










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.41 no.1 Bogotá Jan./June 2015
Fluctuación poblacional de arañas (Araneae: Tetragnathidae, Pholcidae) en el agroecosistema cacao en Tabasco, México
Population fluctuation of spiders (Araneae: Tetragnathidae, Pholcidae) in cocoa agroecosystem Tabasco, México
ARACELY DE LA CRUZ-PÉREZ,12, MANUEL PÉREZ-DE LA CRUZ1,3, SAÚL SÁNCHEZ-SOTO5 y MAGDIEL TORRES-DE LA CRUZ1,4
1 Profesor Investigador, Ph. D. División académica de Ciencias Biológicas. Universidad Juárez Autónoma de Tabasco, Carretera Villahermosa-Cárdenas Km
0.5. C. P. 86039. Villahermosa, Tabasco, México.
2 arace_lycp@hotmail.com.
3 perezmandoc@hotmail.com.
4 magtorre@colpos.mx. Autor para correspondencia.
5 Dr. Colegio de Postgraduados, Campus Tabasco, Km. 3.5 Carretera Cárdenas-Huimanguillo. Apartado postal 24, 86500, H. Cárdenas, Tabasco, México.
Resumen: La abundancia de Leucauge argyra y Pholcus sp. fue estudiada en diferentes microhábitats del agroecosistema cacao en el sureste de México. Cuatro microhábitats verticales se delimitaron: "drenes", "zona baja", "zona media" y "dosel". Así también, tres épocas del año fueron consideradas: nortes, seca y lluvias. Leucauge argyra fue dominante en la época de lluvias en los cuatro microhábitats y en los drenes durante la época de nortes (D = 0,71). Pholcus sp. fue dominante en la zona baja (D = 0,32) y zona media (D = 0,22) durante la época de nortes. Además, la presencia de Pholcus sp. fue notable durante la época de seca en la zona baja (D = 0,23) y, en menor cantidad, en el dosel del árbol de cacao. La estructura del agroecosistema, el manejo del cultivo, la migración de las arañas y la temperatura y la humedad relativa en el año son factores que influyen sobre la abundancia de las especies dominantes entre los estratos estudiados. La fluctuación de las arañas refleja cambios en la abundancia de ambas especies durante las épocas del año dentro del cultivo cacao. Hasta nuestro conocimiento, este trabajo es uno de los primeros estudios donde se reportan las preferencias de establecimiento de L. argyra y Pholcus sp. dentro de los microhábitats en el agroecosistema cacao en México.
Palabras clave: Abundancia. Cacao. Dominancia.
Abstract: The abundance of Leucauge argyra y Pholcus sp. was studied in different microhabitats of cocoa agroecosystem in the southeast of Mexico. Four vertical microhabitats were defined: "drains", "lower area", "Middle area" and "Canopy". In addition three times of the year were considered: windy, dry and rainy. Leucauge argyra was dominant in the rainy season in the four microhabitats, and in the drains during the windy season (D = 0.71). Pholcus sp. was dominant in the lower area (D = 0.32) and middle area (D = 0.22) during the windy season. In addition, the presence of Pholcus sp. was noticeable during the dry season in the lower area and fewer in the cocoa tree canopy. The agroecosystem stracture, crop management, migration of spiders, and the variation of the temperature and relative humidity in the year are factors that influence the abundance of dominant species between the layers studied. The fluctuation of spiders reflects changes in the abundance of both species during the seasons in the cocoa crop. To our knowledge, this work is one of thefirst studies reporting the preferences of establishment of L. argyra and Pholcus sp. within the microhabitats in the cocoa agroecosystem in México.
Key words: Abundance. Cocoa. Dominance.
