










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.42 no.1 Bogotá Jan.,/June 2016
SECCIÓN BÁSICA / BASIC
ARTÍCULOS DE INVESTIGACIÓN / RESEARCH PAPER
Composición y estructura de estafilínidos (Coleoptera: Staphylinidae) en dos localidades de Santa María (Boyacá, Colombia)
Composition and structure of rove beetles (Coleoptera: Staphylinidae) in two localities from Santa María (Boyaca, Colombia)
Yudy Paulina Sissa-DueñasI; José Luis Navarrete-HerediaII
IBióloga. Universidad Pedagógica y Tecnológica de Colombia. Laboratorio de Entomología, Museo de Historia Natural "Luis Gonzalo Andrade". Avenida Central del Norte 39-115. Tunja, Colombia, megalopinus0926@gmail.com, autor para correspondencia
IIDr. en Ciencias (Biología). Profesor Investigador Titular C. Centro de Estudios en Zoología, Entomología, CUCBA. Universidad de Guadalajara. Apdo. postal 134, 45100. Zapopan, Jalisco, México. glenusmx@gmail.com
RESUMEN
Los bosques húmedos tropicales albergan una gran riqueza y diversidad de especies de estafilínidos. Sin embargo, en Colombia el conocimiento de esta diversidad es escaso y más en ecosistemas sobre la Cordillera Oriental. Este estudio registra la composición y estructura de los coleópteros estafilínidos en dos localidades del municipio de Santa María, en el departamento de Boyacá. Se utilizaron como métodos de colecta transectos compuestos por trampas de caída, tipo "pitfall", con diferentes cebos o atrayentes, además de cernido de hojarasca y colecta manual en un gradiente altitudinal entre los 1.000 y 1.400 msnm. En total se recolectaron 1.378 especímenes, agrupados en 179 morfoespecies, 56 géneros y 11 subfamilias, donde la subfamilia Aleocharinae registró gran parte de la abundancia (58,3%) mientras que en términos de riqueza Aleocharinae (32%), Staphylininae (17%) y Pselaphinae (16%) se encuentran dentro de las subfamilias mejor representadas. Los altos valores de diversidad en la Almenara y el Sendero Ecológico "Hiya Quye", contribuyen con la diversidad regional, además se encontraron diferencias significativas en términos de abundancia entre las dos localidades estudiadas. Las tasas de recambio de especies fueron altas, con una gran variación en la composición de estafilínidos a través del gradiente altitudinal en las localidades estudiadas, que se refleja en los bajos índices de similitud (entre 0,10 y 0,27). Géneros como Belonuchus, Philothalpus, Aleochara, Hoplandria, Atheta, Prionopedinus, Anotylus, Coproporus, Holotrochus, Trimicerus se encontraron distribuidos a lo largo del gradiente altitudinal estudiado.
Palabras clave: Bosque húmedos tropical, Diversidad, Gradiente altitudinal, Recambio de especies.
ABSTRACT
Tropical rain forests contain a high species richness and diversity of rove beetles. However, in Colombia in general, knowledge of this diversity is scarce; and even more so in ecosystems on the Eastern Range. This study records the composition and structure of rove beetles in two municipalities of Santa Maria country, in the department of Boyacá. Pitfall traps baited with different organic matter, as well as leaf litter siftings and manual collection in an altitudinal gradient between 1,000 and 1,400 m.a.s.l were used to evaluate the species richness and diversity of these beetles. In total, 1,378 beetles were collected which were grouped into 179 morphospecies, 56 genera, and 11 subfamilies. Aleocharinae was the most abundant (58.3%), while, by overall species richness, Aleocharinae (32%), Staphylininae (17%), and Pselaphinae (16%) were the best represented. High levels of diversity in La Almenara and the "Hiya Quye" ecological trail contribute to regional diversity; however, significant differences in abundance were found between the two localities studied. Species turnover rates were high, with large variations in the composition of rove beetles throughout the altitudinal gradient in both localities studied reflected in a low index of similarity (between 0.17 and 0.27). Genera such as Belonuchus, Philothalpus, Aleochara, Hoplandria, Atheta, Prionopedinus, Anotylus, Coproporus, Holotrochus, and Trimicerus were found in the altitudinal gradient studied.
Key words: Tropical rainforest. Diversity. Altitudinal gradient. Species turnover.
Introducción
Los escarabajos pertenecen al orden Coleoptera y constituyen el 40% de la diversidad de insectos (Grimaldi y Engel 2005) con 392.415 especies (Zhang 2013). La familia Staphylinidae Latreille, 1802, constituye el grupo más diverso de este orden y de todo el reino animal a ese nivel (Grebennikov y Newton 2009) con 56.000 especies agrupadas en 3.400 géneros (Newton et al. 2005; Ślipiński et al. 2011). Esta diversidad se ve reflejada en la gran variedad de hábitats y hábitos en los que se pueden encontrar, especialmente aquellos con un alto contenido de humedad (García et al. 2001; Navarrete-Heredia et al. 2002).
La hojarasca es un hábitat ideal, en el que se encuentran representantes de todas las subfamilias y donde muchas especies utilizan como alimento materia orgánica en descomposición (Navarrete- Heredia et al. 2002) y otras son depredadores de los artrópodos del suelo (Boháč 1999). Los troncos en descomposición, hongos, la carroña y el excremento de diferente origen animal son también un hábitat ideal para los estafilínidos; además se pueden encontrar asociados a cuerpos de agua (Navarrete-Heredia et al. 2002), flores (López-García et al. 2011), roedores (Ashe y Timm 1987), aves y en nidos o detritos de hormigas y termitas (Boháč 1999; Navarrete-Heredia 2001). Algunas especies del género Paederus son de importancia médica para el ser humano ya que ocasionan una enfermedad conocida como dermatitis linearis con daños en la piel tipo eritemas leves o daños severos (Frank y Kanamitsu 1987).
A pesar del poco conocimiento de Staphylinidae en Sur América, en los últimos años se han realizado trabajos significativos (Asenjo 2011) entre los que se pueden mencionar: la lista preliminar de los estafilínidos para Perú (Asenjo 2004), la lista anotada de estafilínidos de Colombia (Newton et al. 2005) y el más reciente, la lista anotada de estafilínidos de Brasil con nuevos registros y distribución de especies (Asenjo et al. 2013). Además de los trabajos sobre diversidad a nivel de país, se han publicado trabajos taxonómicos recientes, entre los que se tienen: revisiones de algunos géneros de la subtribu Xanthopygina (Chatzimanolis 2012; 2013), subtribu Philonthina (Chani-Posse 2011, 2013; Chani-posse y Asenjo 2013), Tribu Leptochirini (Asenjo y Ribeiro-Costa 2013), entre otros.
