










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.42 no.1 Bogotá Jan.,/June 2016
SECCIÓN BÁSICA / BASIC
ARTÍCULOS DE INVESTIGACIÓN / RESEARCH PAPER
Biología de Diaphorina citri (Hemiptera: Liviidae) bajo condiciones de invernadero en Palmira, Colombia
Biology of Diaphorina citri (Hemiptera: Liviidae) under glass house conditions in Palmira, Colombia
Yesenia GarcíaI; Yaneth Patricia RamosII; Paola Andrea SoteloIII; Takumasa KondoIV
IIngeniera Agrónoma. Universidad Nacional de Colombia, Sede Palmira, Colombia, camilitaroj@gmail.com
IIIngeniera Agrónoma. Corporación Colombiana de Investigación Agropecuaria (CORPOICA), Centro de Investigación Palmira, Calle 23 Carrera 37 Continuo al Penal, Palmira, Colombia, yramos@corpoica.org.co
IIIPh. D., Corporación Colombiana de Investigación Agropecuaria (CORPOICA), Centro de Investigación Obonuco, Km. 5 vía Pasto-Obonuco, Pasto, Colombia, pasotelo@corpoica.org.co
IVPh. D., Corporación Colombiana de Investigación Agropecuaria (CORPOICA), Centro de Investigación Palmira, Calle 23 Carrera 37 Continuo al Penal, Palmira, Colombia, tkondo@corpoica.org.co, autor para correspondencia
RESUMEN
El estudio se realizó bajo condiciones de invernadero (temperatura máxima: 37,1 ± 5,1 °C, temperatura mínima: 25,0 ± 2,0 °C; humedad relativa máxima: 67,0 ± 6,2%, humedad relativa mínima: 39,5 ± 10,9%; 12: 12 h Luz: Oscuridad, régimen natural de luz) en el municipio de Palmira, Valle del Cauca. Se determinaron características morfológicas de cada estadio de desarrollo, tiempo de duración, sobrevivencia, parámetros ecológicos, capacidad reproductiva, longevidad de adultos y mortalidad específica por estadio de Diaphorina citri (Hemiptera: Liviidae) en brotes de mirto (Murraya paniculata). El tiempo desde huevo hasta adulto fue de 15,41 ± 0,49 días. El tiempo medio de supervivencia para las hembras fue de 48,25 días y para machos de 50 días. El 6,86% de las posturas de D. citri llegó al estado adulto, presentándose la mayor mortalidad en los estadios de ninfa I (81%). Las hembras presentaron posturas promedio de 237 huevos durante su ciclo reproductivo. Se presenta una clave dicotómica para separar los diferentes estados de desarrollo del psílido asiático de los cítricos, herramienta indispensable para realizar estudios de biología. Estos estudios sobre la biología de D. citri servirán como base en la cría masiva de su parasitoide T. radiata.
Palabras clave: Ciclo de vida. Morfología. Mortalidad. Murraya paniculata. Parámetros poblacionales.
ABSTRACT
In this study, the morphological characteristics of each stage of development, duration, survival, ecological parameters, reproductive capacity, longevity of adults, and stage-specific mortality of Diaphorina citri (Hemiptera: Liviidae) were determined. It was conducted in a greenhouse (maximum temperature: 37.1 ± 5.1 °C, minimum temperature: 25.0 ± 2.0 °C; maximum relative humidity: 67.0 ± 6.2%, minimum relative humidity: 39.5 ± 10.9%; 12: 12 h Light: Dark, natural light conditions) in the municipality of Palmira, Valle del Cauca. Mean time from egg to adult was 15.41 ± 0.49 days. The median survival time for females was 48.25 days and 50 days for males. The survival rate to adult stage of D. citri was 6.86% with the highest mortality rate recorded for first-instar nymphs (81%). Females oviposited an average of 237 eggs during their reproductive cycle. A dichotomous key, an essential tool for conducting biological studies, is provided to separate the different growth stages of the Asian citrus psillid. Studies on the biology of D. citri will serve as a baseline for the mass rearing of its parasitoid, T. radiata.
Key words: Life cycle. Morphology. Mortality. Murraya paniculata. Population parameters.
Introducción
El psílido asiático de los cítricos, Diaphorina citri Kuwayama (Hemiptera: Liviidae) se encuentra ampliamente distribuido en el sur de Asia y fue recientemente introducido a Norte América y Hawái (Mead y Fasulo 2010), aunque se había reportado en el Nuevo Mundo, en Brasil (Costa Lima 1942; Catling 1970). Actualmente, D. citri se distribuye principalmente en áreas tropicales y subtropicales y se reporta en las siguientes áreas geográficas: El Caribe (Bahamas, Islas Caimán, Jamaica, República Dominicana, Cuba, además de interceptaciones de St. Thomas y Belice), América Central (Guadalupe y México,), África (Isla de la Reunión y Mauricio), Asia (Afganistán, Arabia Saudita, China, Hong Kong, India, Indonesia, Malasia, Myanmar, Nepal, Pakistán, Filipinas, Islas Ryukyu, Sri Lanka, Taiwán y Tailandia), América del Sur (Argentina, Brasil y Venezuela) y Norte América (Estados Unidos y algunos de sus territorios, i.e., Puerto Rico, Guam, islas Marianas del Norte, islas Vírgenes de Estados Unidos y Samoa Americana) (Halbert y Núñez 2004; Mead y Fasulo 2010). El descubrimiento de D. citri en Arabia Saudita (Wooler et al. 1974) fue el primer registro en el Cercano Oriente.