Introducción
Las arañas son consideradas un grupo mega diverso, se distribuyen en casi todos los hábitats terrestres (Levi y Codding-ton 1991); son abundantes entre los estratos de la vegetación donde ocupan nichos espacios-temporales disponibles, alimentándose de una diversidad de insectos y formando parte del complejo de enemigos naturales en casi todos los ecosistemas terrestres (Turnbull 1973; Foelix 1996; Perez-De la Cruz et al. 2007). La presencia de arañas en los agroecosiste-mas ha sido documentada en cultivos de alfalfa, sorgo, algodón y trigo en los cuales la mayoría de las familias reportadas son tejedoras (Nyffeler 1999; Nyffeler y Sunderland 2003). En México se ha explorado la diversidad de arañas en los agrecosistemas de café (Ibarra-Núñez 1990; Ibarra-Núñez et al. 1995; Ibarra-Núñez y García 1998), cacao (Ibarra-Núñez et al. 2004; Pérez-De La Cruz et al. 2007; De La Cruz-Pérez et al. 2009), sistemas agroforestales (Burgos 2013), humedales (Llinas-Gutiérrez y Jiménez 2004) y zonas áridas (Correa 2004). La presencia de especies de la familia Tetragnathidae es notable es estos sistemas; tan sólo las especies Leucauge argyra (Walckenaer, 1841) (Tetragnathidae) y Pholcus sp. (Pholcidae) han sido reportadas como abundantes dentro del agroecosistema cacao, el cual se caracteriza por tener una estructura similar a los estratos vegetales típicos de un ecosistema natural (De La Cruz-Pérez et al. 2009). La estructura de este cultivo contribuye a microambientes menos fluctuantes, favoreciendo el establecimiento de artrópodos que interactúan entre estos sitios (López et al. 2000). En este agroecosistema se han registrado cerca de 1.500 especies de artrópodos (Entwistle 1982) en donde las arañas desempeñan la función de regular las poblaciones de artrópodos, principalmente de insectos al ser depredadoras (Riechert y Bishoz 1990; Wise 1993; Sunderland 1999). Esto ha despertado el interés de investigar el comportamiento de sus poblaciones dentro de los agroecosistemas. Así también, conocer la respuesta de las poblaciones de arañas a los cambios de vegetación, dentro de los cultivos perennes donde las actividades antropógenicas varían en función al tipo de cultivo, a las estaciones del año y a la zona geográfica. Además, conocer si las condiciones microclimáticas afectan el desplazamiento de las especies de arañas entre los estratos de la vegetación dentro de los cultivos perennes. Por tal motivo, el objetivo del presente trabajo fue conocer la fluctuación de L. argyra y Pholcus sp. en cuatro microhábitat durante las épocas del año y correlacionar la temperatura y humedad con la abundancia de ambas especies.
Materiales y métodosÁrea de estudio. El estudio se realizó durante 2004-2005 en una plantación de cacao de 6 ha con 35 años de edad, sembrada en un marco real de 4 x 4 m. La plantación se localiza en las coordenadas 17°38'5.1"N 92°55'48.2"W a 10 msnm, en el Ejido "Las Delicias" de la Ranchería José María Morelos y Pavón, Teapa, Tabasco, México. La fisiografía de la zona es de valle abierto con un clima cálido húmedo con lluvias todo el año (Af), la temperatura promedio es de 25 °C. La precipitación alcanza una media anual de 3.933 mm (Anónimo 2000).
Las plantaciones de cacao en Tabasco requieren de dos tipos de sombra: la temporal, que se utiliza para proteger cuando el árbol es pequeño, y la permanente en la fase de establecimiento y edad productiva del cultivo. La presencia de leguminosas, frutales y maderables proveen sombra al cultivo. Las especies de árboles de sombra típicas dentro de una plantación de cacao son: Cedrella odorata L., Swietenia macrophylla King., Persea americana Mill., Tabebuia rosea (Bertol.) Dc., Bursera simaruba (L.) Sarg., Manilkara zapo-ta (L.) Van Royen, Artocarpus altilis Fosberg, Mangifera indica L., Citrus sinensis (L.) Osbeck, Colubrina arborescens (Mill.) Sarg., Gliricida sepium (Jacq.) Kunth ex Walp., Gua-zuma ulmifolia Lam. y Musa spp. Las aráceas, piperáceas y helechos se establecen en los claros dentro de la plantación y en la orilla de los drenes. Los drenes varían en profundidad y longitud facilitando el desalojo del agua dentro de la plantación durante la época de lluvia y se elaboran en función al declive del terreno, la profundidad y longitud varían dependiendo los requerimientos del terreno (López et al. 2000; De La Cruz-Pérez et al. 2009).