En Colombia la diversidad de estafilínidos registra 796 especies y 230 géneros, pero se calcula que esta cifra puede estar cerca de 5.000 especies, teniendo en cuenta las muchas especies sin describir observadas en colecciones y nuevas adicionadas en trabajos posteriores (Newton et al. 2005). Algunos de los trabajos ecológicos registran la diversidad de estafilínidos en bosques secos (García et al. 2001; García y Chacón 2005), ecosistemas subandinos y altoandinos con diferentes grados de intervención (Sanabria et al. 2008; Méndez-Rojas et al. 2009; Vásquez-Vélez et al. 2010; Méndez-Rojas et al. 2012) y en gradientes altitudinales que abarcan bosques amazónicos de tierras bajas, subandinos y bosques andinos, en donde se encuentra una gran diversidad y riqueza de estafilínidos para la Cordillera Oriental con altas tasas de recambio de especies (Gutiérrez-Chacón y Ulloa-Chacón 2006). Con un enfoque más taxonómico, recientemente se realizó una sinopsis de la subtribu Xanthopygina para el país (Torres et al. 2012) y la descripción de nuevas especies (López-García et al. 2011; López-García y Méndez-Rojas 2014). A pesar de ello, aún falta conocer mucha de la fauna de Staphylinidae en el país, especialmente en ecosistemas de los andes orientales, que sumado a los "impedimentos taxonómicos", ocasionalmente desalientan el interés para el trabajo con este grupo.
Los bosques de Santa María (Boyacá), los cuales albergan una gran riqueza de flora y fauna, han sido de un especial interés en estudios de diversidad, como lo expresa la empresa Aes Chivor en su compromiso de cuidar, conservar y velar por la preservación de las 1.700 hectáreas de bosques (Aguirre-Ceballos 2011); los cuales cuentan con una gran riqueza y abundancia de insectos como hemípteros, coleópteros, himenópteros (Amat-García 2009) y mariposas con 146 especies registradas para esta zona (Andrade-C et al. 2007). Debido al escaso conocimiento de la familia Staphylinidae en el país, y en especial en ecosistemas naturales tan importantes como son los bosques húmedos tropicales y muy húmedos premontanos, en este estudio se determinó la composición y estructura de los estafilínidos en dos localidades de Santa María.
Materiales y métodos
Área de estudio. Ubicado en la región Centro Oriente de Colombia, sobre la Cordillera Oriental del Macizo Andino se encuentra el municipio de Santa María a 04°51´48"N y 73°16´04"O (Fig. 1), con temperatura media de 24 ºC y precipitación anual de 4.282 mm (Fernández-Alonso 2009).
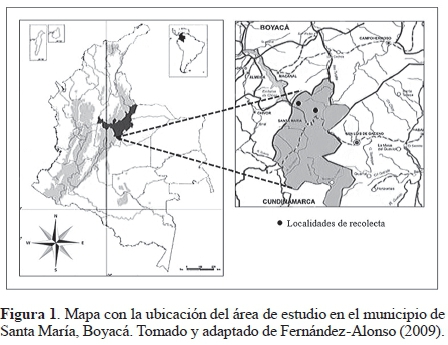
El estudio se desarrolló dentro de la reserva privada de la empresa Aes Chicor, que comprende bosque húmedo tropical y muy húmedo premontano (Fernández-Alonso 2009), con altitudes entre los 800 y 2.000 msnm. Allí se escogieron dos localidades representativas, las cuales se seleccionaron teniendo en cueta su estado de conservación: (1) La Almenara, bosques secundarios, con árboles de entre 10-20 m de dosel, con gran cantidad de helechos y pocas hierbas y arbustos en el sotobosque. Estos bosques se encuentran junto a quebrada y a una matriz de pastizal y (2) Sendero Ecológico "Hiya Quye", bosques secundarios, con árboles de entre 15-30 m de dosel, sotobosque con hierbas y arbustos más densos y exuberantes, con un alto contenido de hojarasca, con presencia de troncos caídos naturalmente en estado de descomposición en los cuales crecen algunos hongos. Estos bosques se ubican junto a cuerpos de agua (quebradas y cascadas) y las precipitaciones son más frecuentes en esta zona.
Diseño de colecta. Se realizó un muestreo de 10 días en marzo de 2012 en la localidad La Almenara y en septiembre de 2012 en la localidad del Sendero Ecológico "Hiya Quye", implementando en cada localidad el siguiente diseño:
Se trazaron dos transectos en cada gradiente altitudinal (1.000, 1.200 y 1.400 msnm). En cada transecto se colocaron cinco estaciones de muestreo separadas cada 20 m, cada estación constaba de tres trampas de caída, las cuales poseían un tipo de cebo (fruta en descomposición, calamar en descomposición y sin cebo) separadas 5 m entre sí. La trampa de caída consiste en un vaso plástico de 16 onzas de capacidad enterrado a ras de piso el cual contiene como agente preservante etilenglicol y etanol 70% en proporción 1:1. En total se instalaron 90 trampas distribuidas en los seis transectos y se dejaron actuar durante los diez días del muestreo.
Para la colecta activa se empleó golpeteo arbustivo de 8:00 a 10:00 am y búsqueda activa de 10:00 a 12:00 pm en cada una de las altitudes. Se realizó cernido de 2 m2 de hojarasca paralelo a cada estación del transecto de trampas de caída que posteriormente se procesó en embudo de Berlesse por cuatro días (60 m2 de hojarasca procesada en cada localidad y 10 m2 de hojarasca procesada por transecto), para un total de 210 unidades muestrales.