Otra especie de psílido, Trioza erytreae (Del Guercio) también ocurre en Arabia Saudita, prefiriendo las zonas orientales y tierras altas donde los climas son extremos, mientras que D. citri está presente a lo largo de las zonas costeras occidentales (Mead y Fasulo 2010). En Colombia, D. citri fue reportada por primera vez en el 2007 y desde entonces se ha registrado en los departamentos de Antioquia, Atlántico, Bolívar, Caldas, Cauca, Cesar, Córdoba, Cundinamarca, Magdalena, Meta, Norte de Santander, Quindío, Risaralda, Santander, Sucre, Tolima, y Valle del Cauca (ICA 2010; Kondo y Simbaqueba 2014).
Los daños causados por D. citri resultan por la remoción de grandes cantidades de la savia de las hojas y de servir como vector de la bacteria causante de la enfermedad catastrófica llamada "huanglongbing" (HLB) o "enverdecimiento de los cítricos" (Mead y Fasulo 2010). El HLB también se conoce como clorosis de los cítricos en Java, moteado de las hojas y moteado de hojas amarillas en las Filipinas, likubin (descenso rápido) en Taiwán, y huanglongbing en China, que se traduce como "enfermedad del dragón amarillo" o a menudo "enfermedad de brotes amarillos" (Mead y Fasulo 2010). El HLB es responsable de la destrucción de las industrias citrícolas de varios países de Asia y África (Manjunath et al. 2008). En el 2015, en Colombia se reportó una nueva especie de bacteria del mismo grupo que el HLB, determinada como Candidatus Liberibacter caribbeanus (Manjunath et al. 2015). Hasta finales del 2015, el HLB no había sido detectado en Colombia (Ángel et al. 2014), sin embargo, en diciembre 2015, el Instituto Colombiano Agropecuario (ICA), declaró una emergencia fitosanitaria en territorio colombiano al detectar adultos de D. citri que dieron positivos para la bacteria Candidatus Liberibacter asiaticus en el departamento de la Guajira (ICA 2015).
Dentro de los enemigos naturales de D. citri se incluyen sírfidos, crisópidos, coccinélidos, avispas parasitoides y hongos entomopatógenos (González et al. 2002; Rodríguez-Palomera et al. 2012). En Colombia hay pocos estudios sobre inventarios de enemigos naturales de D. citri, con excepción de reportes sobre su enemigo natural principal, el ectoparasitoide Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae) (Ebratt et al. 2011a, b; Kondo et al. 2012). Kondo et al. (2015a) reportaron a la libélula Erpetogomphus sabaleticus Williamson, 1918 (Odonata: Gomphidae) depredando al adulto de D. citri en el Valle del Cauca. Más recientemente, Kondo et al. (2015b) enlistan un total de 16 especies de enemigos naturales de D. citri distribuidos en seis familias y cinco órdenes en Colombia. Con base en la literatura científica presentan un listado actualizado de 95 artrópodos (arácnidos e insectos) enemigos naturales de D. citri en nueve órdenes y 23 familias en el mundo. Debido a la importancia de D. citri como vector de la bacteria causante del HLB, el control de D. citri es un componente crítico en la prevención, contención y manejo de la enfermedad.
El objetivo de esta investigación fue estudiar la biología de D. citri sobre brotes tiernos de mirto [Murraya paniculata L. (Jack) (Rutaceae)] para determinar el tiempo de desarrollo de cada uno de los estados del insecto y evaluar cualitativa y cuantitativamente las características morfológicas de cada estado (tamaño, color y otros cambios físicos), tiempo de duración de cada estado ninfal, movilidad, preferencia de ubicación en la planta, sobrevivencia, parámetros ecológicos, capacidad reproductiva, longevidad de adultos y mortalidad específica por estadio.