Caracterización de los microhábitat en el área de estudioUna visita previa fue necesaria para caracterizar los microhá-bitat dentro del agroecosistema cacao y conocer las especies vegetales presentes en el área de estudio. Las colectas botánicas se realizaron dentro del cultivo y en las orillas de los drenes. El material vegetal fue colectado con prensas botánicas y se colocaron en la secadora durante tres días, posteriormente, fueron identificadas por comparación de ejemplares en la Colección botánica del Colegio Superior de Agricultura Tropical (CSAT) actualmente Colegio de Postgraduados (CP). Las especies de árboles de sombra fueron identificadas con el manual de Pennington y Sarukhán (1998). En el Anexo I, se listan las especies vegetales presentes en cada microhábitat en el agroecosistema cacao en estudio.
Método de muestreo y colecta. Las colectas de arañas fueron realizadas en 1 ha de cacao por tres personas cada 15 días, dedicando cuatro horas-hombre (07:00-11:00). Los números de cuadrantes se determinaron por un muestreo piloto previo, realizado al azar en el cual se determinaron 24 cuadrantes con una superficie de 16 m2 cada uno. En cada cuadrante se delimitaron cuatro microhábitats utilizando la clasificación propuesta por Duffey (1966) con modificaciones al tipo de agroecosistema, considerando la caracterización vegetal del área de estudio (Anexo I). Se les denominó "drenes" (DN): a las zanjas localizadas dentro del agroecosistema, estos varían en profundidad y longitud. La "zona baja" (ZB): consistió en la hojarasca, piedras y vegetación no mayor a 15 cm de altura. La "zona media" (ZM): se determinó por la vegetación mayor a 15 cm y menor a 150 cm de altura. El "dosel" (DL): constituido por la copa de los árboles de cacao (se colectó en el dosel bajo a una altura de 1,50 a 1,80 m).
Las colectas se realizaron en cada cuadrante mediante la búsqueda intensiva de las arañas (Gaviño et al. 1979). Para la captura se utilizaron aspiradores bucales y pinzas entomológicas. Los ejemplares fueron separados por microhábitat y colocados en frascos de plástico con alcohol etílico al 70% para su conservación con los siguientes datos: número de colecta, fecha, número de cuadrante, microhábitat, lugar y recolector. El material identificado fue depositado en la colección entomológica del Colegio Superior de Agricultura Tropical (CSAT) actualmente Colegio de Postgraduados. Los nombres de las especies fueron cotejados en el The World Spider Cata-log (Platnick 2014).
Medición de temperatura y humedad ambiental. La temperatura y la humedad relativa fue medida en cada fecha de muestreo con un termohigrómetro digital modelo C 9120. Las lecturas en cada microhábitat se realizaron durante 5 h, entre las 06:00 y las 11:00 horas.
Análisis de los datos. La especie dominante en cada micro-hábitat fue identificada utilizando la abundancia de ambas especies en cada microhábitat por época del año, utilizando el índice Berger-Parker (Moreno 2001). Las gráficas de fluctuación se obtuvieron de la abundancia mensual de las especies y los promedios de temperatura y humedad en cada microhábitat.
Análisis de las variables ambientales. Los valores de temperatura y humedad se utilizaron para obtener la media, desviación estándar y rango en cada microhábitat con la finalidad identificar cambios mínimos en los valores de las variables estudiadas. Se aplico el índice de correlación de Pearson: r =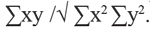 . Donde: x = variable independiente (temperatura y humedad) y = variable dependiente (abundancia y riqueza), empleando los promedios mensuales de la temperatura y humedad utilizando el programa Statistix (1996). Este índice considera que existe una relación directamente proporcional cuando se obtienen valores positivos y cuando los valores son negativos indica una relación inversamente proporcional entre las variables (Zar 1974). La prueba de t<p< 0.05) fue aplicada entre las variables estudiadas para conocer si existía una relación significativa entre ellas.
. Donde: x = variable independiente (temperatura y humedad) y = variable dependiente (abundancia y riqueza), empleando los promedios mensuales de la temperatura y humedad utilizando el programa Statistix (1996). Este índice considera que existe una relación directamente proporcional cuando se obtienen valores positivos y cuando los valores son negativos indica una relación inversamente proporcional entre las variables (Zar 1974). La prueba de t<p< 0.05) fue aplicada entre las variables estudiadas para conocer si existía una relación significativa entre ellas.