Determinación taxonómica. Los estafilínidos colectados se separaron e identificaron hasta nivel taxonómico posible, utilizando claves taxonómicas (Navarrete-Heredia et al. 2002) y revisiones taxonómicas de algunas tribus, subtribus y géneros. Además se contó con la colaboración de los especialistas de diferentes instituciones y especialidad taxonómica: Ph. D. Ulrich Ilmler (University of Kiel) y Ph. D. Angélico Asenjo (Universidade Federal de Mato Grosso) para el caso de la subfamilia Osoriinae, Ph. D. Joseph Parker (Columbia University) y Ph. D. Donald S. Chandler (University of New Hampshire) para Pselaphinae, Ph. D. Margaret K. Thayer (Field Museum of Natural History) para Paederinae y Ph. D. Lee H. Herman (American Museum of Natural History) en el caso de la subfamilia Oxytelinae; para confirmar y complementar la determinación de los ejemplares colectados.
Debido a las dificultades taxonómicas para identificar hasta el nivel de especie, se reconocieron morfoespecies teniendo en cuenta caracteres morfológicos externos y del genital del macho (depositada en microviales con glicerina), así que se utilizó el término de "especies" al referirnos a las morfoespecies diferenciadas. Una vez etiquetado, preservado y montado los ejemplares se incluyeron en la colección entomológica de la Universidad Pedagógica y Tecnológica de Colombia.
Análisis de datos. Para inspeccionar y evaluar la calidad del inventario de Staphylinidae de Santa María y en cada localidad con los diferentes métodos de colecta se calculó el promedio estadístico de adición de especies con el aumento del esfuerzo de muestreo para construir la curva de acumulación y de recolección acumulada de especies (Colwell et al. 2004), además se obtuvó una aproximación al número estimado de especies utilizando los estimadores no paramétricos Chao 1 e ICE (StimateS v 9.1.0; Colwell 2013). Para describir la distribución de especie-abundancia en la muestra y en cada localidad se construyeron curvas rango-abundancia con el log abundancia por especie (RStudio v 0.97.55; R v 3.0.1; Magurran 2004). Para el análisis de la diversidad se calculó el índice de diversidad de Shannon-Wienner (H`) y Equitatividad de Pielou para cada localidad (StimateS v 9.1.0; Colwell 2013) y se aplicó una prueba de "t" para comparar la diversidad entre las localidades (Magurran 1989).
Como medida del recambio de especies a través del gradiente altitudinal se calculó el porcentaje de especies halladas en una, dos y tres altitudes (Gutiérrez-Chacón y Ulloa-Chacón 2006) y con el fin de observar la composición de estafilínidos en el gradiente altitudinal se realizó un dendrograma de similitud usando en coeficiente de similitud de Jaccard y un análisis de agrupamiento (Past v 3.04; Hammer et al. 2001). Finalmente, la abundancia y riqueza de estafilínidos por localidad y por método de colecta fueron comparadas aplicando la prueba no paramétrica chi cuadrado.
Resultados
Composición de estafilínidos. En total se colectaron 1.378 estafilínidos, 179 morfoespecies, 56 géneros y 11 subfamilias. Para el caso de la subfamilia Aleocharinae y algunas morfoespecies de Pselaphinae y Paederinae éstas fueron determinadas a nivel de subfamilia y tribu respectivamente, debido a la dificultad taxonómica de estos grupos. Sin embargo se encontraron cinco géneros de Pselaphinae como nuevos registros para el país (Tabla 1).
La subfamilia mejor representada en la muestra de Santa María en cuanto a abundancia corresponde a Aleocharinae con el 58,3% de los estafilínidos colectados, seguida por Staphylininae (11,1%), Pselaphinae (9%), Oxytelinae (6,2%) y Osoriinae (4,6%). Así mismo la subfamilia Aleocharinae (32%) registra la mayor riqueza, seguida de Staphylininae (17%), Pselaphinae (16%) y Osoriinae (10%) las cuales reúnen el 75% de la riqueza de estafilínidos de Santa María; mientras que las subfamilias Megalopsidiinae, Steninae, Piestinae se encontraron con la menor abundancia y riqueza. Por otro lado, tanto en La Almenara y el Sendero Ecológico la subfamilia Aleocharinae registró la mayor proporción de estafilínidos, seguida de Staphylininae. No se registraron especies de Piestinae y Steninae en La Almenara y de Megalopsidiinae en el Sendero ecológico "Hiya Quye".
Diversidad y composición. Los valores de diversidad para ambas localidades son altos (Tabla 2). Al compararlos mediante la prueba de t (t = 3,5, P < 0,001), se observa que los dos bosques son significativamente distintos, situación que se puede argumentar en función de los diferentes grados de intervención y conservación. Los estimadores asintóticos de riqueza (ICE, Chao 1) estiman que la riqueza de estafilínidos es cercana a 275 y 334 especies. Los valores de riqueza representan tan sólo el 60% de las especies de estafilínidos lo que contrasta con la curva de acumulación de especies la cual no forma una asíntota lo que indica que el número de especies se puede incrementar por la adición de especies esporádicas (Fig. 2A), presentando el mismo comportamiento con el cernido de hojarasca como método de colecta en las dos localidades estudiadas (Figs. 2D, E). Por el contrario, se observó que con las trampas de caída en las dos localidades las curvas de acumulación de especies alcanzan una asíntota (Figs. 2B, C), esto sugiere que el esfuerzo de muestreo fue suficiente ya que se tiende a colectar las mismas especies; indicando que se requiere mayor esfuerzo de muestreo con la colecta manual y el cernido de hojarasca para conocer la diversidad real de Santa María. Finalmente, no se encontraron diferencias significativas en cuanto a riqueza (χ 2 = 1,0181, P = 0,313), pero si en términos de abundancia (χ 2 = 42,499, P = 7e-11) entre las localidades estudiadas.
La distribución de la abundancia en Santa María no es equitativa ya que muy pocas especies fueron abundantes, con una débil dominancia y una larga cola de especies raras (Fig. 3A). El 5% del total de los estafilínidos corresponde a 67 morfoespecies ("singletons") con una sola captura en la muestra, el 68% se encuentra distribuido en 109 especies cuyas abundancias se encuentran entre 2 a 66 individuos. Las especies mas abundantes fueron Aleochara sp.1 con el 10 % de los estafilínidos colectados, seguido por Hoplandria sp. 2 (9%) y Lomechusini sp. 10 (8%). Aunque en La Almenara se presentó una mayor pendiente representada por 2 morfoespecies (Hoplandria sp. 2 y Aleochara sp.1), en ambas localidades estudiadas la distribución de la abundancia es similar (Fig. 3B), cuyas morfoespecies dominantes corresponden a la subfamilia Aleocharinae.
