










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Entomología
versión impresa ISSN 0120-0488
Rev. Colomb. Entomol. vol.42 no.2 Bogotá 2016
Hymenoptera parasitoids in protected area of Atlantic Forest biomes and organic rice field: compared assemblages
Hymenoptera parasitoides en áreas protegidas del bioma bosque Atlántico y de arroz orgánico: ensamblajes comparados
GISELE DE SOUZA DA SILVA1,2, SIMONE MUNDSTOCK JAHNKE2,3 and MARÍA LETICIA GONZÁLEZ FERREIRA4
1 M. Sc. giss_rs@yahoo.com.br.
2 UFRGS Programa de Pós-graduação em Fitotecnia, Departamento de Fitossanidade, Av. Bento Gonçalves, 7712 - CEP 91540-000 - Porto Alegre - RS - Brasil.
3 D. Sc. Professor adjunto, smjahnke@yahoo.com, corresponding author.
4 M. Sc. marileti_1@hotmail.com.
ABSTRACT
One way to improve the sustainability of agricultural systems is to generate similar characteristics to those in natural ecosystems by maintaining energy flows and habitat diversity, thereby ensuring the presence of natural enemies and other beneficial organisms that can regulate pest populations and maintain crop productivity with fewer environmental impacts. The objectives of this study were to identify and compare the diversity of parasitoid assemblages in irrigated rice crops under organic management in a nearby protected area; to compare the efficiency of two kinds of parasitoid traps; and to compare temporal variation in parasitoid species at the two sites. The study was developed in the Banhado dos Pachecos Wildlife Refuge (BPWR) and in Viamão, RS organic rice fields (OR). Specimens were collected monthly from May 2011 to April 2012. Two Malaise and four Moericke traps were used in each area. In the BPWR area, Platygastridae, Ichneumonidae and Braconidae showed the highest abundance (30 %, 21 % and 11 %, respectively), and in the OR area, the dominant taxa were Platygastridae (26 %), Braconidae (18 %) and Encyrtidae (15 %). Malaise traps captured the largest number of parasitoids (58 %). The richness estimators Chao 1, Jack 1 and Bootstrap, pointed to a richness of 229 to 122 species in the OR area and of 454 to 260 in the BPWR area. Parasitoid diversity was higher in the BPWR than in the OR. Parasitoid abundance was highest in the rice crop during months in which crops were growing at the site.
Key words: Agricultural ecosystems. Conservation biological control. Natural enemies.
RESUMEN
Una manera de mejorar la sostenibilidad de los sistemas agrícolas es generar características similares a las de los naturales manteniendo los flujos de energía y la diversidad de hábitat, lo que garantiza la presencia de enemigos naturales y otros organismos benéficos que pueden regular las poblaciones de plagas y mantener la productividad con menores impactos ambientales. Este estudio tuvo como objetivos: identificar y comparar la diversidad de las comunidades de parasitoides en arroz irrigado orgánico y en un área de reserva cercana al cultivo, comparar la eficacia de dos tipos de trampas y la variación temporal de las especies de parasitoides en los dos sitios. El estudio se llevó acabo en el Refugio de Vida Silvestre Bañado Pacheco (RVSBP) y en los campos de arroz orgánico (AO), Viamão, RS. Se realizaron colectas mensuales de mayo de 2011 a abril de 2012 con dos trampas de Malaise y cuatro Moericke. En el área RVSBP, Platygastridae, Ichneumonidae y Braconidae se presentaron con la mayor abundancia (30 %, 21 % y 11 %, respectivamente) mientras que en AO fueron Platygastridae (26 %), Braconidae (18 %) y Encyrtidae (15 %). La trampa Malaise capturó el mayor número de parasitoides (58 %). Los estimadores de riqueza Chao 1, Jack 1 y "bootstrap" apuntaron a una riqueza de 229 a 122 especies en AO y de 454 a 260 en el área RVSBP. La diversidad de parasitoides fue mayor en el RVSBP en comparación a la del AO. La abundancia de parasitoides fue mayor en los meses durante el ciclo del cultivo del arroz.
Palabras clave: Ecosistemas agrícolas. Control biológico de conservación. Enemigos naturales.
Introduction
An ecosystem is a functional system of complementary relationships between living organisms and their surroundings, defined by boundaries within which they are able to maintain a dynamic and stable equilibrium (Gliessman 2001). Because the diversity of ecosystems makes them resistant to disturbance and outside interference, more diverse ecosystems have a greater capacity for recovering from disturbance and restoring equilibrium to their processes of cycling materials and energy flows (Cuddington 2001). The low diversity of conventional agricultural systems (e.g., monocultures treated with large amounts of synthetic fertilizers and pesticides) makes them biologically unstable and vulnerable to economically damaging pests and disease agents (Gliessman 2001). Alternative agricultural practices in harmony with existing ecological processes in agricultural ecosystems can thus help increase the sustainability of food production (Gliessman 2001; Altieri et al. 2003; Altieri 2012). One way to improve the sustainability of agricultural systems is to generate characteristics similar to those in natural ecosystems by maintaining energy flows and habitat diversity, thereby ensuring the presence of natural enemies and other beneficial organisms that can regulate pest populations and maintain crop productivity with fewer environmental impacts (Gliessman 2001). In natural systems, methods to estimate diversity allow for more efficient environmental conservation and monitoring. Even as, in both natural and anthropogenic systems, diversity is considered a synonym of environmental quality, since it responds to adverse impacts such as those caused by pollution or community imbalances. Some estimation methods are based on community structure, while others rely on dominance or evenness (Moreno 2001).