Materiales y métodos
El estudio se realizó en las instalaciones de Corpoica, Centro de Investigación Palmira, en el municipio de Palmira, departamento del Valle del Cauca, Colombia, 03°30'38,7"N 76°18'55,3"O, 999 msnm, en un invernadero (temperatura máxima: 37,1 ± 5,1 °C, temperatura mínima: 25,0 ± 2,0 °C; humedad relativa máxima: 67,0 ± 6,2%, humedad relativa mínima: 39,5 ± 10,9%; 12: 12 h Luz: Oscuridad, régimen natural de luz). Los datos climáticos se midieron con un registrador de datos (Datalogger CEM, DT 171, Shenzhen Everbest Machinery Industry Co., Ltd, Shenzhen, China). Para la cría de D. citri, se dispuso de una planta de mirto Murraya paniculata de un año de edad con tres brotes tiernos, teniendo en cuenta que el psílido asiático de los cítricos siempre oviposita en la base o en las puntas de brotes tiernos (Étienne et al. 2001). La planta se cubrió totalmente con un cilindro de acetato calibre 12 de 80 cm de diámetro por 53 cm de alto con dos perforaciones laterales de 15 cm de diámetro, cubiertas al igual que el extremo superior con una tapa de tela velo. Allí se liberó una hembra grávida de D. citri durante 24 horas como tiempo de oviposición. Pasadas las 24 horas se retiró la hembra y se obtuvo una cohorte de 102 huevos de la misma edad. Con la ayuda de una lupa 30X se observó el desarrollo de la cohorte tomando datos cada 24 horas para identificar las características cualitativas que permitieron diferenciar cada estado de desarrollo del insecto. Las medidas de los diferentes estados de desarrollo se tomaron con el uso de microscopio estereoscópico Nikon AMZ-1500 con un micrómetro ocular. La longitud y el ancho se tomaron midiendo la distancia máxima entre la cabeza y el ápice del abdomen; y el ancho del insecto en la región del tórax, respectivamente. Se midieron tres especímenes por estado de desarrollo los cuales fueron tomados al azar. Las medidas se presentan como rango. Se realizaron observaciones diarias para determinar la duración y características de cada uno de los estados del desarrollo de D. citri cuantificando el tiempo de duración en días, partiendo de huevos depositados en M. paniculata (N=102). Los datos de supervivencia de hembras y machos se generaron a partir de 20 parejas de D. citri que además fueron usados para hacer los estudios de tabla de vida.
Análisis estadístico. Todos los análisis estadísticos se hicieron con el programa estadístico SAS (SAS Institute Inc., 2012). Los datos de supervivencia para hembras y machos se analizaron mediante el procedimiento PROC LIFETEST, el cual permite hacer un análisis de supervivencia de las curvas observadas en la supervivencia de los individuos que alcanzaron el estado adulto mediante la prueba de igualdad de Log-Rank. Los parámetros poblacionales se calcularon mediante la técnica Jackknife y con base en los datos de fecundidad y la longevidad mediante el uso de una plataforma de SAS para estimar parámetros de la tabla de vida (Maia et al. 2000).
Parámetros poblacionales. Con los datos obtenidos de fecundidad, longevidad y ciclo de vida de D. citri, se construyó una tabla de vida y se calcularon los parámetros poblacionales como se describen a continuación: tasa reproductiva neta Ro = Slxmx, tiempo medio de generación T = Sxlxmx/Ro, tasa intrínseca de crecimiento rm = lnRo/T, y tiempo de doblaje Dt = ln2/rm (Andrewartha y Birch 1954; Silveira Neto et al. 1976; Rabinovich 1978; Southwood 1978; Maia et al. 2000).
Resultados y discusión
Características morfológicas de los estados de desarrollo de Diaphorina citri
Huevo (Figs. 1A, 2). La hembra grávida de D. citri oviposita los huevos en brotes tiernos (Fig. 1A), la base foliar y el envés de las hojas tiernas. Los huevos (Fig. 1A, primer plano) son de forma ovoide y tienen una prolongación alargada terminada en punta en el extremo que queda expuesto; son de color amarillo cuando están recién ovipositados y se tornan de color naranja con dos puntos rojos que señalan la posición de los ojos de los embriones cuando los huevos están a punto de eclosionar; miden 0,30 (0,28-0,31) mm de longitud y 0,12 (0,11-0,13) mm de ancho. Los huevos que no terminan su desarrollo se caracterizan por su apariencia deshidratada. Las medidas del huevo tomadas en este estudio concuerdan con las de Fonseca et al. (2007) y Tsai y Liu (2000) quienes reportaron un tamaño de huevo de 0,29 mm y 0,31 mm de longitud, respectivamente.
Estado ninfal I (N1). Se caracteriza por no tener esbozos alares (alas en formación), con antenas pequeñas y de coloración similar al resto del cuerpo. Son de color amarillo-anaranjado y tienen ojos compuestos de color rojo (Fig. 1B). Las ninfas N1 son móviles, miden de 0,24-0,31 mm de longitud y de 0,10-0,18 mm de ancho. Se agrupan debajo de las hojas tiernas donde se alimentan. Secretan abundante cera de color blanco de forma ondulada. El tamaño de las ninfas N1 en este estudio es menor al reportado por Fonseca et al. (2007) quienes registraron un tamaño de 0,35 mm. Por otro lado nuestras medidas concuerdan con aquellas obtenidas por Tsai y Liu (2000) con una longitud promedio de 0,30 mm y un ancho de 0,17 mm.
Estado ninfal II (N2). Se reconoce por tener esbozos alares apenas en desarrollo, antenas totalmente blanquecinas o con tintes de color negro solo en la punta (Fig. 1C). Las ninfas N2 son de color amarillo-anaranjado, móviles, con ojos compuestos, de color rojo, miden de 0,47-0,50 mm de longitud y de 0,27-0,30 mm de ancho (Fig. 1C). Se agrupan en el envés de las hojas tiernas y al igual que las ninfas N1 secretan abundante cera de color blanca de forma ondulada. El tamaño de las ninfas N2 en este estudio es ligeramente mayor a las medidas reportadas por Fonseca et al. (2007) y Tsai y Liu (2000) quienes reportan una longitud de 0,44 mm y 0,45 mm, respectivamente.