Comparación y composición entre métodos de colecta. De las 210 unidades de muestreo 155 fueron efectivas (con el registro de al menos un estafilínido). La mayor abundancia se registró en las trampas de caida con un total de 780 individuos (57% de los estafilínidos colectados) y 92 morfoespecies, mientras que con el cernido de hojarasca se capturaron 320 individuos (78 morfoespecies) y con la colecta manual se capturó 278 individuos (41 morfoespecies), encontrándose diferencias significativas tanto en riqueza (χ 2 = 19,74, P = 5,15975e-05) como abundancia (χ 2 = 337,71; P = 4,64125e-74) entre los métodos de colecta.
Las trampas de caída fueron muy efectivas en la captura principalmente de representantes de las subfamilias Aleocharinae, Staphylininae y Oxytelinae. Sin embargo, con el cernido de hojarasca se recolectaron estafilínidos pertenecientes a las subfamilias Paederinae y Pselaphinae, esta última como la más abundante con el 36% de los individuos colectados en hojarasca. Mientras que las especies de las subfamilias Megalopsidiinae y Steninae se registraron exclusivamente con la colecta manual.
Entre los tipos de cebos utilizados en las trampas de caída o "pitfall", el calamar en descomposición mostró ser el cebo o atrayente más eficaz en la captura de estafilínidos con 548 individuos y 68 mofoespecies colectadas, a diferencia de la fruta en descomposición (160) y las trampas sin cebo (72) que registraron menos de la tercera parte de los estafilínidos colectados con el calamar en descomposición. Por lo tanto, entre los tipos de cebo o atrayectes existen diferencias significativas tanto en riqueza (χ 2 =13,70; P = 0,001) como abundancia (χ 2 = 493,41; P = 7,1814e-108). Siendo el calamar en descomposición muy eficaz en la captura de representantes de los géneros Aleochara, Hoplandria y Philotalpus.
Variación de la composición de especies en el gradiente altitudinal. La composición de estafilínidos entre las localidades estudiadas fue diferente, donde se forman dos grupos bien diferenciados: A y H (que corresponde a cada localidad). El valor más bajo de similitud corresponde a 0,10, con el 76,53% de las morfoespecies exclusivas de cada localidad. La mayor similitud se presentó en gradientes altitudinales variables: 1.000 y 1.200 msnm en el Sendero Ecológico y 1.200 y 1.400 msnm en La Almenara (Fig. 4).
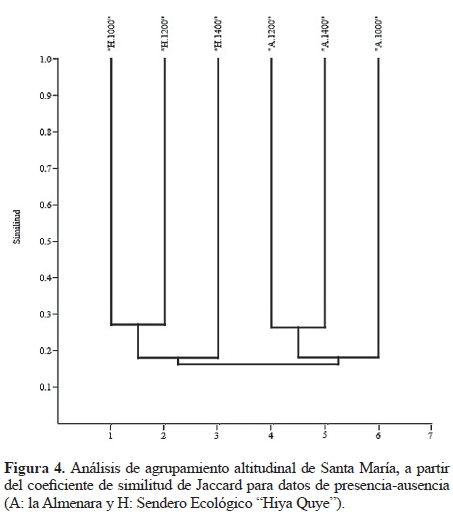
Como medida del recambio de especies a través del gradiente altitudinal se calculó el porcentaje de especies halladas en una sola y/o varias altitudes, siendo esta medida utilizada en un trabajo anterior por otros autores (Gutiérrez-Chacón y Ulloa-Chacón 2006). El 63% de las especies se restringen a un sólo gradiente altitudinal, el 25% de las especies se encuentran en dos altitudes y el 12% en las tres altitudes estudiadas. En las localidades se observó una gran variación en la composición de estafilínidos a través del gradiente altitudinal, que se refleja en los bajos índices de similitud establecidos entre altitudes que se hallan entre 0,10 y 0,27. Las morfoespecies con amplia distribución altitudinal (se registran en las tres altitudes), pertenecen a los géneros Belonuchus, Heterothops, Quedius, Philothalpus, Prionopedinus, Anotylus, Coproporus, Holotrochus, Trimicerus, Aleochara, Hoplandria y Atheta.
Discusión
Composición de estafilínidos. La proporción de la riqueza de especies por subfamilia corresponde con la diversidad de especies dentro de Staphylinidae siendo la subfamilia Aleocharinae la más diversa con aproximadamente 12.851 especies a nivel mundial, seguida de Pselaphinae y Staphylininae con 9.110 y 6.876, respectivamente (Newton et al. 2001; Asenjo et al. 2013), donde estas especies son consideradas depredadores generalistas y dominantes en la fauna del suelo y la hojarasca (Navarrete-Heredia et al. 2002). Aunque la subfamilia Osoriinae aporta un gran número de especies al estudio, esto se debe a que muchas de ellas se encuentran exclusivamente en regiones tropicales (Navarrete-Heredia et al. 2002) y a la implementación en este estudio del método de colecta manual, ya que la mitad de morfoespecies registradas fueron colectadas debajo de la corteza de troncos en descomposición donde son muy frecuentes (Navarrete-Heredia et al. 2002). De las 20 subfamilias registradas para Colombia, en este estudio se reportaron 11, incluyendo la subfamilia Scydmaeninae incorporada recientemente a Staphylinidae (Grebennkov y Newton 2009).
Las subfamilias Megalopsidiinae y Steninae registran el menor número de especies, que coincide con la proporción dentro de Staphylinidae. Sin embargo su registro fue posible mediante su colecta en hábitats específicos, en el caso de Megalopinus sobre un tronco mohoso y Stenus sobre el musgo que crece junto a una cascada o en el material vegetal acumulado entre las rocas de la misma, coincidiendo con el tipo de hábitat que frecuentan estos estafilínidos (Navarrete-Heredia et al. 2002).