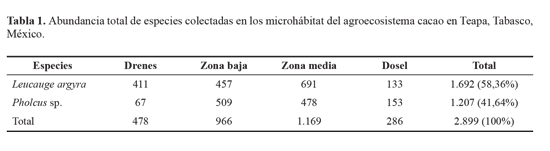
En este estudio contabilizamos 2.899 arañas adultas de las cuales L. argyra aportó el 58,36% del total de la muestra y Pholcus sp. el 41,64%. Leucauge argyra se colectó todo el año en sitios cercanos a los cuerpos de agua (DN) donde la presencia de Pholcus sp. fue menor. Sin embargo, ambas especies interactuaron en la zona baja y la zona media pero en menor cantidad en el dosel (Tabla 1).
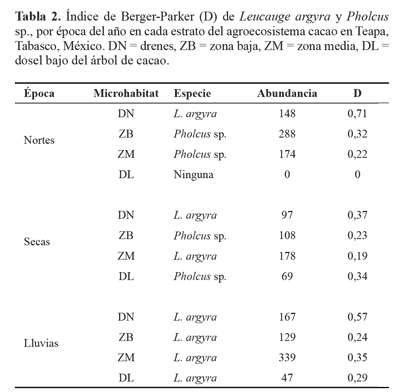
El índice de Berger-Parker indicó que L. argyra es dominante en la época de lluvia en los cuatro microhábitats y en los drenes durante la época de nortes (D = 0,71). Por otra parte, Pholcus sp. fue dominante en los microhábitats con mayor presencia de herbáceas (Anexo 1); esta especie se encontró en la zona baja (D = 0,32) y zona media (D = 0,22) durante la época de nortes. Así también, su presencia fue notable durante la época de seca en la zona baja (D = 0,23) y en menor cantidad en el dosel del árbol de cacao (Tabla 2).
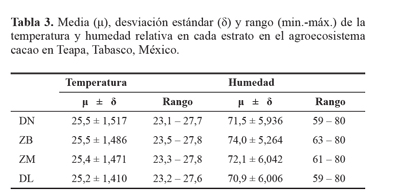
Los promedios de temperatura y humedad, incluyendo las desviaciones estándares, registraron variaciones en decimas y unidades respectivamente. Aunque las diferencias de temperaturas son mínimas, tienen influencia sobre la abundancia de las especies de arañas, pero los ligeros cambios de humedad en los microhábitat (Tabla 3) tienen relación con la abundancia de ambas especies en los distintos microhábitats, en base a la fluctuación de las arañas durante el estudio (Fig. 1).
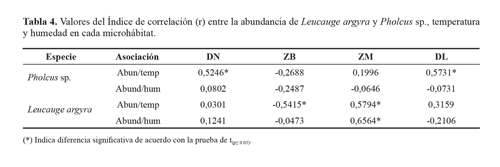
El índice de correlación entre las variables ambientales y la abundancia de las arañas mostraron variaciones en los microhábitats, observándose diferencias significativas al aplicar la prueba de t(P< 0.05) entre la abundancia-temperatura en drenes y dosel para Pholcus sp., a diferencia de L. argyra que registro asociaciones positiva y negativa con la abundancia-temperatura en la zona baja y dosel y una asociación positiva fuerte con la abundancia/humedad en la zona media (r = 0,6564*) (Tabla 4).
La fluctuación de ambas especies dominantes en los cuatro microhábitat indico que, en los drenes, la abundancia de L. argyra fue mayor que la de Pholcus sp. registrándose la mayor densidad en las épocas de lluvias (julio-octubre) y nortes (noviembre-febrero). En la zona baja, ambas especies registraron un comportamiento similar, observándose la máxima abundancia durante los meses que corresponden a la época de nortes (noviembre-febrero). En la zona media Pholcus sp. fue más abundante que L. argyra durante la época de lluvias (julio-octubre), en la cual presentó la mayor población, mientras que la abundancia de L. argyra fue poco variada. En el dosel ambas especies registraron un comportamiento similar con variaciones a lo largo del año, donde cada una presentó cuatro picos poblacionales (Fig. 1).
