










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.41 no.2 Bogotá July/Dec. 2015
Sección Control / Control
Surveillance and genotyping of Varroa destructor (Acari: Varroidae) parasitizing Apis mellifera jemenitica (Hymenoptera: Apidae) in Saudi Arabia
Vigilancia y análisis genotipico del ácaro Varroa destructor (Acari: Varroidae) parásito de Apis mellifera jemenitica (Hymenoptera: Apidae) en Arabia Saudi
YEHYA ALATTAL1,2, MOHAMMAD JAVED ANSARI1,3, AHMAD AL-GHAMDI1,4 and AREF SINGLE1,5
1 Ph. D. Chair of Engineer Abdullah Ahmad Bagshan for Bee Research, Department of Plant Protection, College of Food and Agriculture Sciences, King Saud University, PO Box 2460, Riyadh 11451, Kingdom of Saudi Arabia.
2 yalattal@ksu.edu.sa.
3 mjavedansari@gmail.com. Corresponding author.
4 aalkhazim@gmail.com.
5 asingl67@gmail.com.
Abstract: A total of 72 Apis melllifera jemenitica honeybee samples were collected from eight different provinces within Saudi Arabia. Samples consisted of adult worker bees and capped brood cells. All samples were analyzed to estimate Varroa destructor mite infestation levels, and isolated V. destructor mites (n = 90) were subjected to DNA analysis.Results revealed that all colonies sampled in this survey were infested with V. destructor mites. The overall averageinfestation levels were 5.5 ± 4.8% and 13.1 ± 10.6% on adult worker bees and in capped brood cells, respectively.Differences in worker infestation levels were not significant among sampling locations (F = 1.5; P = 0.19); however,infestation levels in capped brood cells were highly significant among locations (F = 5.2; P < 0.001). Moreover, resultsindicated that all V destructor mites from A. m. jemenitica were recognized as a Korean haplotype of V. destructor .These results should assist forthcoming mite management efforts in Saudi Arabia.
Key words: Varroa mite. Infestation levels. Yemeni honeybee. RFLP. Genotype.
Resumen: Un total de 72 muestras de abejas, Apis melllifera jemenitica , se obtuvieron de ocho diferentes provincias de Arabia Saudita. Las muestras consistieron en abejas obreras adultas y celdas de cría selladas. Se analizaron todaslas muestras para estimar niveles de infestación con el ácaro Varroa destructor . Estos ácaros aislados (n = 90) fueronsometidos a un análisis de ADN. Los resultados revelaron que todas las colonias muestreadas en este estudio estabaninfestadas de V destructor . Los niveles de infestación promedio general fueron 5,5 ± 4,8% y 13,1 ± 10,6% en abejasobreras adultas y en las celdas de cría selladas, respectivamente. Las diferencias en los niveles de infestación de lasobreras no fueron significativas entre los lugares de muestreo (F = 1,5; P = 0,19), sin embargo, los niveles de infestaciónen celdas de cría selladas fueron altamente significativas entre localidades (F = 5,2; P < 0,001). Por otra parte, losresultados indicaron que todos los ácaros V. destructor de A. m. jemeniticafueron reconocidos como haplotipo coreanode esta especie. Estos resultados servirán para esfuerzos futuros en el manejo de ácaros en Arabia Saudita.
Palabras clave: Ácaro de abejas. Niveles de infestación. Abejas Yemeni. RFLP. Genotipo.
Introduction
The mite, Varroa destructor (Anderson y Trueman, 2000) is one of the most destructive pests of the honeybee (Apismellifera L.) worldwide (Rosenkranz et al. 2010). Varroa mitehas two haplotypes that are able to reproduce successfullyon A. mellifera; the Japan-type is relatively non-virulentwhereas the Korea-type, is extremely virulent (Andersonand Trueman 1998; Anderson 2000). In Saudi Arabia, themite was first reported in 1987 (Al-Ghamdi 1990), perhapsthrough a shipment of introduced honeybees from Egypt, theusual package bee supplier of Saudi Arabia. Later, severalstudies reported on the occurrence and impact of Varroa mitethroughout the country (Al-Ghamdi 1990; Al-Ghamdi 2002).Yet, general infestation levels and genotyping of V. destructorin Saudi Arabia is still needed.