Diversidad y composición. Las dos localidades de Santa María presentan una alta diversidad de estafilínidos, pero el Sendero Ecológico registró la mayor diversidad y equitatividad. Esta variación podría estar influenciada por la diferencia en las fechas de muestreo, pues durante el año pueden ocurrir cambios en las condiciones climáticas que afectan la composición de especies de estafilínidos o por el estado de conservación de cada localidad. Esta alta diversidad también ha sido encontrada en bosques tropicales de tierras bajas, bosques subandinos y altoandinos (Gutiérrez-Chacón y Ulloa-Chacón 2006; Méndez et al. 2009; Méndez-Rojas et al. 2012). Por lo tanto, los ecosistemas de los andes colombianos debido a su heterogeneidad paisajística albergan una gran diversidad de estafilínidos, y en especial sobre la cordillera Oriental pese al reducido trabajo ecológico ya que la gran mayoría de estudios se han centrado sobre las cordilleras Occidental y Central.
La Almenara y el Sendero Ecológico son similares en cuanto a riqueza de especies, es probable que su diferencia esté dada por la presencia de morfoespecies diferentes en cada localidad, pues hay un bajo número de morfoespecies compartidas (42) entre estas dos localidades. Sin embargo el Sendero Ecológico registró la mayor abundancia de estafilínidos, lo que puede estar influenciado por la estructura vegetal de la zona y la presencia de cuerpos de agua como quebradas y cascadas que finalmente ayudan a mantener la humedad en los diferentes microhábitats, siendo estos hábitats húmedos ideales para muchos estafilínidos (Navarrete-Heredia et al. 2002).
Por otro lado, se encontró que la abundancia no es equitativa, ya que muy pocas especies fueron muy abundantes y muchas son especies "singletons". Este patrón en la distribución de la abundancia ha sido igualmente encontrado en estudios ecológicos con estafilínidos en el país (García y Chacón 2005; Sanabria et al. 2008; Vásquez-Vélez et al. 2010; Méndez-Rojas et al. 2012), con morfoespecies de los géneros Aleochara, Hoplandria, Atheta y la tribu Lomechusini como las más abundantes. Estos aleocarinos con frecuencia son dominantes y abundantes en muchos microhabitats (Navarrete-Heredia et al. 2002) sin embargo, en este estudio, fueron colectados principalmente con necrotrampas ya que, en condiciones naturales, es la carroña el hábitat donde se desarrollan larvas de dípteros, presas potenciales de estos estafilínidos (Navarrete-Heredia et al. 2002) y donde algunas especies del género Aleochara se han observado depredando larvas de dípteros (Klimaszewski 1984; Maus et al. 1998).
Con un grupo tan hiperdiverso como Staphylinidae el cual es considerado como el más diverso de Coleoptera (Grebennkov y Newton 2009), los inventarios son raramente completos (Jiménez-Valverde y Hortal 2003), y en especial cuando se encontró en este estudio un gran número de especies "singletons", lo cual es común en zonas tropicales (Gotelli y Colwell 2011). Lo que hace que conseguir un inventario representativo de estafilínidos sea complejo, pese a que se muestreó gran variedad de hábitat utilizando métodos eficientes para la captura de estos insectos como las trampas de caída y el cernido de hojarasca (Gutiérrez-Chacón y Ulloa-Chacón 2006; Méndez-Rojas et al. 2009) además de complementarios como la colecta manual.
Comparación y composición entre métodos de colecta. Debido a la necesidad de conocer la verdadera riqueza de estafilínidos se implementaron diferentes métodos de colecta. De todos ellos las trampas de caída "pitfall" resultaron ser las más eficaces en la captura, información resaltada también por Méndez-Rojas et al. (2009), quienes utilizaron este tipo de trampas cebadas con pescado en descomposición como atrayente. Mientras que en trabajos donde no se utilizó ningún tipo de atrayente el método Winkler fue el más eficaz en la captura de estafilínidos comparado con las trampas de caída (García et al. 2001; Gutiérrez-Chacón y Ulloa-Chacón 2006). Por lo tanto, la utilización de cebos o atrayentes aumenta la efectividad de captura de estafilínidos, lo cual explica el bajo número de individuos (72) colectados con las trampas de caída sin cebo.
El calamar en descomposición fue un cebo muy útil para la colecta de estafilínidos. En él se registró cerca de la mitad de las colectas en el muestreo y 64 morfoespecies, con una fracción de géneros de hábitos saprófagos y depredadores como Aleochara, Hoplandria, Atheta, Croaptomus, Belonuchus (Navarrete-Heredia et al. 2002; Frank y Barrera 2010) e individuos del género Philotalpus. Jiménez-Sánchez et al. (2011) también colectaron un gran número de individuos (5.390 en 62 especies) en trampas de caída compuestas con calamar en descomposición, aunque en un periodo de muestreo más largo. Sin embargo, con las trampas de caída se tiende a colectar especies de mayor tamaño (Olson 1994), mientras que con el cernido de hojarasca no sólo se colectan especies de menor tamaño sino también especies hipogeas y con limitada movilidad. Con un gran número de morfoespecies, el cernido de hojarasca es un método muy apropiado en este tipo de estudios de composición, el cual ha sido resaltado como eficaz en la captura de estafilínidos (García et al. 2001; Gutiérrez-Chacón y Ulloa-Chacón 2006) y es justamente con el cernido de hojarasca donde en el presente estudio se encuentra una gran riqueza de morfoespecies y la mitad de las morfoespecies raras o "singlentons", la mayoría de la subfamilia Pselaphinae. Para este grupo tan diverso el cernido de hojarasca resulta ser un método muy apropiado para su colecta (Méndez et al. 2009). De este modo, se puede considerar esta subfamilia como uno de los componentes principales de la fauna de hojarasca o por lo menos en las dos localidades estudiadas, cuyos miembros son considerados depredadores de colémbolos, ácaros y sínfilos (Chandler 2001), y esta alta abundancia y riqueza de pselafínos en la hojarasca es sustancial para el control de las poblaciones de estos artrópodos.
Debido a que los miembros de la familia Staphylinidae cuentan con una amplia gama tanto de hábitats como hábitos, con la colecta manual fue posible registrar subfamilias como Steninae, Megalopsidiinae y algunos géneros de la subfamilia Osoriinae los cuales se encontraron en hábitat específicos como musgo, troncos mohoso y debajo de la corteza de troncos en descomposición; los cuales son hábitats ideales para estos estafilínidos (Navarrete-Heredia et al. 2002; Betz 2002).
A pesar de que se intentó muestrear la mayoría de microhábitats posibles utilizando diferentes métodos de colecta y cebo o atrayentes, las especies esperadas son casi el doble de las observadas. Esto puede atribuirse a que falta mayor esfuerzo de muestreo por parte del cernido de hojarasca, hábitat que alberga una gran riqueza de estafilínidos y la colecta manual en hábitats más específicos.