Estado ninfal III (N3). Las bases de los botones alares están presentes por debajo del nivel de los ojos y la margen posterior de estos no se extiende más allá del segundo segmento abdominal. Las antenas se tornan negruzcas solo en la parte distal. Los ojos son compuestos, de color rojo (Fig. 1D). Las ninfas N3 son muy móviles, miden de 0,95-1,00 mm de longitud y de 0,71-0,75 mm de ancho. Secretan cera en menos cantidad que las ninfas N1 y N2 y son de forma esférica. Se ubican en grupos en el envés de las hojas tiernas y en algunos casos migran hacia las bases foliares o a las ramas. Las medidas de las N3 en este estudio fueron ligeramente mayores a las medidas reportadas por Fonseca et al. (2007) y Tsai y Liu (2000) quienes reportaron una longitud de 0,76 mm y 0,74 mm, respectivamente.
Estado ninfal IV (N4). Se reconoce por tener esbozos alares aún más desarrollados que los previos instares ninfales, extendiéndose anteriormente hasta tocar la parte posterior del ojo o un tercio de los ojos y posteriormente hasta el tercer segmento abdominal. La parte media distal de las antenas son frecuentemente negruzcas; con ojos compuestos de color rojo (Fig. 1E). Las ninfas N4 son bastante móviles, miden de 1,40-1,52 mm de longitud y de 1,09-1,13 mm de ancho. Se ubican en grupos como medida de sobrevivencia y descienden de los brotes tiernos a las ramas más desarrolladas de la planta. Las medidas de las ninfas N4 en este estudio son mayores a las reportadas por Fonseca et al. (2007) y las de Tsai y Liu (2000) quienes reportan una longitud de 1,05 mm y 1,01 mm, respectivamente.
Estado ninfal V (N5). Mide de 1,61-1,66 mm de longitud y de 1,10-1,12 mm de ancho; es muy móvil; tiene esbozos alares bien desarrollados que se extienden anteriormente hasta el margen frontal de los ojos o sobrepasando los ojos y posteriormente hasta el cuarto segmento abdominal. Las antenas frecuentemente son negruzcas en su totalidad a excepción del escapo; con ojos compuestos de color rojo (Fig. 1F). Frecuentemente con un abdomen de coloración verde azul o anaranjado-amarillento; se agrupan en las ramas quedándose ahí hasta la emergencia del adulto. Las medidas de las ninfas N5 en este estudio son similares a las reportadas por Fonseca et al. (2007) y Tsai y Liu (2000) quienes reportan una longitud de 1,66 mm y 1,60 mm, respectivamente.
Adultos. Se reconocen por la posición de descanso que toman sobre el sustrato formando un ángulo de 45 grados (Fig. 2). Tienen alas moteadas de color castaño, ojos compuestos de color rojo; antenas pequeñas con una coloración negra en la punta (Figs. 1G y 2). El adulto recién emergido tiene una coloración blanquecina y en el transcurso de horas cambian a su color definitivo. Miden de 2,24-2,30 mm de longitud y de 0,61-0,65 mm de ancho. El dimorfismo sexual se presenta en el ápice del abdomen; la hembra tiene un abdomen finalizado en punta fina y el macho tiene un abdomen finalizado en punta roma (Fig. 1G). Las medidas del estado adulto en este estudio fueron menores a las reportadas por Fonseca et al. (2007) quienes registraron una longitud de 3,1 mm para la hembra y 2,5 mm para el macho. Por otro lado Tsai y Liu (2000) obtuvieron medidas de 3,3 mm de largo por 1,0 mm de ancho para la hembra y 2,7 mm de largo por 0,8 mm de ancho para el macho. Para las medidas del estado adulto no se diferenció el tamaño de los sexos en el presente estudio.
Los vuelos de D. citri son normalmente cortos de menos de 1 metro de distancia, volando entre las ramas de una planta o entre plantas adyacentes. Sin embargo, en estudios realizados en Florida, Estados Unidos, se determinó que el psílido asiático de los cítricos cada mes vuela una distancia entre 30-100 metros (Hall y Hentz 2011).
Ciclo de vida
El tiempo desde huevo hasta adulto fue de 15,41 ± 0,49 días. En la Tabla 1 se resume la duración de cada estado de desarrollo de D. citri. Solo 7 individuos llegaron al estadio de adulto, representando un 6,86% de la cohorte inicial (n =102 huevos). Los adultos de esta cohorte vivieron más de 64 días bajo las condiciones establecidas en casa de vidrio. La relación de sexos fue 1: 1.
Debido a la cantidad mínima de individuos que llegaron a etapa adulta durante el ciclo de vida, se partió de un grupo inicial de 20 parejas a las cuales se les evaluó la longevidad y los parámetros ecológicos requeridos para los análisis de la tabla de vida. Sin embargo, 8 de las hembras no presentaron oviposición durante el experimento y por lo tanto fueron excluidas de los estudios ecológicos. La longevidad entre hembras y machos de D. citri no fue significativamente diferente (Log-Rank χ2 = 0,27 df = 1, P < 0,6031) en condiciones de invernadero. El tiempo medio de supervivencia para las hembras fue de 48,25 días (95% intervalo de confianza (IC) percentil 50: 35-59 días, n = 12), y el tiempo medio de supervivencia para los machos fue de 50 días (IC del 95% percentil 50: 41-60 días, n = 12) (Fig. 3).