DiscusiónLos Resultados muestran que L. argyra y Pholcus sp. responden a los cambios en la diversidad y complejidad vegetal y a los gradientes de altitud en el agroecosistema cacao. Estas especies se presentan constantemente de un estrato vegetal a otro dentro del cultivo. La presencia y dominancia de L. argyra y Pholcus sp. durante la mayor parte del año, en los distintos microhábitats estudiados, muestran que ambas especies responden a los cambios en la estructura del hábitat. Esto es diferente a lo que ocurre en los monocultivos donde la baja diversidad y complejidad vegetal tienen efecto sobre la presencia y abundancia de las especies de arañas tejedoras (Duffey 1978; Halaj et al. 1998). La dominancia de algunas familias y especies de arañas en monocultivos y en agroeco-sistemas complejos varía según la región, manejo y tipo de cultivo, donde la mayor diversidad de arañas se presenta en agroecosistemas complejos. Las familias Araneidae, Tetrag-nathidae y Tomisidae son abundantes en cultivos de avena, calabaza, chile, col, frijol y sorgo en Durango y Zacatecas, México (Suárez-Forero et al. 2012).
A nuestro conocimiento, en este estudio se reporta por primera vez la preferencia de microhábitats de L. argyra y Pholcus sp. dentro del cultivo cacao en México. De acuerdo con Suárez-Forero et al. (2012), la preferencia de microhábitats de algunas especies de arañas se encuentra determinada por los sitios para el establecimiento de sus redes. Así, la presencia de plantas con determinada estructura para soporte de las telas tiene importancia en la distribución de algunas especies de arañas, por ejemplo Pérez-De La Cruz y De La Cruz-Pé-rez (2005) mencionan la asociación de Micrathena sagittata (Walckenaer, 1841) (Araneidae) con la planta Philodendron sagittifolium Liebm. (Araceae) y reportan la preferencia de Theridion evexum (Keyserling, 1884) (Theridiidae) al follaje del árbol de cacao. Así también, Tidarren mixtum (O. P Cambridge, 1896) (Theridiidae) es abundante en el tronco de cocohite Gliricida sepium (Jacq. Kunth ex Walp, 1842) (Fa-baceae) en pastizales (Pérez-De La Cruz y De La Cruz-Pérez, 2005). Por otra parte, consideramos que la distribución de las especies de arañas entre los microhábitats se encuentra relacionada con la preferencia de alimento. Al respecto Pérez-De La Cruz et al. (2007) reportaron para el agroecosistema cacao seis especies de Tetragnatidae, entre ellas L. argyra, las cuales muestran preferencia por alimentarse de Culicidae, Formicidae, Termitidae, Delphacidae, Thripidae, Gryllidae y Nymphalidae, a diferencia de las de Pholcidae (Pholcus sp. 1 y Pholcus sp. 2) que se alimentan de Culicidae, Formicidae, Termitidae y Aphidiidae.
En este estudio se considera que la estructura del agro-ecosistema cacao, las actividades del manejo del cultivo (poda, eliminación de renuevos y raleo), las variables climáticas a lo largo del año y la migración de las arañas entre los estratos de la vegetación son factores que influyen sobre la abundancia de las especies dominantes entre los estratos estudiados, al observarse correlaciones positivas y negativas. La fluctuación de las arañas refleja el efecto de los factores antes mencionados sobre la abundancia de ambas especies. Estos Resultados son similares a los reportados por Gertsch y Riechert (1976), Lubin (1978), Abraham (1983) y Ayazo y Soto (2007), quienes reportan que las especies de arañas varían de un estrato a otro y que las variables ambientales influyen en la selección de los microhábitats por las arañas. La complejidad del agroecosistema cacao favorece la presencia o ausencia de ambas especies de arañas entre los estratos de la vegetación al proveer sitios disponibles para el anclaje de las redes, disponibilidad de áreas de refugio, sitios de reproducción y disponibilidad de presas. Esto también se ha documentado en los trabajos de Lubin (1978), Dobel et al. (1990), Uetz y Hodge (1990), Rypstra et al. (1999) y Samu et al. 1999. Además de lo mencionado por estos autores, consideramos que la humedad y la temperatura son determinantes. En los sistemas forestales de monocultivo y policul-tivo en Yucatán, México se ha registrado que la comunidad de arañas puede aumentar o disminuir con los períodos de lluvia (Burgos 2013); por otra parte, Flórez (1997) menciona que factores como la temperatura y la humedad están relacionados con la abundancia de las poblaciones de arácnidos. Además se ha reportado que la fenología de la artropodofau-na tropical demuestra cambios en las densidades poblaciona-les relacionadas con las variaciones interanuales de lluvia y sequía.