Variación en la composición de especies en el gradiente altitudinal. Las altas tasas de recambio de especies a través del gradiente altitudinal encontradas en este estudio se refleja en el alto porcentaje de especies halladas en una sola altitud 63% y en los bajos índices de similitud entre las altitudes, en un rango altitudinal estrecho entre los 1.000 y 1.400 msnm. Estas altas tasas de recambio también han sido encontradas sobre la Cordillera Oriental entre los 700 a 2.500 msnm, y donde los miembros de la familia Staphylinidae presentan estrechos rangos de distribución altitudinal (Gutiérrez-Chacón y Ulloa-Chacón 2006), lo que puede ser válido para los estafilínidos en los bosques de Santa María también sobre la cordillera Oriental. Esta gran variación en la composición de estafilínidos en un rango altitudinal estrecho entre los 1.000 a 1.400 msnm puede deberse a que los bosques de Santa María son una transición entre los bosques húmedos tropicales y los bosque nublados subandinos (Fernández-Alonso 2009). Por lo tanto, la variación en la temperatura, humedad y cobertura vegetal hace que se generen gran variedad de hábitats. Por otro lado debe tenerse en cuenta un rango altitudinal más amplio en posteriores trabajos ya que estos bosques presentan influencia tanto de la fauna de la Orinoquia como la fauna subandina.
En otros estudios sobre la Cordillera Oriental, uno de ellos con escarabajos coprófagos, Escobar et al. (2005) encuentran dos grupos de especies uno en altitudes entre 1.750-2.250 msnm y el otro en menores a 1.750 msnm. Por otra parte, Carrero et al. (2013) también registran dos grupos de mariposas uno de 1.200 a 1.600 msnm de altitud y el segundo de 1.800 a 2.000 msnm al comparar la vegetación, encontrando la misma configuración que con la comunidad de mariposas. Para el caso hormigas y mariposas, Fagua (1999) define dos comunidades, una de tierras bajas y la de zonas montanas, al igual que Guerrero y Sarmiento (2010) quienes también definen dos grupos de hormigas, el primero entre 835 y 968 msnm y el otro entre los 1.198 y 1.395 msnm. Por otro lado, aunque Gutiérrez-Chacón y Ulloa-Chacón (2006) no incluyen datos de zonas bajas también encuentran dos grupos de estafilínidos, uno sobre los 1.450 msnm y otro por debajo de este nivel. Sin embargo, en el presente trabajo no se observó este patrón, esto puede deberse en gran medida al bajo número de altitudes estudiadas (3 altitudes) y al estrecho rango altitudinal tenido en cuenta, aunque al observar esta composición de especies a través del gradiente altitudinal es transcendental para comprender los procesos que generan y mantienen la biodiversidad (Hodkinson 2005).
Las especies pertenecientes a los géneros Belonuchus, Philothalpus, Aleochara, Hoplandria, Atheta y Prionopedinus presentaron una amplia distribución a través del gradiente altitudinal estudiado, esto se debe a que muchos de los estafilínidos poseen sus alas bien desarrolladas y efectivamente vuelan (Newton 1990) lo que les permite posiblemente una mayor dispersión a través del gradiente; además algunas especies de los géneros mencionados son consideradas depredadoras presentes frecuentemente en carroña, excremento, fruta en descomposición (Navarrete-Heredia et al. 2002), por lo que se verían principalmente atraídos. Los géneros Anotylus, Coproporus, Holotrochus, Trimicerus, también se encontraron distribuidos en todo el gradiente altitudinal estudiado, sin embargo, sus especies se encuentran más asociados a la hojarasca. Tanto Anotylus como Coproporus son saprófagos y habitan varios ambientes como bosques secundarios, plantaciones, pasturas y bosque seco (Navarrete-Heredia et al. 2002; García y Chacón 2005; Sanabria et al. 2008; Méndez-Rojas et al. 2009; Vásquez-Vélez et al. 2010), es posible que la capacidad de estos géneros para ocupar múltiples ambientes les permita también una mayor distribución a través del gradiente altitudinal donde se presentan cambios en la temperatura, humedad y la vegetación.
Agradecimientos
Los autores queremos agradecer a la empresa AES CHIVOR, Angel A. Ciro (AES Chivor), Eibar Algarra y al Museo de Historia Natural "Luis Gonzalo Andrade" de la Universidad Pedagógica y Tecnológica de Colombia por el apoyo y logística. A David Rodríguez, Ximena Galindo, Willan Sissa y Nevardo Sissa por su colaboración en el trabajo de campo. A Daniel Silva, Margarita López y Diana Torres por sus sugerencias y aportes al trabajo. A los especialistas Angélico Asenjo, Ulrich Ilmler, Joseph Parker, Margaret K. Thayer, Lee H. Herman, y Donald S. Chandler, por su colaboración en la determinación y confirmación taxonómica.