Beekeeping in Saudi Arabia is based on native as well as on the exotic honeybee subspecies, A. mellifera (Alattal et al.2014 and Alqarni et al. 2011). About 70 % of these coloniesare kept in log hives colonized by native honeybee, A. m.jemenitica (Al-Ghamdi et al. 2013). Disease monitoring isnominal or absent and control of Varroa mite species is set upon calendar bases with almost consistent high colony losses. Inaddition to Varroa mite species, adverse climatic conditions in Saudi Arabia are the most obvious causes of colony losses in the country (Al-Ghamdi et al. 2013; Alqarni etal. 2011).
In temperate areas, infestation levels are critical and honeybee colonies with no annual treatment againstVarroa mite will collapse within 2-3 years (Rosenkranzet al. 2010), but in the majority of tropical climates theinfestation levels remain under the injury threshold (Ritterand De Jong 1984). Apparently, it is not well explainedwhether the bee types of the tropics or climate contributemore to the documented stable honeybee-Varroa speciesrelationships (Rosenkranz 1999; Carneiro et al. 2007;Rosenkranz et al. 2010).
Ruttner (1988) described the native honeybee of Saudi Arabia as the smallest and the most heat and drought toleranthoneybee subspecies of A. mellifera. Drought and highaverage ambient temperatures are dominant in Saudi Arabia.Consequently, the infestation levels and buildup of Varroaspecies population density under Saudi Arabia conditionsmay be different than that in the temperate regions whereabundant flows of nectar and pollen lead to profoundlypopulated colonies causing an exponential population growthof the parasite (Renz 2003). This study focuses on mitehaplotyping and on assessment of general infestation levelsof Varroa mite species in Saudi Arabia.
Materials and methods
Determination of infestation levels. Samples were taken from eight locations representing beekeeping areas withinSaudi Arabia. Samples were collected shortly after thesummer season during September and October 2013. Samplescolonies were last treated with acaricides eight months beforesample collection. Some related metereological parametersof the sampled locations are shown in Table 1. Varroa speciesinfestation levels of adult worker bees were anticipated bysampling 100 adult workers from brood combs using plasticjars (450 cm3) and were then stored at -20 °C. To determinethe number of adult Varroa mite species in each sample, thejars were filled with water and detergent and shaken for 2-3min. and were then given into a double-layer honey sievewhere adult bees were separated from Varroa mite speciesby a jet of water and the infestation level was calculated asnumber of Varroa mites/ number of adult bees and expressedin percentage.
Brood infestation rates were estimated by inspecting about 100 capped worker brood cells. After being frozen,the capping’s of the frozen brood samples were cut and thebrood with the Varroa mite were rinsed with water in thedouble-layer honey sieve to separate Varroa mites that werethen counted to determine brood infestation. Then, relativeinfestation ratio = brood infestation rates/worker infestationrates (Renz 2003) was calculated.
Comparation of means were computed based on ANOVA and LSM performed on data after transformation using arcsin function (SAS 1989).
V. destructor haplotype identification. A number of female adult mites isolated from worker or brood samples werecollected (n = 10 mites per apiary). Collected mites wereplaced in 1.5 micro-centrifuge tubes containing 96% ethanol,labeled and stored at -20 °C until DNA was extracted. Totalgenomic DNA was extracted from individual female mitesby chelex resinTM according to Cano et al. (1993). After thatextracted DNA was used to amplify the cytochrome oxidasesubunit one (CO-I) region of the mtDNA with the primers(COXF) 5'-GG(A/G)GG(A/T)GA(C/T)CC(A/T)ATT(C/T)T(A/T)TATCAAC-3' and (COXRa) 5'-CCTGT(A/T)A(A/t)AATA GCAAATAC-3' using PCR (Saiki et al. 1988)following the method described by Anderson and Fuchs(1998). PCR products were then separated by electrophoresis,stained with ethidium bromide, visualized with UV light(260 nm) and photographed. Amplified CO-I region wasthen used as a template for the RFLP reaction. The restrictionenzymes Sac-I, which distinguish the G/AGCTC sequenceand Xol-I which distinguish the C/TCGAG sequence wereadded separately each to 12.5 pl of the PCR product asdescribed by Anderson and Fuchs (1998). The mixture wasthen incubated at 37 °C for about one hour and was thenrun in a 1.5% agarose gel. The resulting bands were thenvisualized with UV light and photographed. One amplifiedCO-I region from each apiary were then sequenced using anautomated 96 capillary ABI 3730XI DNA genetic analyzer(Applied Biosystem). Sequences were manually checked andassembled using GeneBank database (NCBI). Sequenceswere then exposed to two procedures. First, sequences werealigned using CLUSTALW software (Thompson et al. 1994),BLASTed and compared with other sequences availableon the GenBank database (NCBI). Sequences were finallysubmitted to Genebank data base (NCBI).
