregon State University. Oregon, EEUU. 99 p. [ Links ]
RATTI, C. M. 2006. Bee abundance and diversity in berry agriculture. Tesis de Maestría, Simon Fraser University, British Colum-bia, Canadá. 87 p. [ Links ]
RATTI, C. M.; HIGO, H. A.; GRISWOLD, T.; WINSTON, M. L. 2008. Bumblebees influence berry size in commercial Vaccinium spp. cultivation in British Columbia. The Canadian Entomologist 140 (3): 348-363. [ Links ]
RUBIO D. 2011. Disponibilidad, uso y preferencia por los recursos florales en una comunidad de abejorros (Hymenoptera: Apidae: Bombus) en el páramo de Chingaza. Tesis de maestría. Universidad Nacional de Colombia. Bogotá, Colombia. 92 p. [ Links ]
STUBBS, C.; DRUMMOND, F. A. 1997. Blueberry and cranberry (Vaccinium spp.) pollination: a comparison of managed and native bee foraging behavior. Acta Horticulturae 437 (1): 341-344. [ Links ]
THOMSON, D. 2004.Competitive interactions between the invasive European honeybee and native bumblebees. Ecology 85 (2): 458-470. [ Links ]
THOMSON, D. 2006. Detecting the effects of introduced species: a case study of competition between Apis and Bombus. Oikos 114 (3): 407-418. [ Links ]
TORRES, W. S.; MONTOYA, I. A.; LIGARRETO, G. A. 2009. Aspectos sociales y económicos de la producción de agraz o mortiño (Vaccinium meridionale Swartz). pp. 113-134. En: Ligarreto, G. A. (Eds.). Perspectiva del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en zona altoandina de Colombia. Universidad Nacional de Colombia. Facultad de Agronomía. Bogotá, Colombia. 134 p. [ Links ]
VAISSIERE, B.; FREITAS, B. M.; GEMMILL-HERREN, B. 2011. Protocol to detect and assess pollination deficits in crops: a handbook for its use. Food and Agriculture Organization of the United Nations (FAO). Roma, Italia. 72 p. [ Links ]
WILLIAMS, P.; OSBORNE, J. L. 2009. Bumblebee vulnerability and conservation worldwide. Apidologie 40 (3): 367-387. [ Links ]
Recibido: 5-mar-2014
Aceptado: 30-mar-2015
Citación sugerida:
PINILLA-GALLEGO, M. S.; NATES-PARRA, G. 2015. Visitantes florales y polinizadores en poblaciones silvestres de agraz (Vaccinium meridionale) del bosque andino colombiano. Revista Colombiana de Entomología 41 (1): 112-119. Enero-Junio 2015. ISSN 0120-0488.