Literatura citada
AGUIRRE-CEBALLOS, J. 2011. Guía de campo de los mamíferos, anfibios, y reptiles de Santa María (Boyacá, Colombia). Series de guías de campo. Instituto Nacional de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. 260 p. [ Links ]
AMAT-GARCÍA, G. 2009. Biodiversidad regional: Santa María, Boyacá. Artrópodos, Miriápodos, Crustáceos, Insectos. Serie de guías de campo. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. 236 p. [ Links ]
ANDRADE-C, M. G.; CAMPOS-SALAZAR, L. R.; GONZÁLEZ-MONTAÑA, L. A.; PULIDO-B, H. W. 2007. Santa María mariposas alas y color. Serie de guías de campo, Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. 248 p. [ Links ]
ASENJO, A. 2004. Lista preliminar de las especies de Staphylinidae (Coleoptera) registradas para Perú. Revista Peruana de Entomología 44: 55-64. [ Links ]
ASENJO, A. 2011. Catálogo documentado de los Staphylinidae presentes en América del Sur. Versión electrónica 2.0. Laboratorio de Sistemática e Biotecnología de Coleoptera (Insecta). Disponible a través del autor: pukara8@yahoo.com. [ Links ]
ASENJO, A.; RIBEIRO-COSTA, C. S. 2013. Revision of the neotropical subgenus Tropiochirus of the genus Leptochirus Germar 1824 (Coleoptera: Staphylinidae: Osoriinae). Journal of Natural History 47 (19-20): 1257-1285. [ Links ]
ASENJO, A.; IRMLER, U.; KLIMASZEWSKI, J.; HERMAN, L. H.; CHANDLER, D. S. 2013. A complete checklist with new records and geographical distribution of the rove beetles (Coleoptera, Staphylinidae) of Brazil. Insecta Mundi 0277: 1-419. [ Links ]
ASHE, J. S.; TIMM, R. M. 1987. Predation by and activity patterns of "parasitic" beetles of the genus Amblyopinus (Coleoptera: Staphylinidae). Journal of Zoology (London) 212 (3): 429-437. [ Links ]
BETZ, O. 2002. Performance and adaptive value of tarsal morphology in rove beetles of the genus Stenus (Coleoptera, Staphylinidae). The Journal of Experimental Biology 205: 1097-1113. [ Links ]
BOHAC, J. 1999. Staphylinidae beetles as bioindicators. Agriculture, Ecosystems and Environment 74: 357-372. [ Links ]
CARRERO, D. A.; SÁNCHEZ, L. R.; TOBAR, D. E. 2013. Diversidad y distribución de mariposas diurnas en un gradiente altitudinal en la región nororiental andina de Colombia. Boletín Científico Centro de Museos, Museo de Historia Natural 17 (1): 168-188. [ Links ]
CHANDLER, D. S. 2001. Biology, morphology, and systematics of the ant-like litter beetle of Australia (Coleoptera: Staphylinidae: Pselaphinae). En: Memoirs on Entomology, International x, 1-562 p. [ Links ]
CHANI-POSSE, M. 2011. Redescription of some South American species of Belonuchus (Coleoptera: Staphylinidae), with a checklist of the southern South America species of the genus. Revista de la Sociedad Entomológica Argentina. 70 (3-4): 213-229. [ Links ]
CHANI-POSSE, M. 2013. Towards a natural classification of the subtribe philonthina (Coleoptera: Staphylinidae: Staphylinini): a phylogenetic analysis of the Neotropical genera. Systematic Entomology 38: 390-406. [ Links ]
CHANI-POSSE, M.; ASENJO, A. 2013.Systematics and phylogeny of the Andean genera Leptopeltus Bernhauer and Leptopeltoides gen. nov. (Coleoptera: Staphylinidae), with biogeographical notes. Zoologischer Anzeiger 252: 440-456. [ Links ]
CHATZIMANOLIS, S. 2012. Zackfalinus, a new genus of Xanthopygina (Coleoptera: Staphylinidae: Staphylinini) with description of 20 new species. Annals of Carnegie Museum 80 (4): 261-308. [ Links ]
CHATZIMANOLIS, S. 2013. Terataki, a new genus of Staphylinini (Coleoptera: Staphylinidae: Staphylininae) from South America. Zootaxa 3750 (3): 251-264. [ Links ]
COLWELL, R. K.; CHANG, X. M.; JING, C. 2004. Interpolating, Extrapolating, and Comparing incidence-based species accumulation curves. Ecology 85 (10): 2717-2727. [ Links ]
COLWELL, R. K. 2013. EstimateS. Versión 9.1.0. Disponible en: http://viceroy.eeb.uconn.edu/EstimateS. [Fecha de revisión: 25 septiembre 2013]. [ Links ]
ESCOBAR, F.; LOBO, J. M.; HALFFTER, G. 2005. Altitudinal variation of dung beetle (Scarabaeidae: Scarabaeinae) assemblages in the Colombian Andes. Global Ecology and Biogeography 14 (4): 327-337. [ Links ]
FAGUA, G. 1999. Variación de las mariposas y hormigas de un gradiente altitudinal de la cordillera Oriental (Colombia). pp. 317-335. En: Amat, G.; Andrade-C, G.; Fernández, F. (Eds.). Insectos de Colombia. Volumen II. Academia de Ciencias Exactas, Física y Naturales. Colección Jorge Álvarez Lleras. No. 13. Editora Guadalupe Ltda. Bogotá. Colombia. 492 p. [ Links ]
FERNANDEZ-ALONSO, J. L. 2009. Flora de Santa María (Boyacá), guía de campo de los géneros de angiospermas. Serie de guías de campo. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá. 279 p. [ Links ]
FRANK, J. H.; BARRERA, R. 2010. Natural history of Belonuchus Nordmann spp. and allies (Coleoptera: Staphylinidae) in Heliconia L. (Zingiberales: Heliconiaceae) flower bracts. Insecta Mundi 0110: 1-12. [ Links ]
FRANK, J. H.; KANAMITSU, K. 1987. Paederus, sensu lato (Coleoptera: Staphylinidae): Natural history and medical importance. Journal of Medical Entomology 24 (2): 155-191. [ Links ]
GARCÍA, R.; ARMBRECHT, I.; ULLOA-CHACÓN, P. 2001. Staphylinidae (Coleoptera): composición y mirmecofilia en bosques secos relictuales de Colombia. Folia Entomológica Mexicana 40 (1): 1-20. [ Links ]
GARCÍA, R.; CHACÓN DE ULLOA, P. 2005. Estafilínidos (Coleoptera: Staphylinidae) en fragmentos de bosque seco del valle geográfico del río Cauca. Revista Colombiana de Entomología 31 (1): 43-50. [ Links ]