No other feature of agricultural systems offers as many fundamental ecosystem services for protecting plants against insect pests as plant diversity (Altieri and Letourneau 1982; Altieri et al. 2003). For that reason, conserving native forests close to agricultural ecosystems helps maintain and increase biodiversity in the latter, thereby boosting ecological processes there. Brazil';s government mandated the conservation of native forests within rural properties, by the Brazilian Forest Code (12.651/12, with alterations in Law 12.727/12) (Presidência da República Federativa do Brasil 2012). These undisturbed patches of native forest within a property (known as Legal Reserves) ensure that economic activities carried out there use natural resources sustainably, help preserve and restore ecological processes, and help conserve biodiversity, wildlife, and the native flora.
Rice (Oryza sativa L.), an annual grass native to Asia, was domesticated roughly 10,000 years ago (Khush 1997; Heinrichs 1998; Bambaradeniya and Amarasinghe 2003). Some insect and other phytophagous species can become common enough in irrigated rice crops to be economically important pests, causing losses in productivity on the order of 10-35 % (Martins et al. 2009). One method for controlling these pests without serious environmental impacts is biological control. Parasitoids are one of the most important biological control agents for pests in agricultural systems, due both to their natural occurrence and to their use in biological control programs (Bale et al. 2008). Given that most organisms regarded as pests of rice are known to be attacked by parasitoids, controlling them via conservation of parasitoid hymenopterans is an important tool. The diversity of such parasitoids in different agricultural systems depends on environmental and biological factors, as well as management practices (Chay-Hernandez et al. 2006). Rice plantations are surrounded by aquatic and terrestrial habitats that form a mosaic of dynamic environments harbouring large stores of biological diversity, which are maintained both by rapid colonization and by the organisms'; rapid reproductive and growth rates (Fritz et al. 2008). The fauna associated with these systems includes vertebrates and invertebrates that inhabit the vegetation, water, and soils of rice plantations (Hook 1994). This diversity provides a more complex environment around the cultivated areas, where ecological interactions can be monitored by using parasitoid richness as a biological indicator (Lockwood et al. 1996).
The objectives of this study were: 1) to identify and compare the diversity of parasitoid assemblages in an irrigated rice crop under organic management and in a nearby protected area (both located in an Environmental Protection Area [Área de Proteção Ambiental]); 2) to compare the efficiency of two kinds of parasitoid traps; and 3) to compare the temporal variation in parasitoid species at the two sites.
Materials and methods
Study sites. The study was carried out in the 133,000 ha Banhado Grande Environmental Protection Area. Banhado Grande APA includes portions of the Pampa and Atlantic Forest biomes and accounts for 2/3 of the watershed of the Gravataí River. The original vegetation is primarily wetlands and "restinga" forest, but today the area also harbours urban centres and agricultural lands, where rice crops predominate (SEMA 2011).
Collections were made at two study sites: one a well protected area of native vegetation and the other an organic rice plantation in the Águas Claras district (30°08';8.60"S, 50°53';52.60"W) of the town of Viamão, in the state of Rio Grande do Sul, Brazil.
Wildlife refuge. The first study site, a well preserved area of native vegetation known as the Banhado dos Pachecos Wildlife Refuge (BPWR), comprises 2,560 ha and is designated as a Legal Reserve of the "Filhos de Sepé" settlement (see below) under the Brazilian Forest Code (12.651/12, with alterations in Law 12.727/12).
The site has vegetation consisting of various different associations of plant species. Because plant collections are not permitted at the site, plants that occur frequently in the BPWR were identified based on photographs and the taxonomic literature (Lorenzi 1982; 1994), or by consulting descriptions published in other studies (Accordi and Hartz 2006). The families most commonly observed at the site were: Annonaceae, Apiaceae, Araliaceae, Asteraceae, Arecaceae, Blechnaceae, Cyperaceae, Ephedraceae, Eriocaulaceae, Lauraceae, Malvaceae, Melastomataceae, Fabaceae, Moraceae, Myrsinaceae, Myrtaceae, Onagraceae, Orchidaceae, Phytolaccaceae, Poaceae, Polygonaceae, Pontederiaceae, Rubiaceae, Typhaceae, and Verbenaceae.
Rice field. The second study site is an approximately 20 ha rice plantation that forms part of the "Filhos de Sepé" Landless Rural Workers Movement Settlement, which surrounds the Banhado dos Pachecos. It is the largest such settlement in the state (9,406 ha) and home to 376 families (SEMA 2011). Because they are located within an Environmental Protection Area, since 2007 these rice plantations have been managed with organic practices.
The rice cultivar IRGA417 was planted with pregerminated seeds under a layer of water or in mud in October 2011 and harvested in late February 2012.
The rice field has a variety of wild vegetation that grows on the levees and in the paddies outside of planting season. To identify the wild plant species on the levees, reproductively mature material of the most frequently observed species was collected and herbarium specimens prepared. Plants were identified using the taxonomic literature (Lorenzi 1982; 1994) and with the help of Dr. Ilsi Boldrini of the Postgraduate Program in Botany at the Federal University of Rio Grande do Sul (UFRGS). The most commonly observed families at this site were Asteraceae, Boraginaceae, Commelinaceae, Convolvulaceae, Cyperaceae, Onagraceae, Poaceae, Polygonaceae, and Solanaceae.
Sampling. Traps were installed at two sampling points in the BPWR study site. Both points featured "restinga" vegetation, a characteristic ecosystem of the Atlantic Forest biome (Presidência da República Federativa do Brasil 1993). Point 1 (R1) had vegetation resembling that of High Restinga Forest while Point 2 (R2) had vegetation characteristic of Low Restinga Forest (Secretaria do Meio Ambiente do Estado de São Paulo 2013). Two sampling points were also established on the levees of the rice plantation: Point 1 (A1) and Point 2 (A2).