Results
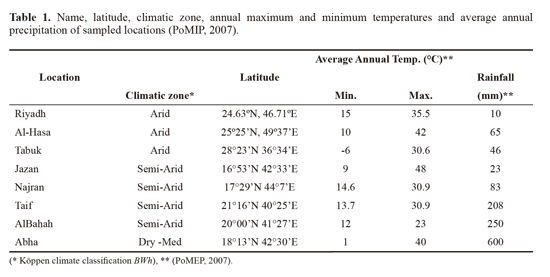
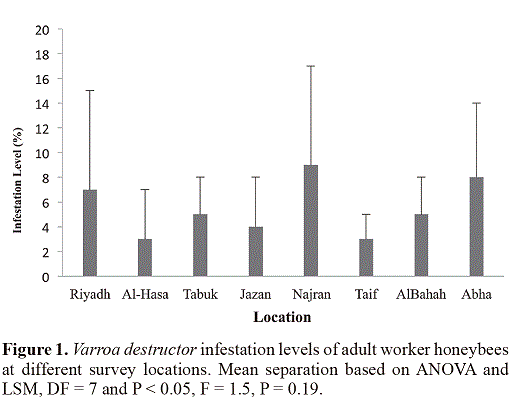
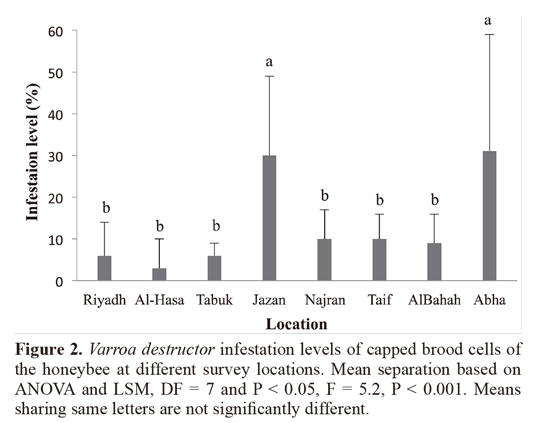
Results revealed that all colonies sampled in this survey (n = 72) were infested with Varroa mite species. The generalaverage infestation levels were 5.5 ± 4.8% and 13.1 ± 10.6%on adult worker bees and in capped brood cells, respectively.Differences in infestation levels of adult worker bees werenot significant among sampling locations (F = 1.5, P = 0.19,Figure 1), however, infestation levels in capped brood cellswere highly significant among locations (F = 5.2, P < 0.001,Figure 2) the highest brood infestation levels were in Jazanthen Abha. Generally, relative infestation of capped workerbrood was about 4.7 ± 6.3 times higher than the infestationon adult bees, and differences between locations weresignificant (P < 0.001, Figure 3). The highest value beingin Jazan.
The banding pattern of the RFLP reaction and sequence analysis of the mtDNA revealed that all mite samples were ofV. destructor Korean haplotype. Fragment digestion occurredonly when using Xol-I and resulted bands had a fragment sizebetween 200 and 300 bp. Sequences were 99 to 100% identicalto that of Korean type of V. destructor previously describedby Anderson and Trueman (2000) (GeneBank accessionnumber AF106899). In all Varroa mite samples analysed, twogenotypes were detected with more than 99% similarity. Thegenotypes were deposited in the NBCI GenBank under theaccession numbers KJ507740 and KJ507741.
Discussion
The survey demonstrated that V destructor Korean haplotype is distributed in all beekeeping areas within Saudi Arabia;results also indicated that relative infestation levels mayreach higher levels within few months without Varroatreatment. However these infestation levels are lower thanwhat was described for other V. destructor-Apis mellifera systems with different subspecies such as A. m. carnica incentral Europe (Rosenkranz et al. 2010), A.m. syriaca systemin Jordan (Alattal et al. 2005) or A.m. lamarckii in Egypt(Ismail et al. 2006). This epidemic spread of the mite withinSaudi Arabia could be mainly related to mass importationof Varroa species infested package bees from neighboringcountries mainly Egypt. In addition to poor managementpractices in Varroa mite control, an un-controlled migrationof bee colonies among locations increases the contactbetween highly infested colonies, where Varroa mite speciescan build up more quickly such as Jazan and Abha, andcolonies with low infestation. When highly infested coloniesdie, its remaining bees with the Varroa mites will enter newcolonies. Even that damage and colony losses were notvisible during the assessment; increased colony mortalitydue to Varrroa associated viruses is probable after the end ofspring and till the end of winter. Although, several locationsshowed relative infestation levels of adult bees below 15%,a proposed economic threshold on adult worker bees intemperate regions (Toscano 1996), we believe that the impactof Varroa infestation under arid conditions with very limitednectar and pollen resources, such as Saudi Arabia, would bealso dramatic if colonies were not treated.