Conclusión y recomendacionesLos Resultados de la presente investigación son relevantes para el conocimiento de la biología de las arañas en el agroecosistema cacao en México, donde se reportan las preferencias de establecimiento de las arañas de L. argyra (Tetragnathidae) y Pholcus sp. (Pholcidae). Ambas especies interactúan al desplazarse de un estrato vegetal a otro e incluso coexisten en un mismo microhábitat. Consideramos la importancia de repetir el estudio durante un tiempo más prolongado, donde se tomen en cuenta las actividades de manejo del cultivo; así también se sugiere colocar un instrumento que permita medir de forma constante los cambios mínimos de temperatura y humedad en los microhábitat dentro de la plantación. Los Resultados mostrados en el presente documento no deben ser generalizados para el cultivo cacao, pues la finalidad y el manejo que se le da al cultivo en diversas zonas en México (orgánicos, convencionales o subsistencia) modifica la estructura vegetal presente en este cultivo incluso en zonas dentro de una misma región y con esto la presencia y abundancia de las especies de arañas puede variar. En trabajos futuros es importante evaluar las diferencias entre las actividades en el manejo del agroeco-sistema cacao convencional y tradicional que modifican la estructura del hábitat de las arañas. Esto ayudaría a entender cómo las arañas se adaptan o desplazan en ausencia o disponibilidad de un recurso dentro del agroecosistema, e incluso conocer especies de arañas con asociaciones hacia una especie de planta en particular.
AgradecimientosAgradecemos a la familia Alvarado y al dueño de la parcela de cacao por las facilidades para realizar el estudio. Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por financiar la investigación.
Literatura citadaABRAHAM, B. J. 1983. Spatial and temporal patterns in a sagebrush steppe spider community (Arachnida: Araneae). Journal Arachnology 11: 31-50. [ Links ]
ANALYTICAL SOFTWAER. 1996. Statistix. Version 1.0. [ Links ]
ANÓNIMO. 2000. Cuaderno estadístico municipal. Teapa, Tabasco. Instituto Nacional de Estadística Geografía e Informática, INEGI. Villahermosa, Tabasco. 77 p. [ Links ]
AYAZO, B. R.; SOTO, M. R. 2007. Influencia en la estructura vegetal en la comunidad de arañas (Arachnida: Araneae) del suelo en un sistema silvopastoril de Córdoba, Colombia. 113 p. [ Links ]
BURGOS, S. J. Y. 2013. Composición y abundancia de artrópodos en monocultivos y policultivos de Swietenia macrophilla en el sur de Yucatán, México. Informe de residencia profesional. Universidad Autónoma de Yucatán. Campus de Ciencias Biológicas y Agropecuarias. Mérida, Yucatán. 32 p. [ Links ]
CORREA, R. M. M. 2004. Estudio comparativo de las familias Anyphaenidae, Araneidae, Mimetidae, Tetragnathidae y Theridii-dae (Arachnida: Araneae) en los humedales San Isidro y San José Comondú, Baja California Sur, México. Tesis de posgrado. Centro de Investigaciones Biológicas del Noroeste, S.C. La Paz, Baja California Sur. 103 p. [ Links ]
DE LA CRUZ-PÉREZ, A.; SÁNCHEZ-SOTO, S.; ORTÍZ-GARCÍA, C. F.; DE LA CRUZ- PÉREZ, M.; ZAPATA-MATA, R. 2009. Boletín del Museo de Entomología de la Universidad del Valle 10 (2): 1-9. [ Links ]