Sampling was carried out every month from May 2011 to April 2012. To collect hymenopteran parasitoids, two Malaise traps (Towes 1972a) and four Moericke traps (Granger 1970) were installed by two transects distant about 400 m from each other and 20 m between them at both the OR site and the BPWR site. The traps were left for 24 hours.
The collected insects were stored in 70 % alcohol, labelled with sampling point and trap type, and transported to the Biological Control laboratory of the Plant Health Department of the Federal University of Rio Grande do Sul (UFRGS). The insects were separated into morphospecies with a Nikon SMZ445 stereomicroscope. Families of Hymenoptera were identified followed the classification system adopted by Goulet and Huber (1993).
Meteorological data of the city of Viamão for mean daily temperature and monthly rainfall were obtained from Brazil';s National Meteorological Institute (INMET 2012).
Data analysis. The average number of hymenopteran parasitoids individuals collected on each sampling occasion was compared between trap types, sampling sites, and seasons using analysis of variance (ANOVA, Kruskal-Wallis test) (Hammer et al. 2001).
Rarefaction curves were constructed to compare richness between the rice field and the native forest. Estimated richness was calculated for each study site via the Chao 1, Jackknife 1, and Bootstrap estimates, using EstimateS software, version 8.2.0 (Colwell 2009).
Qualitative differences were demonstrated separating exclusive and shared morphospecies between areas using Venn diagram.
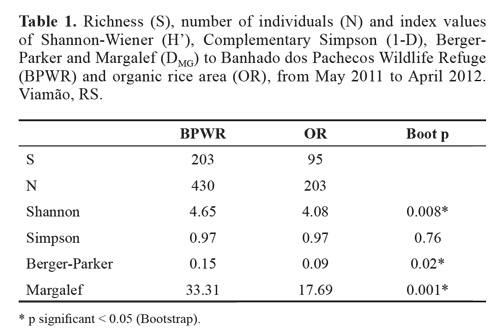
Biological diversity was analysed with the Shannon- Wiener (H';), Simpson (1-D), Margalef (DMg), and Berger- Parker indices, using Past software, version 2.10 (2011) (Hammer et al. 2001).
Results
A total of 430 individual parasitoid wasps were collected at the Banhado dos Pachecos Wildlife Refuge (BPWR), belonging to 203 morphospecies and 20 families. A total of 203 individuals were collected at the organic rice plantation (OR), belonging to 95 morphospecies and 19 families. Platygastridae, Ichenumonidae, and Braconidae were the most abundant families at the BPWR site, accounting for 30 %, 21.40 %, and 11.40 % of relative frequency of individuals, respectively (Fig. 1). At the OR site, Platygastridae (26.11 %), Braconidae (18.23 %), and Encyrtidae (15.27 %) (Fig. 1).were the most abundant families.
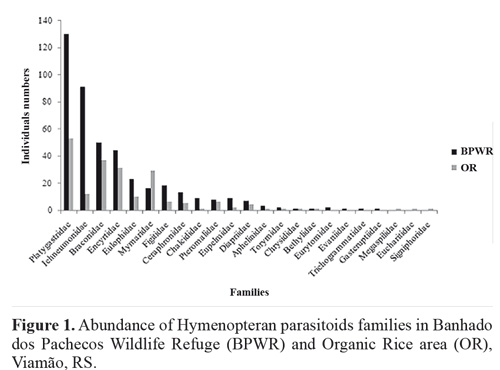
Each Malaise trap captured a mean of 13.8 ± 3.46 individuals, significantly more than the Moericke traps (5.5 ± 1.15) (H = 4.84; d.f.= 1; P < 0.05). Ichneumonidae (21.80 %), Platygastridae (18.53 %), and Braconidae (16.62 %) were the most abundant families captured in the Malaise traps, while Platygastridae (42.85 %), Encyrtidae (18.42 %), and Braconidae (9.40 %) were the most abundant in Moericke traps. Malaise traps yielded all the families reported in this study, while Moericke traps were more selective, and did not capture individuals of Aphelinidae, Bethylidae, Eucharitidae, Eurytomidae, Evaniidae, Gasteruptiidae, Megaspilidae, Signiphoridae, Trichogrammatidae, and Torymidae.
Total richness was 203 morphospecies for the BPWR site and 95 for the OR site.
Thirty-seven morphospecies were shared between sites (Fig. 2). The highest number (26) was during the crop cycles (October/2011 to February/2012).

Non-parametric estimators of species richness are typically based on the richness of rare species, and thus on four variables: singletons and doubletons (i.e., species represented by one or two individuals at a site), and unicates and duplicates (i.e., species collected during just one or two sampling periods) (Colwell 2009). The observed number of species (Sobs) at each site indicates that richness was not fully sampled, as illustrated by the ascending curves in Figs 4 and 5. At the BPWR site there were 137 singletons, 36 doubletons, 153 unicates, and 32 duplicates, while at the OR site there were 64 singletons, 14 doubletons, 72 unicates, and 14 duplicates. The richness estimators suggested that 40 % to 78 % of total richness was collected at the two sites (Figs. 3 and 4).