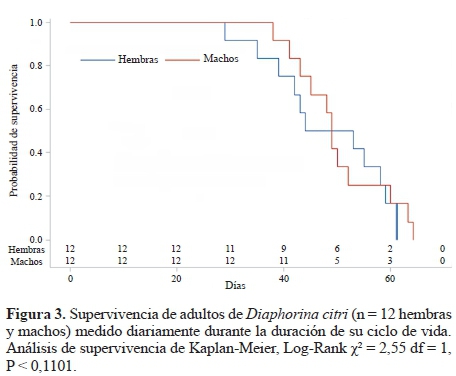
La duración del ciclo de vida de D. citri bajo condiciones de invernadero concuerdan con los de Fernández y Miranda (2005) quienes reportaron el tiempo de duración desde huevo hasta adulto entre 16 a 17 días a una temperatura de 25 °C. Por otro lado, Nava et al. (2007) encontraron que el tiempo de desarrollo de D. citri sobre M. paniculata dura 17,74 días a una temperatura de 24 °C y una HR de 70 ± 10%. En estudios anteriores, Morales et al. (2010) reportaron diferencias en el ciclo de vida de D. citri dependiendo de la planta hospedera, 15,2 (± 2,8) días sobre lima Citrus aurantifolia (Christm.) Swing, 17,4 (± 2,1) días sobre naranja Citrus sinensis (L.) Osbeck, 18,8 (± 2,8) días sobre mandarina Citrus reticulata Blanco, 13,6 (± 1,2) días sobre pomelo Citrus grandis (L.) Osbeck y 15,8 (± 2,8) días sobre mirto M. paniculata, bajo condiciones de laboratorio a una temperatura de 24,89 (± 1,48) °C y humedad relativa de 64,89% (± 4,95%). Por otro lado, los resultados del presente estudio coinciden con los de Tsai y Liu (2000) quienes estudiaron la biología de D. citri sobre Citrus jambhiri Lush, C. aurantium L., C. x paradisi Macfad. y M. paniculata en condiciones de laboratorio con una temperatura de 25 °C, obteniendo resultados similares entre hospederos en cuanto a duración de ciclo de vida con un promedio de 17,2 días. Según Liu y Tsai (2000), la temperatura tiene un efecto significativo en la tasa de desarrollo, supervivencia, reproducción y longevidad de D. citri. De acuerdo a Catling (1970), el rango de la temperatura adecuada para el desarrollo de D. citri es de 25-26 °C, coincidiendo con la temperatura en este estudio (= 27 ± 2 °C).
Parámetros poblacionales
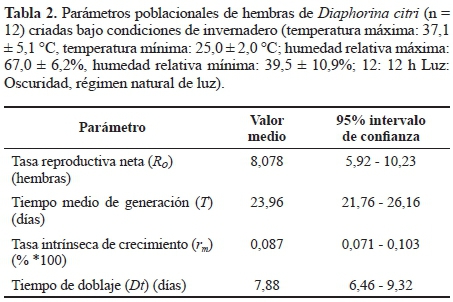
Respecto a los parámetros poblacionales, se observó un periodo de preoviposición aproximado de dos días y posteriormente se observaron posturas regulares hasta los 49 días con un pico de oviposición a los 35 días (Fig. 4), lo cual coincide con el promedio de supervivencia de las hembras. Las hembras presentaron posturas promedio de 237,6 huevos (rango 91-362 huevos) durante su ciclo reproductivo. Así mismo, se observó que no hubo contribución reproductiva de hembras mayores a 50 días (Fig. 4). Así mismo, los estudios poblacionales mostraron que una hembra de D. citri es reemplazada por aproximadamente 8 hijas, lo cual supone un crecimiento constante de la población (Southwood 1978; Rabinovich 1980) (tasa reproductiva neta (Ro) = 8,078) (Tabla 2). Adicionalmente, la tasa intrínseca de crecimiento natural (rm) fue de 0,087 (Tabla 2), indicando que la población de D. citri puede crecer casi un 9% bajo las condiciones climáticas evaluadas. En cuanto al tiempo generacional (T), se estimó en 23 días (Tabla 2), es decir, que desde que un huevo de D. citri es puesto, se requieren de 23 días para que una hembra proveniente de esa generación empiece a ovipositar. Esto supondría que se podrían obtener aproximadamente 16 generaciones por año bajo las condiciones evaluadas en este estudio. El tiempo de duplicación (Dt) fue de 7,86 días, indicando el periodo de tiempo requerido para duplicar la población inicial bajo las condiciones evaluadas.