DOBEL, H. G.; DENNO, R. F.; CODDINGTON, J. A. 1990. Spider (Araneae) community structure in an intertidal salt marsh: Effects of vegetation structure and tidal flooding. Environmental Entomology 5(19): 1356-1370. [ Links ]
DUFFEY, E. 1966. Spider ecology and habitat structure (Arachnida: Araneae). Senck Biology 47: 45. [ Links ]
DUFFEY, E. 1978. Ecological strategies in spiders including some characteristics of species in pioneer y nature habitats. Symposium zoology. Society London 42: 109-123. [ Links ]
ENTWISTLE, P. F. 1982. Los insectos y el cacao. pp. 209-251. En: Wood, G. A. R. (Ed.). Cacao. CECSA. México. 363 p. [ Links ]
FOELIX, F. R. 1996. Biology of spiders. Oxford. New York. 330 p. [ Links ]
FLÓREZ, E. 1997. Estudio de la comunidad de arañas del bosque tropical de la estación biológica "El Vínculo". Cespedesia. 22 (69): 37-57. [ Links ]
GAVIÑO, D. L. T. G.; JUÁREZ, L. C.; FIGUEROA, T. H. H. 1979. Técnicas selectas de laboratorio y de campo. Limusa-Willey S.A. de C.V. México. 297 p. [ Links ]
GERTSCH, W. J.; RIECHERT, S. E. 1976. The spatial and temporal partitioning of a desert spider community, with descriptions of new species. American Museum Novitates 2604: 1-25. [ Links ]
HALAJ, J. D.; ROSS, W.; MOLDENKE, A. R. 1998. Habitat structure and prey availability as predictors or the abundance and community organization of spider in western Oregon forest canopies. Journal of Arachnology 26: 203-220. [ Links ]
IBARRA-NÚÑEZ, G. 1990. Los artrópodos asociados a cafetos en un cafetal mixto del Soconusco, Chiapas, México. I Variedad y abundancia. Folia Entomológica Mexicana 79: 207-231. [ Links ]
IBARRA-NÚÑEZ, G.; GARCÍA, J. A. 1998. Diversidad de tres familias de arañas tejedoras (Araneae: Araneidae, Tetragnathidae, Theridiidae) en cafetales del Soconusco, Chiapas, México. Folia Entomológica Mexicana 102:11-20. [ Links ]
IBARRA-NÚÑEZ, G.; GARCÍA, J. A.; MORENO, M. 1995. La comunidad de artrópodos de dos cafetales con diferentes prácticas agrícolas (orgánico y convencional): el caso de las arañas. Memorias XXX Congreso Nacional de la Sociedad Mexicana de Entomología. Texcoco, Estado de México, 12-13 p. [ Links ]
IBARRA-NÚÑEZ, G.; MORENO-MOLINA, E. B.; RUIZ-COLMENARES, A.; TRUJILLO-OLIVERA, M.; GARCÍA-BALLINAS, J. A. 2004. Las arañas tejedoras (Araneidae, Tetrag-nathidae, Theridiidae y Uloboridae) de una plantación de cacao en Chiapas, México. pp. 38-41. In: Morales-Moreno, A.; Ibarra-González, M.; Rivera-González, A. P.; Stanford-Camargo, S. (Eds.). Entomología mexicana. Vol. 3. Montecillo, Texcoco: El Colegio de Postgraduados. México. [ Links ]
LEVI, H. W.; CODDINGTON, J. 1991.Sytematics and evolution of spiders (Araneae). Annual Review of Ecology and Systematics 22: 565-92. [ Links ]
LÓPEZ, A. P. A.; DELGADO, V. H.; AZPEITIA, M. A.; CASTAÑEDA, C. R. 2000. Tecnología para la producción de cacao en Tabasco. INIFAP. Villahermosa. 73 p. [ Links ]
LUBIN, Y. D. 1978. Seasonal abundance and diversity of web-buil-ding spiders in relation to habitat structure on Barro Colorado Island, Panama. Journal of Arachnology 6: 31-51. [ Links ]
LLINAS-GUTIÉRREZ, J.; JIMÉNEZ, M. L. 2004. Arañas de humedales del sur de Baja California, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México. Serie Zoología 75 (2): 283-302. [ Links ]