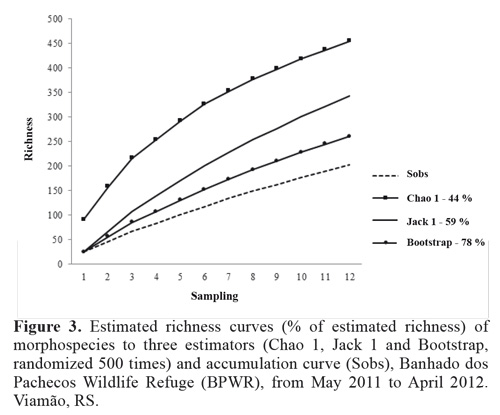
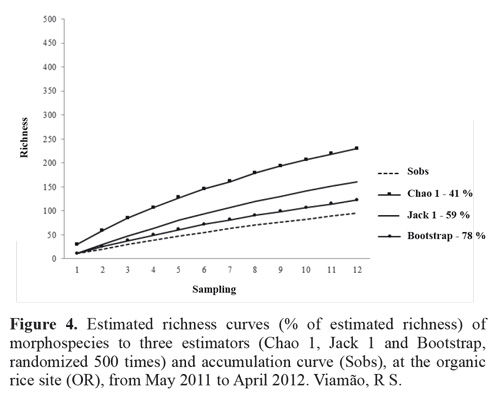
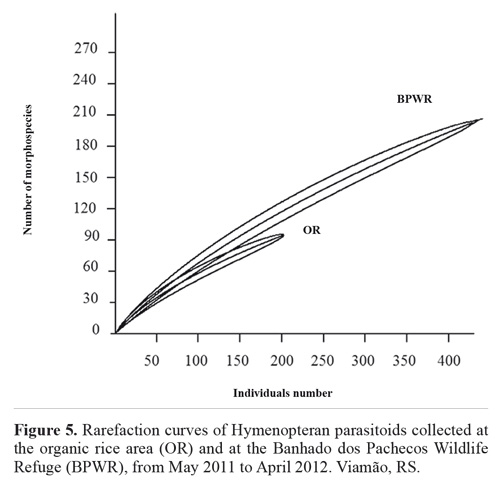
The rarefaction curve for the abundance data shows species richness differing between the two sites (Fig. 5), and the Margalef, Shannon, Simpson, and Berger-Parker indices (Table 1) also indicated higher diversity at the BPWR site. As expected, abundances at both study sites varied over time throughout the study (Fig. 6). June, July, and August had the fewest captures and were very rainy months, but no correlation between rainfall and number of captured insects was observed. Abundances peaked in November for both study sites.
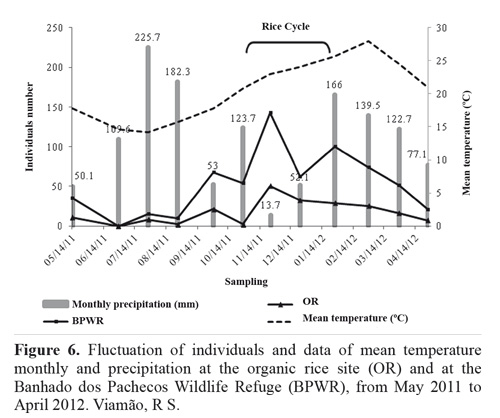
The Pearson correlation coefficient indicated a positive correlation between abundance and temperature for the OR site (P < 0.05, R2 = 0.393) and the BPWR site (P < 0.05; R2 = 0.383).
Discussion
The most abundant family and morphospecies shared in this study, Platygastridae, has been recorded in several other studies of well preserved environments in Brazil';s Atlantic Forest, even when different sampling methods were used (e.g., sweep nets), or when all three methods (Malaise, Moericke, and sweep nets) were used (Azevedo and Santos 2000; Azevedo et al. 2002; Azevedo et al. 2003). Likewise, some families found to be rare in this study (e.g. Aphelinidae, Chrysididae, Evaniidae, Eurytomidae, Torymidae, and Trichogrammatidae), have also been reported to have abundances below 1 % in other studies (Azevedo and Santos 2000; Azevedo et al. 2002).
The low abundance of Ichneumonidae in the rice plantation may be explained by the fact that species of the family are more numerous in temperate and wet tropical regions (Townes 1972b). Humidity is higher in forested environments, and is associated with the abundance of these insects (Townes 1972b). Although the rice plantation is irrigated, it is an open environment more vulnerable to variation in environmental conditions, which may have contributed to its lower species richness in this family. By contrast, the BPWR site has more shade due to its denser vegetation, which offers a more humid microclimate with less extreme variations in temperature and humidity inside the forest (Tanque et al. 2010). It would thus appear that oscillations in environmental conditions affect the ichneumonid community to a lesser degree at the BPWR site. This result is similar to that of another survey carried out in an organic rice plantation in Rio Grande do Sul state, in the towns of Capivari do Sul, Eldorado do Sul, and Cachoeira do Sul. That study found Eulophidae (131 individuals), Braconidae (69 individuals), and Platygastridae (39 individuals) to be the most common families (Fritz et al. 2011). Species in these families were also found in our study and have been noted in other studies to attack pests in rice plantations.
A parasitism rate of 32 % was found in Tibraca limbativentris Stål, 1860, parasitized by Telenomus podisi (Ashmead) and Trissolcus urichi (Crawford) (Platygastridae) and Oencyrtus submetallicus (Howard) (Encyrtidae) (Maciel et al. 2007). This natural parasitism is important since Pentatomidae have been reported throughout Brazil and can reduce crop productivity by up to 90 % (Ferreira et al. 1997). In rice crops in southern Brazil, T. limbativentris was found to be parasitized by T. uruchi (7.7 %) and T. podisi (75 %) (Riffel et al. 2010; Idalgo et al. 2013), and eggs of O. poecilus were found to be parasitized by T. podisi (Krinski et al. 2011).
The composition of families and species of parasitoid communities can also vary between sites and between crops due to their pest hosts, which may vary dramatically from one crop to another. The most abundant parasitoid found by Bambaradeniya and Edirisinghe (2008) in a study of rice in Sri Lanka, for example, was the family Mymaridae (39 %), which attack the insects Nilaparvata lugens (Stål, 1854) and Sogatella furcifera (Horváth, 1899) (Hemiptera: Delphacidae), important pests in that region, different from our work in which this family came fourth in abundance.