GOTELLI, N. J.; COLWELL, R. K. 2001. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4: 379-391. [ Links ]
GREBENNIKOV, V. V.; NEWTON, A. F. 2009. Good-bye Scydmaenidae, or why the ant-like stone beetles should become megadiverse Staphylinidae sensu latíssimo (Coleoptera). European Journal of Entomology 106: 275-301. [ Links ]
GRIMALDI, D.; ENGEL, M. S. 2005. Evolution of the Insects. Cambridge University Press, New York. 755 p. [ Links ]
GUERRERO, R. J.; SARMIENTO, C. E. 2010. Distribución altitudinal de hormigas (Hymenoptera, Formicidae) en la vertiente noroccidental de la Sierra Nevada de Santa Marta (Colombia). Acta Zoológica Méxicana 26 (2): 279-302. [ Links ]
GUTIÉRREZ-CHACÓN, C.; ULLOA-CHACÓN, P. 2006. Composición de estafilínidos (Coleoptera: Staphylinidae) asociados a hojarasca en tres localidades de la cordillera oriental de Colombia. Folia Entomológica Mexicana 45 (2): 69-81. [ Links ]
HAMMER, Ø.; HARPER, D. A. T.; RYAN, P. D. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4 (1): 1-9. Disponible en http://folk.uio.no/ohammer/past/. [Fecha de revisión: 15 septiembre 2014]. [ Links ]
HODKINSON, I. D. 2005. Terrestrial insects along elevation gradients: species an community responses to altitude. Biological Reviews 80 (3): 489-513. [ Links ]
JIMÉNEZ-SÁNCHEZ, E.; JUÁREZ-GAYTÁN, O. M.; PADILLA-RAMÍREZ, J. R. 2011. Estafilínidos (Coleoptera: Staphylinidae) necrófilos de Malinalco, Estado de México. Universidad de Guadalajara, Dugesiana 18 (1): 73-84. [ Links ]
JIMÉNEZ-VALVERDE, A.; HORTAL, J. 2003. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología 8: 1576-9518. [ Links ]
KLIMASZEWSKI, J. 1984. A revision of the genus Aleochara Gravenhorst of the American north of Mexico (Coleoptera: Staphylinidae, Aleocharinae). Memoirs of the Entomological Society of Canada 129: 1-211. [ Links ]
LÓPEZ-GARCÍA, M. M.; MÉNDEZ-ROJAS, D. M. 2014. A new species of the genus Linoderus Sharp, 1885 (Coleoptera, Staphylinidae, Philonthina) from the Colombian Andes. Zootaxa 3795 (1): 085-090. [ Links ]
LÓPEZ-GARCÍA, M. M.; MÉNDEZ-ROJAS, D. M.; NAVARRETE-HEREDIA, J. L. 2011. First record and a new species of Megarthrus (Staphylinidae: Proteininae) for the Colombian Central Andes. Zootaxa 2916: 62-64. [ Links ]
MAGURRAN, A. E. 1989. Diversidad ecológica y su medición. Ediciones Vedra, España. 200 p. [ Links ]
MAGURRAN, A. E. 2004. Measuring biological diversity. The commonness, and rarity, of species. Blackwell Science, Oxford. 256 p. [ Links ]
MAUS, C.; MITTMANN, B.; PESCHKE, K. 1998. Host records of parasitoid Aleochara Gravenhorst (Coleoptera, Staphylinidae) attacking puparia of cyclorrhapheous Diptera. Deutsche Entomologische Zeitschrift 45: 231-254. [ Links ]
MÉNDEZ-ROJAS, D. M.; LÓPEZ-GARCÍA, M. M.; GARCÍA-CÁRDENAS, R. 2009. Diversidad de escarabajos (Coleoptera, Staphylinidae) en dos localidades del departamento del Quindío. Boletín Científico Centro de Museos, Museo de Historia Natural 13 (2): 148-156. [ Links ]
MÉNDEZ-ROJAS, D. M.; LÓPEZ-GARCÍA, M. M.; GARCÍA-CÁRDENAS, R. 2012. Diversidad de escarabajos (Coleoptera, Staphylinidae) en los bosques altoandinos restaurados de los Andes centrales de Colombia. Revista Colombiana de Entomología 38 (1): 141-147. [ Links ]
NAVARRETE-HEREDIA, J. L. 2001. Beetles associated with Atta and Acromyrmex ants (Hymenoptera: Formicidae: Attini). Transactions of the American Entomological Society 127 (3): 381-429. [ Links ]
NAVARRETE-HEREDIA, J. L.; NEWTON, A. F.; THAYER, M. K.; ASHE, J. S.; CHANDLER, D. S. 2002. Guía ilustrada para los géneros de Staphylinidae (Coleoptera) de México. Universidad de Guadalajara, CANABIO, México. 401 p. [ Links ]
NEWTON, A. F. 1990. Insecta: Coleoptera: Staphylinidae adults and larvae. pp. 1137-1174. En: DINDAL, D. L. (Ed.). Soil biology guide, New York. 1376 p. [ Links ]
NEWTON, A. F.; GUTIERREZ-CHACÓN, C.; CHANDLER, D. S. 2005. Checklist of the Staphylinidae (Coleoptera) of Colombia. Biota Colombiana 6 (1): 1-75. [ Links ]
NEWTON, A. F.; THAYER, M. K.; ASHE, J. S.; CHANDLER, D. S. 2001. Staphylinidae Latreille, 1802. pp 272-418. En: ARNETT, R. H.; THOMAS, M. C. (Eds.). American beetles, Boca Raton. 443 p. [ Links ]
OLSON, D. M. 1994. The distribution of leaf litter invertebrates alonga neotropical altitudinal gradient. Journal of Tropical Ecology 10: 129-150. [ Links ]
SANABRIA, M. C.; AMRBRECHT, I.; GUTIÉRREZ, C. 2008. Diversidad de estafilínidos (Coleoptera: Staphylinidae) en cinco sistemas productivos de los Andes Colombianos. Revista Colombiana de Entomología 34 (2): 217-223. [ Links ]
SLIPINSKI, S. A.; LESCHEN, R. A.; LAWRENCE, J. F. 2011. Order Coleoptera Linnaeus, 1758. pp 203-208. En: Zhang, Z.-Q. (Ed.). Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 3148: 203-208. [ Links ]
TORRES-RODRÍGUEZ, D.; AMAT-GARCÍA, G. D.; NAVARRETE-HEREDIA, J. L. 2012. Sinopsis de los géneros de Xanthopygina (Coleoptera: Staphylinidae: Staphylinini) en Colombia. Universidad de Guadajara, Dugesiana 18 (2): 217-241. [ Links ]
VÁSQUEZ-VÉLEZ, L. M.; BERMÚDEZ, C.; CHACÓN, P.; LOZANO-ZAMBRANO, F. H. 2010. Analysis of the richness of Staphylinidae (Coleoptera) on different scales of a sub-Andean rural landscape in Colombia. Biodiversity Conservation 19: 1917-1931. [ Links ]
ZHANG, Z.-Q. 2013. Phylum Arthropoda. En: Zhang, Z.-Q. (Ed.). An outline of higher-level classification and survey of taxonomic richness (Addenda 2013). Zootaxa 3703: 1-81. [ Links ]
Recibido: 10-feb-2015
Aceptado: 19-ene-2016