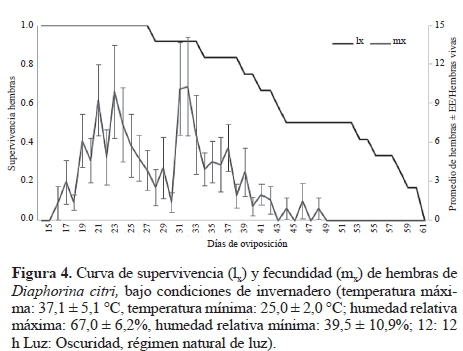
Los resultados obtenidos mediante el análisis de los parámetros ecológicos muestran que hay una mortalidad muy alta en el primer estadio ninfal, cercana al 81% y solo un 7% de la población inicial llega a estado adulto. En términos de control de D. citri, se puede pensar en estrategias de manejo enfocadas al primer estadio de desarrollo y por lo tanto las acciones deben lograrse rápidamente durante la primera semana de presencia de adultos en los cultivos de acuerdo a la duración de los estadios de huevo y primer instar ninfal. Los daños directos causados por D. citri no son considerados graves, pero adquiere una gran importancia por servir como vector del HLB, ya que un solo espécimen que sobreviva a las estrategias de manejo de la plaga supone un potencial portador de la enfermedad. En promedio, una hembra puede tener 237 posturas de las cuales solo el 7% llegaría a estado adulto, es decir la población sobreviviente seria de 17 adultos, todos potenciales transmisores de la enfermedad HLB. Por esta razón, un control racional mediante el uso de insecticidas sistémicos y de contacto sigue siendo importante para evitar la diseminación de la enfermedad. Para el uso de agentes de control biológico se debe tener en cuenta los periodos de carencia ya que estos suelen ser más susceptibles a estos productos que D. citri.
Clave para separar los estados ninfales y el adulto de
Diaphorina citri
1. Insecto con alas bien desarrolladas, moteadas ................. 2
- Insecto con esbozos alares (alas en desarrollo) ................. 3
2. Abdomen puntiagudo (Fig. 1G, izquierda): ................................................................................................ adulto, hembra
- Abdomen en punta de romo (Fig. 1G, derecha): ............................................................................................ adulto, macho
3. Especímenes entre 0,28-0,50 mm de longitud. Botones alares ausentes o muy poco desarrollados; antenas blanquecinas, sin tintes oscuros, segmentación de las antenas no evidente .................................................................................... 4
- Especímenes entre 0,74-1,66 mm de longitud. Botones alares más desarrollados; antenas con coloración oscura, especialmente en las puntas, algunos especímenes pequeños con antenas blanquecinas, segmentación de las antenas evidente …………..…………………….………….………….......... 5
4. Especímenes entre 0,28-0,35 mm de longitud. Antenas completamente blanquecinas. Botones alares no desarrollados (Fig. 1B): ..................................................... Ninfa I (N1)
- Especímenes entre 0,44-0,50 mm de longitud; Antenas a veces con tintes oscuros en la punta. Botones alares rudimentarios (Fig. 1C): ................................................ Ninfa II (N2)
5. Ninfas entre 1,60-1,66 mm de longitud; botones alares bien desarrollados que se extienden anteriormente hasta el margen frontal de los ojos o sobrepasando los ojos y posteriormente hasta el cuarto segmento abdominal; antenas frecuentemente oscuras en su totalidad a excepción del escapo (Fig. 1F):........................................................... Ninfa V (N5)
- Ninfas entre 0,74-1,52 mm de longitud; botones alares bien desarrollados pero no se extienden anteriormente hasta el margen frontal de los ojos ni hasta el cuarto segmento abdominal; antenas no tan oscuras, máximo hasta la mitad de la longitud de las antenas ..................................................... 6
6. Especímenes miden entre 0,74-1,00 mm de longitud; base de los botones alares presentes por debajo del nivel de los ojos, margen posterior de los esbozos alares no extendiéndose más allá del segundo segmento abdominal; antenas sólo oscuras en la parte distal (Fig. 1D): ................ Ninfa III (N3)
- Especímenes miden entre 1,01-1,52 mm de longitud; botones alares se extienden anteriormente hasta tocar la parte posterior del ojo o un tercio de los ojos y posteriormente hasta el tercer segmento abdominal; parte media distal de las antenas frecuentemente oscuras (Fig. 1E): .................. Ninfa IV (N4)
Esta clave requiere de la ayuda de un estéreo-microscopio. El rango de las medidas de la clave para la diferenciación de los estados de desarrollo de D. citri están basadas en mediciones del presente estudio y en las reportadas por Fonseca et al. (2007) y Tsai y Liu (2000).
Conclusiones
El ciclo de vida de D. citri tuvo una duración promedio de 15 días desde huevo a adulto bajo condiciones de invernadero (temperatura máxima: 37,1 ± 5,1 °C, temperatura mínima: 25,0 ± 2,0 °C; humedad relativa máxima: 67,0 ± 6,2%, humedad relativa mínima: 39,5 ± 10,9%; 12: 12 h Luz: Oscuridad, régimen natural de luz). Solamente el 7% de las posturas de D. citri llegó al estado adulto, presentándose la mayor mortalidad en los estadios de ninfa I (81%). Las hembras presentaron posturas promedio de 237 huevos durante su ciclo reproductivo. Los parámetros poblacionales sugieren que si bien solo 8 hembras reemplazan a cada hembra viva de D. citri, el hecho de poder presentar un potencial de 16 generaciones al año, hace que su nivel de daño sea alto, con el agravante de que un nivel bajo poblacional no deja de ser preocupante para un insecto que sirve como vector del HLB, enfermedad catastrófica para los cultivos de cítricos. Es necesario enfocar la atención en la combinación efectiva de tácticas de manejo que permitan la reducción de poblaciones de D. citri y a la vez promover estudios en la reducción de daño de la bacteria causante del HLB.