In rice fields, Braconidae is known by parasitize Spodoptera frugiperda (Smith, 1797), Elasmopalpus lignosellus (Zeller) (Lepidoptera: Pyralidae) and Cotesia flavipes Cameron [= Apanteles flavipes (Cameron)] (Braconidae) has been reported to D. saccharalis (Embrapa Arroz e Feijão 2004).
The highest number of morphospecies shared, 39 % of OR site, indicates the importance of Legal Reserve Area like natural enemies repository. The diversity vegetation increase in agricultural landscape can be a solution for beneficial insects increase (Thomas et al. 1991).
Trap efficiency can also depend on the surrounding vegetation, species'; habitats, and specific attributes such as flight capacity and host type (Southwood 1978). In our study, Malaise traps were installed in areas with sparse vegetation cover that may serve as corridors for dispersing insects (i.e., levees and resting forest), and this could have resulted in the greater number of individuals captured in the Malaise traps. In heterogeneous environments such as tropical forest, using multiple sampling methods is recommended for assessing the diversity of hymenopteran parasitoids (Noyes 1989). Malaise traps are efficient at collecting winged parasitoids because they intercept any insect in flight, while Moericke traps depend on the degree to which each group is attracted to colour (Mazón and Bordera 2008).
Marchiori et al. (2003) obtained similar results to ours. They found that the Ichneumonidae family accounted for the largest number of individuals in Malaise traps (35.1 %), followed by Braconidae (19.1 %), while Diapriidae (26.7 %) and Encyrtidae (11.5 %) were the most abundant families in Moericke traps in south eastern Brazil. Thus, inventories designed to compare insect diversity and abundance should use sampling methods based on the sampling site and the group under study, both of which influence which types of insects will be captured.
The estimated richness of the study sites appears similar to those published in other studies of Neotropical environments (Querino et al. 2011). The variation in diversity values obtained with the estimators reflects the different parameters used in each. The first-order Jackknife estimator uses the unicates (Heltshe and Forrestor 1983). By contrast, the Chao 1 estimator is a function of the ratio between singletons and doubletons. At both study sites there were large numbers of singletons, which played an important role in defining the value of the Chao 1 estimator (Figs. 4 and 5).
Differences between the diversity indices of BPWR and OR were expected. An increase in the index value may reflect higher richness, greater evenness, or both (Magurran 2011). In a survey of hymenopteran parasitoid diversity in soybean crops, for example, Lara et al. (2009) found H'; values of 0.48, much lower than that found in our study, even though those authors recorded 22 families. The authors argued that these low values reflect the low evenness of their sample. Since Margalef';s index reflects richness based on the number of individuals (Moreno 2001) and both number of species and abundance were more than twice as high at the BPWR site than at the OR site, this index showed a strong difference between the sites. The only index that did not find a difference between the sites was Simpson';s, which is strongly affected by the most dominant species in the sample and less sensitive to species richness (Magurran 2011). As both sites had few very abundant species and many rare species, this index found no difference between the two.
In our study, the observed diversity in the smaller community did not reach the 95 % confidence interval of the larger community, by rarefaction technique. The comparison is made at the point at which the abundance level of the larger community is identical to that of the smaller community (Gotelli and Entsminger 2001).
The difference in the diversity of hymenopteran parasitoids between the OR and BPWR sites may reflect their different vegetation structure. Although the OR site is managed organically, the agricultural environment is still less complex (Gliessman 2001) and the differences in plant families and genera between the two sites are apparent and well documented (Accordi and Hartz 2006). Diversity in vegetation structure and species is one of the factors that create niche diversity (Wilson 1994), and consequently resources for adult parasitoids to find shelter and food. Several studies have shown that the abundance and richness of entomophagous insects within an agricultural environment are closely related to the nature of the surrounding vegetation (Altieri et al. 2003). Adult hymenopteran parasitoids are also free-living and feed on honey and pollen (Jervis et al. 1993), which are provided by several wild plants, especially during flowering season.
Species richness, abundance, and composition can vary both with niche-related spatial heterogeneity and with temporal variation in weather and seasonal changes within a given area. Studies with hymenopteran parasitoids have shown highest abundance in austral spring and summer, lower abundance in autumn, and lowest abundance in winter in "restinga" environments in the town of Rio Grande, in Rio Grande do Sul state (Oliveira et al. 2009). The same authors found Ichneumonidae and Braconidae to be the most common parasitoid families, and also reported a 70 % correlation between temperature and abundance. No correlation was found with rainfall, corroborating our data from the BPWR site. While these abiotic factors are important, other parameters unrelated to weather, such as food availability, can also affect seasonal diversity patterns (Wolda 1988).
The peak abundance in November at both sites was strongly influenced by the abundance of Platygastridae and Braconidae. The greater abundance of Braconidae can be explained by the family';s preference for open vegetation (like the rice plantation and "restinga") and for mean temperatures between 20 and 24 °C (Cirelli and Penteado-Dias 2003).
Platygastridae reached its highest abundance in the austral summer and spring, at both the BPWR and OR sites. Peak abundance in this family could potentially be related to host availability. The months with the greatest abundance of platygastrids were those in which the rice crop was growing (November-February). The BPWR site showed high abundance during the same months and also in October, when the rice was starting to be planted. This may be additional evidence that the BPWR can supply the rice crop with parasitoids during the season in which it is most vulnerable to pest attacks.
Parasitoid abundance in the rice plantation is affected not only by temperature but also by the presence or absence of the crop. The months between November and February had the highest abundance, since as rice matures there is more food available for the pests that serve as parasitoid hosts. Once seedlings emerge after November, these form a type of vegetation that is different from that of the winter landscape. During this same time wild plants growing on the levees, mostly Asteraceae and Cyperaceae, flower and provide resources for adult parasitoids (Coombes and Sotherton 1986; Corbett and Planta 1993; Jervis et al. 1993).