MORENO, C. E. 2001. Métodos para medir la biodiversidad. MyT-Manuales y Tesis SEA. Vol.1. Zaragoza. 84 p. [ Links ]
NYFFELER, M. 1999. Prey selection of spiders in the field. Journal of Arachnology 27: 317-324. [ Links ]
NYFFELER, M.; SUNDERLAN, K. 2003. Composition, abundance and pet control potential of spider communities in agroecosys-tems: a comparison of European and US studies. Agriculture, Ecosystems & Environment 95: 579-612. [ Links ]
PENNINGTON, T. D.; SARUKHÁN, J. 1998. árboles tropicales de México Manual para la identificación de las principales especies. UNAM. México. 520 p. [ Links ]
PÉREZ-DE LA CRUZ, M.; DE LA CRUZ-PÉREZ, A. 2005. Diversidad de Teridiidos (Araneae: Theridiidae) en cuatro asociaciones florísticas, en el ejido "Las Delicias", Teapa, Sureste de México. Universidad y Ciencia 21 (41): 41-44. [ Links ]
PÉREZ-DE LA CRUZ, M.; SÁNCHEZ-SOTO, S.; ORTÍZ-GARCÍA, C. F.; ZAPATA-MATA, R.; DE LA CRUZ-PÉREZ, A. 2007. Diversidad de insectos capturados por arañas tejedoras (Arachnida: Araneae) en el agroecosistema cacao en Tabasco, México. Neotropical Entomology 36: 90-101. [ Links ]
PLATNICK, N. I. 2014. The World Spider Catalog. Versions 15.0. American Museum of Natural History, New York. Disponible en: http://research.amnh.org/entomology/spiders/catalog/index. html [Fecha revisión: 22 octubre 2014] [ Links ].
RIECHERt, S. E.; BISHOP, L. 1990. Prey control by an assemblage of generalist predators: Spiders in garden test systems. Ecology 71 (4): 1441-1450. [ Links ]
RYPSTRA, A. L.; CARTER, P. E.; MARSHALL, S. D. 1999. Architectural features of agricultural habitats and their impact on the spider inhabitants. Journal of Arachnology 27: 371-377. [ Links ]
SAMU, F. K.; SUNDERLAND, D.; SZINETÁR, C. 1999. Scale-dependent dispersal and distribution patterns of spiders in agricultural systems: a review. Journal of Arachnology 27: 325-332. [ Links ]
SUÁREZ-FORERO, D. A.; ÁLVAREZ, Z. R.; CORREA, R. M. M. 2011. Gremios ecológicos de arañas (Arachnida: Araneae) asociados a cultivos y su vegetación de borde en el estado de Durango y Zacatecas, México. Vidsupra 3 (2): 37-44. [ Links ]
SUNDERLAND, K. 1999. Mechanisms underlying the effects of spiders on pest populations. Journal Arachnology 27 (1): 308-316. [ Links ]
TURNBULL, A. L. 1973. Ecology of true spiders (Araneomorphae). Annual Review of Entomology 18: 305-348. [ Links ]
UETZ, G. W.; HODGE, A. M. 1990. Influence of habitat and prey availability on spatial organization and behavior of colonial web-building spiders. National Geographic Research 6: 22-40. [ Links ]
WISE, D. H. 1993. Spiders in Ecological Webs.Cambridge University Press. 328 p. [ Links ]
ZAR, H. J. 1974. Biostatistical Analysis. Pretince- Hall, INC. Londres, Inglaterra. 220 p. [ Links ]
Recibido: 16-feb-2014
Aceptado: 16-feb-2015
Citación sugerida:
DE LA CRUZ-PÉREZ, A.; PÉREZ-DE LA CRUZ, M.; SÁNCHEZ-SOTO, S.; TORRES-DE LA CRUZ, M. 2015. Fluctuación po-blacional de arañas (Araneae: Tetragnathidae, Pholcidae) en el agroecosistema cacao en Tabasco, México. Revista Colombiana de Entomología 41 (1): 132-138. Enero-Junio 2015. ISSN 0120-0488.
Anexo 1. Composición vegetal de cada estrato del agroecosistema de cacao en Teapa, Tabasco, México (agosto 2004 a julio 2005).