Agradecimientos
Al Ministerio de Agricultura y Desarrollo Rural (MADR) por la financiación del proyecto "Evaluación de entomopatógenos y productos químicos para el control de Diaphorina citri Kuwayama (Hemiptera: Liviidae) y estudios biológicos de D. citri y su enemigo natural principal Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae)", convenio 1828; desarrollado durante el 24 de enero hasta el 31 de diciembre de 2014. A los revisores anónimos por la crítica revisión del manuscrito.
Literatura citada
ANDREWARTHA, H. G.; BIRCH, L. C. 1954. The Distribution and Abundance of Animals. Chicago. University of Chicago. [ Links ]
ÁNGEL, J. E.; HERNÁNDEZ, E. G.; HERRERA, N. A.; GÓMEZ, L. Y.; CASTRO, A. P.; SEPÚLVEDA, A. M.; EBRATT, E. E. 2014. Citrus huanglongbing: validation of Real-Time PCR (qPCR) for the detection of Candidatus Liberibacter asiaticus and Candidatus Liberibacter americanus in Colombia. Agronomía Colombiana 32 (3): 377-389. [ Links ]
CATLING, H. D. 1970. Distribution of psyllid vectors of citrus greening disease with notes on the biology and bionomics of Diaphorina citri. Food and Agriculture Organization, Plant Protection Bulletin 18: 8-15. [ Links ]
COSTA LIMA, A. M. da. 1942. Homopteros. Insetos do Brazil. Escuela Nacional De Agronomía. Ministerio de Agricultura 3: 1-327. [ Links ]
EBRATT-RAVELO, E. E.; RUBIO-GONZÁLEZ, L. T.; COSTA, E. M.; ZAMBRANO GÓMEZ, V. A.; CASTRO-ÁVILA, A. P.; SANTAMARÍA GALINDO, M. Y. 2011a. Primer registro de Tamarixia radiata (Waterston, 1922) (Hymenoptera: Eulophidae) en Colombia. Revista Facultad Nacional de Agronomía Medellín 64 (2): 6141-6146. [ Links ]
EBRATT-RAVELO, E. E.; RUBIO-GONZÁLEZ, L. T.; COSTA, V. A.; CASTRO-ÁVILA, A. P.; ZAMBRANO-GÓMEZ, E. M.; ÁNGEL-DÍAZ, J. E. 2011b. Diaphorina citri (Kuwayama, 1907) and Tamarixia radiata (Waterston, 1922) in citrus crops of Cundinamarca, Colombia. Agronomía Colombiana 29 (3): 487-493. [ Links ]
ÉTIENNE, J.; QUILICI, S.; MARIVAL, D.; FRANCK, A. 2001. Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits 56: 307-315. [ Links ]
FONSECA, O.; VALERA, N.; VASQUEZ, C. 2007. Registro y ciclo de vida de Diaphorina citri Kuwayama (Hemiptera: Psyllidae) en tres hospederos en el estado Lara, Venezuela. Entomotrópica 22 (3): 145-152. [ Links ]
GONZÁLEZ, C.; HERNÁNDEZ, D.; CABRERA, R. I.; TAPIA, J. R. 2002. Diaphorina citri Kuw., inventario y comportamiento de los enemigos naturales en la citricultura cubana. Resúmenes de las sesiones orales. Instituto de Investigaciones de Cítricos y otros Frutales. La Habana, Cuba. 10 p. [ Links ]
HALBERT, S. E.; NUÑEZ, C. A. 2004. Distribution of the Asian citrus psyllid, Diaphorina citri Kuwayama (Rhynchota: Psyllidae), in the Caribbean basin. Florida Entomologist 87 (3): 401-402. [ Links ]
HALL, D. G.; HENTZ, M. G. 2011. Seasonal flight activity by the Asian citrus psyllid in east central Florida. Entomologia Experimentalis et Applicata 139: 75-85. [ Links ]
ICA [Instituto Colombiano Agropecuario]. 2010. Situación actual de HLB (Huanglonbing) y su vector el psilido asiático de los cítricos (Diaphorina citri Kuwayama) en Colombia. Boletín Epidemiológico. 9 p. [ Links ]
ICA [Instituto Colombiano Agropecuario]. 2015. Declaración el estado de emergencia fitosanitaria en el territorio nacional por la presencia de adultos de Diaphorina citri infectados con la bacteria de la enfermedad del HLB. 10 de diciembre, 2015. Resolución No. 00002390. 7 p. [ Links ]
KONDO, T.; PALACINO-RODRÍGUEZ, F.; PEÑA-CUELLAR, R. D. 2015a. Report of Erpetogomphus sabaleticus Williamson, 1918 (Odonata: Gomphidae) feeding on Diaphorina citri Kuwayama (Hemiptera: Liviidae). Boletín del Museo de Entomología de la Universidad del Valle 16 (1): 17-26. [ Links ]