Given that there are 61 families of hymenopteran parasitoids in the word, and that several are restricted to specific zoogeographical regions such as the Australian and the Holarctic (Azevedo and Santos 2000), and considering that 36 hymenopteran families with parasitizing species and Chrysidoidea have been recorded in Brazil (De Santis 1980), the sites we studied appear to have a high parasitoid diversity and to play a key role in maintaining the diversity of these natural enemies.
Conclusions
The Malaise traps yielded more hymenopteran parasitoids and a higher diversity of those parasitoids than the Moericke traps. Parasitoid diversity was higher at the Banhado dos Pachecos Wildlife Refuge than at the organic rice crop. Parasitoid abundance was highest in the rice crop during the months in which the crop was growing at the site.
Literature cited
ACCORDI, I. A.; HARTZ, S. M. 2006. Distribuição espacial e sazonal da avifauna em uma área úmida costeira do sul do Brasil. Revista Brasileira de Ornitologia 14 (2): 117-135. [ Links ]
ALTIERI, M. A.; LETOURNEAU, D. L. 1982. Vegetation management and biological control in agroecosystems. Crop Protection 1: 405-430. [ Links ]
ALTIERI, M. A.; SILVA, E. N.; NICHOLLS, C. I. 2003. O papel da biodiversidade no manejo de pragas. Holos, Ribeirão Preto. 223 p. [ Links ]
ALTIERI, M. A. 2012. Agroecologia: bases científicas para uma agricultura sustentável. 3. ed. rev. ampl. Expressão Popular, São Paulo, AS-PTA, Rio de Janeiro. 400 p. [ Links ]
AZEVEDO, C. O.; SANTOS, H. S. 2000. Perfil da fauna de himenópteros parasitoides (Insecta, Hymenoptera) em uma área de Mata Atlântica da Reserva Biológica de Duas Bocas, Cariacica, ES, Brasil. Boletim do Museu de Biologia Mello Leitão 11 (12): 117-126. [ Links ]
AZEVEDO, C. O., KAWADA, R., TAVARES, M. T.; PERIOTO, N. W. 2002. Perfil da fauna de himenópteros parasitóides (Insecta, Hymenoptera) em uma área de Mata Atlântica do Parque Estadual da Fonte Grande, Vitória, ES, Brasil. Revista Brasileira de Entomologia 46 (2): 133-137. [ Links ]
AZEVEDO, C. O.; CORRÃA, M. S.; GOBBI, F. T.; KAWADA, R.; LANES, G. O.; MOREIRA, A. R.; REDIGHIERI, E. S.; SANTOS, L. M. DOS.; WAICHERT, C. 2003. Perfil das famílias de vespas parasitoides (Hymenoptera) em uma área de Mata Atlântica da Estação Biológica de Santa Lúcia, Santa Teresa, ES, Brasil. Boletim do Museu de Biologia Mello Leitão16: 39-46. [ Links ]
BALE, J. S.; LENTEREN, J. C. VAN.; BIGLER, F. 2008.Biological control and sustainable food production. Philosophical Transactions of The Royal Society Biological Science 363: 761- 776. [ Links ]
BAMBARADENIYA, C. N. B.; AMERASINGHE, F. P. 2003. Biodiversity associated with the rice field agro-ecosystem in Asian countries: a brief review. International Water Management Institute, Colombo, Sri Lanka, 63. 24 p. [ Links ]
CIRELLI, K. R. N.; PENTEADO-DIAS, A. M. 2003. Análise da riqueza da fauna de Braconidae (Hymenoptera, Ichneumonoidea) em remanescentes naturais da Área de Proteção Ambiental (APA) de Descalvado, SP. Revista Brasileira de Entomologia 47 (1): 89-98. [ Links ]
CHAY-HERNANDEZ, D. A.; DELFÍN-GONZÁLEZ, H.; PARRATABLA, V. 2006. Ichneumonoidea (Hymenoptera) community diversity in an agricultural environment in the State of Yucatan, Mexico. Environmental Entomology 35 (5): 1286-1297. [ Links ]
COLWELL, R. K. 2009. EstimateS 8: statistical estimation of species richness and shared species from samples. Hartford: University of Connecticut. Available at: http://viceroy.eeb.ucon.edu/estimates. [Review date: 14 August 2011] [ Links ].
COOMBES, D. S.; SOTHERN, N. M. 1986. The dispersal and distribution of polyphagous predatory Coleoptera in cereals. Annals of Applied Biology 108: 461-474. [ Links ]
CORBETT, A.; PLANTA, R. E. 1993. Role of movement in the response of natural enemies to agroecosystem diversification: a theoretical evaluation. Environmental Entomology 22: 519-531. [ Links ]
CUDDINGTON, K. 2001. "The 'Balance of Nature'; metaphor and equilibrium in population ecology". Biology and Philosophy 16: 463-479. [ Links ]
DE SANTIS, L. 1980. Catalogo de los himenópteros brasileños de la serie parasítica incluyendo Bethyloidea: Editora da Universidade Federal do Paraná, Curitiba. 395 p. [ Links ]
EMBRAPA ARROZ E FEIJÃO. 2004. Cultivo do arroz irrigado no Estado do Tocantins. Manejo dos principais insetos fitófagos. Sistemas de Produção, n° 3, versão eletrônica Nov. 2004. Available at: http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Arroz/ArrozIrrigadoTocantins/manejo_insetos_ fitofagos.htm. [Review date: 07 September 2011] [ Links ].