Although the significant differences in the infestation of worker brood cells among different locations might indicatesreal spatial differences in the course of colony development inthese areas, it was not possible to classify locations accordingto their climatic zones or climatic parameters in predictinghigh or low infestation rates.
This demonstrates that spatial variations in infestation levels are rather a result of interaction among various factorsincluding bee type, Varroa mite species genotype, and otherecological factors. Since only the highly virulent Koreangenotype has been reported in all sampling areas withinSaudi Arabia, variation in infestation levels may be relatedto other factors. On the other hand, nearby colonies in thesame apiary may have clearly uneven relative infestationrates, which might be related to the variations in the patternsof colony build-up and colony population growth rate duringthe season. Consequently, Varroa mite population increasedrapidly in some colonies than in others.
The infestation ratio is a sign of the relative number of reproducing mites in the honeybee colony. The maximumratio took place in Abha and Jazan, which demonstratesthat the increase in Varroa mite population is higher inthese locations compared to other locations of this study.Consequently, colony losses due to higher infestation levelsof adult workers at the time the brood area is reduced couldbe high. For that reason, monitoring of infestation levels bysampling of brood cells and adult bees is an essential practiceto prevent late treatment or multiple treatments.
On the other hand, results indicated an extremely low divergence in the CO-I of Varroa mite in an extensivegeographical area and all of these Varroa mite species wereof a Korean haplotype identity, this may also suggest thepresence of the same genotype in neighboring countries.The Varroa mite species may have a common origin in theinvestigated populations as also mentioned by Solignac et al.(2005) for the European Varroa mites.
Acknowledgements
This project was funded by the National Plan for Science, Technology and Innovation (MAARIFAH), King AbdulazizCity for Science and Technology, Kingdom of Saudi Arabia,Award number (11-BIO1743-02). We are grateful to thebeekeepers who allowed us to sample their hives and JamesMontoya Lerma (Editor in charge Revista Colombiana deEntomología) for writing Spanish "resumen" and editing themanuscript.
Literature cited
ALATTAL Y; ROSENKRANZ, P; ZEBITZ, C. W. P. 2005. Reproduction of Varroa destructor (Anderson and Trueman2000) in sealed worker brood cells of Apis mellifera carnicaand Apis mellifera syriaca in Jordan. pp. 256. In: Proceedings ofthe Tagung der DGaaE, 21-24, March 2005, Dresden, Germany. [ Links ]
ALATTAL, Y.; AL-GHAMDI, A.; ALSHARHI, M. 2014. Populationstructure of the Yemeni honey bee (Apis mellifera jemenitica )entails an urgent conservation strategy in Saudi Arabia. Journalof Entomology 11: 163-169. [ Links ]
AL-GHAMDI, A. A. 1990. Survey of honeybee diseases, pests and predators in Saudi Arabia. M. Phil. Thesis, University of Wales,UK. [ Links ]
AL-GHAMDI, A. A. K. 2002. The comprehensive study of the mite Varroa destructor on honeybees Apis melliefra indigenous andimported. Final report, King Abdulaziz City for Science andTechnology, MS-4-29. [ Links ]
AL-GHAMDI, A. A.; NURU, A.; KHANBASH, M. S.; SMITH, D. R. 2013. Geographical distribution and population variationof Apis mellifera jemenitica Ruttner. Journal of ApiculturalResearch 52: 124-133. [ Links ]
ALQARNI, A. S.; HANNAN, M. A.; OWAYSS, A. A.; ENGEL, M. S. 2011. The indigenous honey bees of Saudi Arabia(Hymenoptera, Apidae, Apis mellifera jemenitica Ruttner):Their natural history and role in beekeeping. ZooKeys 134: 8398. [ Links ]