KONDO, T.; GONZÁLEZ F., G.; TAUBER, C.; GUZMÁN-SARMIENTO, Y.C.; VINASCO-MONDRAGON, A.F.; FORERO, D. 2015b. A checklist of natural enemies of Diaphorina citri Kuwayama (Hemiptera: Liviidae) in the department of Valle del Cauca, Colombia and the World. Insecta Mundi 0457: 1-14. [ Links ]
KONDO, T.; QUINTERO, E. M.; CAMPUZANO, M.; WYCKHUYS, K. A. G.; HERATY, J. 2012. First report of Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae), a parasitoid of the Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae) in the department of Valle del Cauca, Colombia. Boletín del Museo de Entomología de la Universidad del Valle 13 (1): 48-51. [ Links ]
KONDO, T.; SIMBAQUEBA CORTÉS, R. 2014. Sarucallis kahawaluokalani (Kirkaldy) (Hemiptera: Aphididae), a new invasive aphid on San Andres island and mainland Colombia, with notes on other recent invasive species. Insecta Mundi 0362: 1-10. [ Links ]
LIU, Y. H.; TSAI, J. H. 2000. Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Annals of Applied Biology 137 (3): 201-206. [ Links ]
MAIA A. DE H. N.; LUIZ, A. J. B.; CAMPANHOLA, C. 2000. Statistical inference on associated fertility life table parameters using Jackknife technique: Computational aspects. Journal of Economic Entomology 93: 511-518. [ Links ]
MANJUNATH, K. L.; HALBERT, S. E.; RAMADUGU, C.; WEBB, S.; LEE, R. F. 2008. Detection of 'Candidatus Liberibacter asiaticus' in Diaphorina citri and its importance in the management of citrus Huanglongbing in Florida. Phytopathology 98 (4): 387-396. [ Links ]
MANJUNATH, K. L.; RAMADUGU, C.; CASTANEDA, A.; DIAZ, J. E.; PEÑARANDA, E. A.; CHEN, J.; DUAN, Y. P.; HALBERT, S. E.; LEE, R. F. 2015. Report of Candidatus Liberibacter caribbeanus, a new citrus- and psyllid-associated Liberibacter from Colombia, South America. The American Phytopathological Society. Proceedings of the APS Annual Meeting. Agosto 1-5, 2015. Pasadena, California, EE.UU. 1 p. [ Links ]
MEAD, F. W.; FASULO, T. R. 2010. Asian citrus psyllid, Diaphorina citri Kuwayama (Insecta: Hemiptera: Psyllidae). FDACS/DPI Entomology. Circular 180. University of Florida, Gainesville, FL. [ Links ]
MORALES, P.; FONSECA, O.; NOGUERA, Y.; CABAÑA, W.; RAMOS, F.; ESCALONA, E.; SANDOVAL, E. 2010. Evaluacion del ciclo de vida del psilido asiático de los citricos en cinco plantas hospederas. Agronomia Tropical 60 (3): 283-286. [ Links ]
NAVA, D. E.; TORRES, M. L. G.; RODRIGUEZ, M. D. L.; BENTO, J. M. S.; PARRA, J. R. P. 2007. Biology of Diaphorina citri (Hem: Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology 131 (9-10): 709-715. [ Links ]
RABINOVICH, J. E. 1978. Ecología de las Poblaciones Animales. Washington: Secretaría General de la Organización de los Estados Americanos. [ Links ]
RABINOVICH, J. E. 1980. Introducción a la Ecología de Poblaciones Animales. Compañía Editorial Continental S.A., México, D.F. [ Links ]
RODRÍGUEZ-PALOMERA, M.; CAMBERO-CAMPOS, J., ROBLES-BERMÚDEZ, A.; CARVAJAL-CAZOLA C.; ESTRADA-VIRGEN, O. 2012. Natural enemies associated to Diaphorina citri Kuwayama (Hemiptera: Psyllidae) in Citrus latifolia Tanaka, in the State of Nayarit, Mexico. Acta Zoológica Mexicana (n.s.) 28: 625-629. [ Links ]
SAS INSTITUTE INC. 2012. SAS procedures guide, versión 9.3. SAS Institute Inc., Cary, NC. [ Links ]
SILVEIRA-NETO, S.; NAKANO, O.; BARBIN. D. 1976. Manual de Ecología dos Insetos. São Paulo: Ceres. [ Links ]
SOUTHWOOD, T. R. E. 1978. Ecological Methods with Particular Reference to the Study of Insects Populations. 2nd. edition. Londres: Chapman and Hall. [ Links ]
TSAI, J. H.; LIU, Y. H. 2000. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. Journal of Economic Entomology 93 (6): 1721-1725. [ Links ]
WOOLER, A.; PADGHAM, D.; ARAFAT, A. 1974. Outbreaks and new records. Saudi Arabia. Diaphorina citri on citrus. FAO Plant Protection Bulletin 22: 93-94. [ Links ]
Recibido: 09-sep-2015
Aceptado: 31-ene-2016