FERREIRA, E.; ZIMMERMANN, F. J. P.; SANTOS, A. B. DOS; NEVES, B. P. DAS. 1997. O percevejo-do-colmo na cultura do arroz. Embrapa-CNPAF, Goiânia. 43 p. [ Links ]
FRITZ, L. L.; HEINRICHS, E. A.; PANDOLFO, M.; SALLES, S. M. DE; OLIVEIRA, J. V. DE; FIUZA, L. M. 2008. naturais e manejo integrado. Oecologia Brasiliensis 12 (4): 720- 732. [ Links ]
FRITZ, L. L.; HEINRICHS, E. A.; MACHADO, V.; ANDREIS, T. F.; PANDOLFO, M.; SALLES, S. M. DE; OLIVEIRA, J. V. DE; FIUZA, L. M. 2011. Diversity and abundance of arthropods in subtropical rice growing areas in the Brazilian south. Biodiversity and Conservation 20: 2211-2224. [ Links ]
GLIESSMAN, S. R. 2001. Agroecologia: processos ecológicos em agricultura sustentável. Editora da UFRGS, Porto Alegre. 653 p. [ Links ]
GOTELLI, N. J.; ENTSMINGER, G. L. 2001. Ecosim null model software ecology. Version 6.0 Acquired Intelligence Inc. & Keysey-Bear. Available at: http://www.uvm.edu/~ngotelli/EcoSim/EcoSim.html. [Review date: 01 August 2011] [ Links ].
GOULET, H.; HUBER, J. T. 1993. Hymenoptera of the world: an identification guide to families. Agriculture Canada Publication, Ottawa. 668 p. [ Links ]
GRANGER, C. A. 1970. Trap design and color as factors in trapping the salt marsh greenhead fly. Journal of Economic Entomology 63: 1670-1672. [ Links ]
HAMMER, Ã.; HARPER, D. A. T.; RYAN, P. D. 2001. PASt: Paleontological statistics software package for education and data analyses. Paleontologia Electronica 4 (1): 9. [ Links ]
HEINRICHS, E. A. 1998. Management of rice insect pests. Radcliffe';s IPM World Textbook, Minnesota. p. 16. Available at: http://ipmworld.umn.edu. [Review date: 12 August 2011] [ Links ]
HELTSCHE, J.; FORRESTOR, N. E. 1983. Estimating species richness using the jackknife procedure. Biometrics 39: 1-11. [ Links ]
HOOK, T. V. 1994. The conservation challenge in agriculture and the role of entomologists. Florida Entomologist 77: 42-73. [ Links ]
IDALGO, T. D. N.; SANT';ANA, J.; REDAELLI, L. R.; PIRES, P. D. S. 2013. Parasitismo de ovos de Tibraca limbativentris Stål (Hemiptera: Pentatomidae) em lavoura de arroz irrigado, Eldorado do Sul, RS. Arquivos do Instituto Biológico 80 (4): 453-456. [ Links ]
INMET (Instituto Nacional de Metereologia) 2012. Estações e dados. Available at: http://www.inmet.gov.br. [Review date: June 2012] [ Links ].
JERVIS, M. A.; KIDD, N. A. C.; FITTON, M. G.; HUDDLESTON, T.; DAWAH, H. A. 1993. Flower-visiting by hymenopteran parasitoids. Journal of Natural History 27: 67-105. [ Links ]
KHUSH, G. S. 1997. Origin, dispersal, cultivation and variation of rice. Plant Molecular Biology 35: 25-34. [ Links ]
KRINSKI, D.; LOIÁCONO, M. S.; MARGARÍA, C. B.; AQUINO, D. 2011. Primeiro registro de parasitismo em ovos de Oebaluspoecilus(Dallas) (Hemiptera: Pentatomidae) por Telenomus podisi Ashmead (Platygastridae: Telenominae) em arroz tipo sequeiro no Estado do Pará, Brasil. In: SIMPÓSIO DE CONTROLE BIOLÓGICO, São Paulo, 2011. São Paulo: SICONBIOL, 2011. Available at: seb.org.br/eventos/ SINCONBIOL2011/.../PT0628.pdf. [Review date: 12 October 2012] [ Links ].