ANDERSON, D. L.; FUCHS, S. 1998. Two genetically distinct populations of Varroa jacobsoni with contrasting reproductiveabilities on Apis mellifera . Journal of Apicultural Research 37:69-78. [ Links ]
ANDERSON, D. L. 2000. Variation in the parasitic bee mite Varroa jacobsoni Oud. Apidologie 31: 281-292. [ Links ]
ANDERSON, D. L; TRUEMAN, J. W. H. 2000. Varroa jacobsoni (Acari: Varroidae) is more than one species. Experimental andApplied Acarology 24: 165-189. [ Links ]
CANO, R. J.; HENDRIK, N. P; PIENIAZEK, N. J.; ACRA, A.; POINAR, G. O. 1993. Amplification and sequencing of DNAfrom a 120-135 million year old weevil. Nature 363: 536-538. [ Links ]
CARNEIRO, F. E.; TORRES, R. R.; STRPAZZON, R.; RAMÍREZ,S. A.; GUERRA, J. R.; KOLING, D. F.; MORETTO, G.2007. Changes in the reproductive ability of the mite Varroa destructor (Anderson and Trueman) in africanized honey bees (Apis mellifera L.) (Hymenoptera: Apidae) Colonies in SouthernBrazil. Neotropical Entomology 36: 949-952. [ Links ]
ISMAIL, A. M.; GHONIEMY, H.; OWAYSS, A. 2006. Combatting honeybee Varroa mites by plant oils alone or in an IPM program.The 2nd conference of Farm Integrated Pest Management, 1618 Jan., Faculty of Agriculture, Fayoum University, 172-185 pp. [ Links ]
PRESIDENCY OF METEOROLOGY AND ENVIRONMENT PROTECTION, National Meteorology and EnvironmentCenter. 2007. Surface Annual Climatological Report, Riyadh,Saudi Arabia. [ Links ]
RENZ, M. 2003. Die Populationsdynamik der Bienenmilbe Varroa destructor bei Völkern von Apis mellifera unterbesonderer Berücksichtigung der reinvasion. Ph.D, Institut fürParasitologie und internationale Tiergesundheit, FachbereichVeterinärmedizin, Freie Universität Berlin. [ Links ]
RITTER, W.; DE JONG, D. 1984. Reproduction of Varroa jacobsoni in Europe, the Middle East and tropical South America.Zeitschrift für Angewandte Entomologie 98: 55-57. [ Links ]
RUTTNER, F. 1988. Biogeography and taxonomy of honey bees. Springer-Verlag, New York, USA. [ Links ]
ROSENKRANZ, P. 1999. Honeybee Apis mellifera L. tolerance to Varroa jacobsoni Oud. in South America. Apidologie 30: 159172. [ Links ]
ROSENKRANZ, P.; AUMEIER, P; ZIEGELMANN, B. 2010. Biology and control of Varroa destructor . Journal of InvertebratePathology 103: 96-119. [ Links ]
SAIKI, R. K.; GELFAND, D. H.; STOFFEL, S.; SCHARF, S. J.; HIGUCHI, R.; HORN, G. T.; MULLIS, K. B.; ERLICH, H.1988. Primer-directed enzymatic amplification of DNA withthermostable DNA polymerase. Science 239: 487-91. [ Links ]
SAS, INSTITUTE. 1989. Proc User's manual, Version 6,4th ed. SAS Institute Inc, Cary, NC, USA. [ Links ]
SOLIGNAC, M.; CORNUET, J. M.; VAUTRIN, D.; CONTE, Y; ANDERSON, D. L.; EVANS, J.; CROS-ARTEIL, S.;NAVAJAS, M. 2005. The invasive Korea and Japan types of theVarroa destructor, ectoparasitic mite of the eastern honeybee(Apis cerana ), are two partly isolated clones. Proceedings of theRoyal Society of London 272: 411-419. [ Links ]
THOMPSON, J. D.; HIGGINS, D. G.; GIBSON, T. J. 1994. CLUSTAL-W: improving the sensitivity of progressive multiplesequence alignments through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic AcidsResearch 22: 4673-4680. [ Links ]
TOSCANO, H. 1996. Estado actual de algunas enfermedades apicolas en Uruguay, periodo 1985-1995. Reporte de ladireccion de laboratorios veterinarios 'Dr. MC. Rubino' (DI.LA.VE) pp. 6-8. [ Links ]
Received: 16-Dec-2014
Accepted: 6-Nov-2015
Suggested citation:
ALATTAL, Y; ANSARI, M. J.; AL-GHAMDI, A.; SINGLE, A. 2015. Surveillance and genotyping of Varroa destructor (Acari:Varroidae) parasitizing Apis mellifera jemenitica (Hymenoptera: Apidae) in Saudi Arabia. Revista Colombiana de Entomologia 41 (2): 180-183. Julio - Diciembre 2015. ISSN 01200488.