LARA, R. I. R.; PERIOTO, N. W.; RAMIRO, Z. A. 2009. Número mínimo de armadilhas de Möricke em amostragem de himenópteros parasitoides na cultura da soja Glycine max (L.) Merrill. Arquivos do Instituto Biológico 76 (1): 55-59. [ Links ]
LOCKWOOD, J.; SHAW, S. R.; STRUTTMANN, J. 1996. Biodiversity of wasp species (Insecta: Hymenoptera) in burned and unburned habitats of Yellowstone National Park, USA. Journal of Hymenoptera Research 5: 1-15. [ Links ]
LORENZI, H. 1982. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas, tóxicas e medicinais. Nova Odessa, São Paulo. 425 p. [ Links ]
LORENZI, H. 1994. Plantas daninhas do Brasil. Nova Odessa, Plantarum. 440 p. [ Links ]
MACIEL, A. A. S.; LEMOS, R. N. S. DE, SOUZA; J. R. DE, COSTA, V. A.; BARRIGOSSI, J. A. F.; CHAGAS, E. F. DAS. 2007. Parasitismo de ovos de Tibraca limbativentris Stål (Hemiptera: Pentatomidae) na cultura do arroz no Maranhão. Neotropical Entomology 36 (4): 616-618. [ Links ]
MAGURRAN, A. E. 1988. Ecological diversity and its measurement. Croom Helm, London. 179 p. [ Links ]
MAGURRAN, A. E. 2011. Medindo a diversidade biológica.Editora UFPR, Curitiba. 261 p. [ Links ]
MARCHIORI, C. H.; SILVA, M. H. O.; BRITO, B. M. C.; FILHO, O. M. S.; PEREIRA, L. A. 2003. Levantamento de famílias de parasitóides coletadas em Araporã-MG usando armadilhas de bacias amarelas e malaise. Semina: Ciências Agrárias 24 (2): 317-320. [ Links ]
MARTINS, J. F. DA S.; BARRIGOSSI, J. A. F.; OLIVEIRA, J. V. DE; CUNHA, U. S. DA. 2009. Situação do manejo integrado de insetos-praga na cultura do arroz no Brasil. Embrapa Clima Temperado, Pelotas. 40 p. [ Links ]
MAZÓN, M.; BORDERA, S. 2008. Effectiveness of two sampling methods used for collecting Ichneumonidae (Hymenoptera) in the Cabañeros National Park (Spain). European Journal of Entomology 105: 879-888. [ Links ]
MORENO, C. E. 2001. Métodos para medir la biodiversidad. M & T-Manuales y Tesis SEA.Unesco & SEA, Zaragoza. 84 p. [ Links ]
NOYES, H. S. 1989. The study of five methods of sampling Hymenoptera (Insecta) in a tropical rainforest, with special reference to the parasitica. Journal of Natural History 23 (3): 285-298. [ Links ]
OLIVEIRA, E. A.; CALHEIROS, F. N.; CARRASCO, D. S.; ZARDO, C. M. L. 2009. Famílias de Hymenoptera (Insecta) como ferramenta avaliadora da conservação de restingas no extremo sul do Brasil. EntomoBrasilis 2 (3): 64-69. [ Links ]
PRESIDÊNCIA DA REPÚBLICA FEDERATIVA DO BRASIL. 1993. Decreto 750, de 10 de fevereiro de 1993. Dispõe sobre o corte, a exploração e a supressão de vegetação primária ou nos estágios avançado e médio de regeneração da Mata Atlântica, e dá outras providências. Presidência da República, Casa Civil, Subchefia para Assuntos Jurídicos, Brasília, 10 fev. 1993, 172° da Independência e 105° da República. Available at: http://www.planalto.gov.br/ccivil_03/decreto/1990-1994/D750.htm. [Review date: 05 February 2013] [ Links ].
PRESIDÊNCIA DA REPÚBLICA FEDERATIVA DO BRASIL. 2012. Código Florestal Brasileiro Lei 12651 de 2012. Dispõe sobre a proteção da vegetação nativa, altera as Leis nos 6.938, de 31 de agosto de 1981, 9.393, de 19 de dezembro de 1996, e 11.428, de 22 de dezembro de 2006, revoga as Leis nos 4.771, de 15 de setembro de 1965, e 7.754, de 14 de abril de 1989, e a Medida Provisória no 2.166-67, de 24 de agosto de 2001, e dá outras providências. Presidência da República, Casa Civil, Subchefia para Assuntos Jurídicos, Brasília, 25 mai. 2012, 191° da Independência e 124° da República. Available at: http://www.planalto.gov.br/ccivil_03/_Ato2011-2014/2012/Lei/L12651.htm. [Review date: 26 December 2012] [ Links ].
QUERINO, R. B.; COUCEIRO, S. R. M.; QUEIROZ, L. O.; PENTEADO-DIAS, A. M. 2011. The spatial distribution of Hymenoptera parasitoids in a forest reserve in Central Amazonia, Manaus, AM, Brazil. Brazilian Journal of Biology 71 (4): 865-871. [ Links ]
RIFFEL, C. T.; PRANDO, H. F.; BOFF, M. I. C. 2010. Primeiro relato de ocorrência de Telenomus podisi (Ashmead) e Trissolcus urichi (Crawford) (Hymenoptera: Scelionidae) como parasitoides de ovos do percevejo-do-colmo-do-arroz, Tibraca limbativentris (Stål) (Hemiptera: Pentatomidae), em Santa Catarina. Neotropical Entomology 39 (3): 447-448. [ Links ]
SECRETARIA DO MEIO AMBIENTE DO ESTADO DE SÃO PAULO. 2013. Ecp - consultoria ambiental. Fisionomia de restinga. Available at: www.consultoriaambiental.com.br. (Review date: 18 January 2013). [ Links ]
SEMA. 2011. Refúgio de vida silvestre Banhado dos Pachecos. Available at: www.sema.rs.gov.br. (Review date: 15 September 2011). [ Links ]
SOUTHWOOD, T. R. E. 1978. Ecological methods with particular reference to the study of insect populations.Chapman and Hall, London. 524 p. [ Links ]
TANQUE, R. L.; KUMAGAI, A. F.; FRIEIRO-COSTA, F. A.; SOUZA, B. 2010. Ichneumonidae (Insecta: Hymenoptera) da Reserva do Boqueirão, Ingaí – MG . Revista Brasileira de Zoociências 12 (3): 241-247. [ Links ]
THOMAS, M. B.; WRATTEN, S. D.; SOTHERTON, N. W. 1991. Creation of 'island'; habitats on farm land to manipulate populations of beneficial arthropods: predator densities and emigration. Journal of Applied Ecology (28): 906-917. [ Links ]
TOWNES, H. 1972a. A light-weight Malaise trap. Entomological News 83 (9): 239-247. [ Links ]
TOWNES, H. 1972b. Ichneumonidae as biological control agents. Proceedings Tall Timbers Conference on Ecological Animal Control by Habitat Management 3: 235-248. [ Links ]
WILSON, E. O. 1994. Diversidade da vida. Companhia das Letras, São Paulo. 447 p. [ Links ]
WOLDA, H. 1988. Insect seasonality, why? Annual Review of Ecology and Systematics 19: 1-18. [ Links ]
Received: 13-Aug-2014
Accepted: 12-Aug-2016

