











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroducción
El psílido asiático de los cítricos Diaphorina citri Kuwayama, 1908 (Hemiptera: Liviidae), presenta distribución amplia en áreas tropicales y subtropicales de numerosos países en Asia, América del Sur, América del Norte, África y el Caribe (Halbert y Núñez 2004; Mead y Fasulo 2010) y se considera como una de las principales plagas de los cítricos que se hospeda en unos 25 géneros de la familia Rutaceae (Halbert y Manjunath 2004). Los daños directos causados por D. citri resultan del proceso de alimentación, en donde se remueven grandes cantidades de azúcares, se observa encrespamiento de las hojas y la excreción del exceso de azúcares favorece la formación de fumagina (Capnodium citri Berk. & Desm., 1849). En caso de alta infestación, puede causar la muerte de la yema apical y la caída de las hojas nuevas o yemas terminales (Tsai et al. 2002). Los daños indirectos causados por D. citri se relacionan a su papel como vector de dos bacterias, Candidatus Liberibacter asiaticus y Ca. L. americanus, patógenos asociados a la enfermedad “Huanglongbing” (HLB), la cual produce efectos devastadores en las plantaciones de cítricos (Halbert y Manjunath 2004; Bové 2006; Hall et al. 2013; Qureshi et al. 2009; Grafton-Cardwell et al. 2013).
Además de estos patógenos, la enfermedad HLB se asocia con otras especies del género Candidatus (Ca. L. africanus), presentes en las regiones más frías, como en algunos países de África y Europa y cuya transmisión está asociada con el vector Trioza erytreae (Del Guercio 1918) (Hemiptera: Triozidae) y a dos fitoplasmas (Teixeira et al. 2008; Chen et al. 2009). Además, la distribución de las especies de Candidatus puede estar influenciada por la temperatura, ya que Ca. L. asiaticus y Ca. L. africanus difieren en su sensibilidad a altas temperaturas (Bové 2006; Gottwald 2010).
La presencia del pato-sistema (HLB- D. citri) ha causado un efecto devastador en la citricultura mundial (Belasque Jr. et al. 2010; Llauger et al. 2008) y, especificamente, después de la detección de HLB en el estado de Sao Paulo, en Brasil en 2004, D. citri fue considerada la plaga más importante para la citricultura brasilera (Parra et al. 2010). Resultados de Belasque Jr. et al. (2010) plantean que en plantaciones adultas de cítricos sin medidas de control apropiadas, el HLB puede generar inviabilidad económica de los huertos en aproximadamente de 10 años después de la detección inicial de las primeras plantas infectadas. En Yucatán, México, en huertos de limón persa (Citrus latifoliaTanaka, 1951), con presencia de Ca. L. asiaticus se observó una reducción del 17,31 % en peso y del 18,62 % en volumen de jugo de los frutos colectados en árboles con HLB en comparación con frutos de árboles sanos, implicando una pérdida en producción de 2,4 toneladas/ha (Flores-Sánchez et al. 2015).
La presencia de D. citri está reportada en aproximadamente 56 especies pertenecientes a la familia Rutaceae, incluyendo todas las del género Citrus, aunque se ha registrado que D. citri no necesariamente completa su desarrollo en todas ellas (Aubert 1987). De igual manera, se ha indicado que existe preferencia de D. citri por mirto (Murraya exotica (L.) Jack, 1820) para alimentación y oviposición (Halbert y Manjunath 2004; Teck et al. 2011). Así mismo, estudios de la biología bajo condiciones de laboratorio en Florida, EUA mostraron que, en ausencia de brotes nuevos, las hembras de D. citri detenían la oviposición (Tsai y Parra 2000). Además, se ha observado que en regiones donde las condiciones climáticas son favorables, el único factor limitante para la oviposición es la presencia o ausencia de brotes nuevos, las hembras adultas presentan preferencia para ovipositar en brotes nuevos de aproximadamente 5 mm y el desarrollo de la plaga está sincronizado con el crecimento vegetativo del cultivo (Fernández y Miranda 2005; Padulla 2007).
Los estudios encaminados a evaluar el efecto de resistencia de hospederos a la presencia de D. citri, se han enfocado en ofrecer alternativas de la estrategia de manejo del patosistema HLB-D. citri. La mayoría de estos hospederos estudiados tienen gran importancia para los programas de mejoramiento como porta-injertos s (Albrecht y Bowman 2011), con el objetivo de seleccionar combinaciones de copas comerciales-patrones que sean tolerantes o que muestren algún grado de resistencia a los patógenos de HLB. De la misma manera, los estudios sobre la biología de los insectos nos permiten conocer el comportamiento de una especie de insecto sobre una determinada especie vegetal, así como determinar elementos básicos para el diseño de una estrategia de manejo. Sin embargo, la mayoría de los trabajos sobre la biología de D. citri no contemplan los estudios del desarrollo de la plaga y sus parámetros reproductivos en combinaciones de porta-injertos s y variedades de naranjas de interés comercial (Tsai y Liu 2000; Nava et al. 2007; Pérez-Artiles et al. 2011; Alves et al. 2014). Por lo tanto, los estudios de la biología de D. citri en diferentes combinaciones entre variedades de naranjas comerciales y porta-injertos sirven de base para la selección de nuevas combinaciones que sean menos favorables para el desarrollo de D. citri. De esta manera, este trabajo tuvo como objetivo el estudio de la biología de los parámetros poblacionales de D. citri en combinaciones de porta-injertos y variedades de naranjas, con el fin de definir la influencia de los patrones evaluados sobre la biología y características reproductivas de la plaga cuando se alimentan directamente en los patrones y combinaciones de éstos con copas de naranja.
Materiales y métodos
Cría y mantenimiento de Diaphorina citri
Para el inicio de la cría de D. citri se utilizaron plantas de mirto (M. exotica) e insectos de D. citri provenientes de las crías de laboratorio localizadas en el fondo de defensa de la citricultura Brasilera “Fundecitrus” en Araraquara, Sao Pablo. Las plantas usadas para el mantenimiento de la cría de D. citri y un porcentaje de los insectos, se evaluaron con una frecuencia semestral mediante la técnica molecular de qPCR (reacción en cadena de la polimerasa en tiempo real), con el fin de garantizar que los insectos requeridos para los ensayos estuvieran libres de la bacteria del HLB. Se realizaron podas periódicas para estimular la brotación vegetativa y garantizar la disponibilidad de brotes jóvenes para la sobrevivencia de la plaga. El pie de cría inicial bajo condiciones de laboratorio contó con 20 plantas de mirto de aproximadamente 40 cm de altura y un año de plantadas y 400 adultos de D. citri. Para el mantenimiento de la cría, cada 15 días se colectaron adultos de D. citri con el fin de infestar nuevas plantas que presentaran brotes menores de 0,5 mm. Este pie de cría fue mantenido bajo condiciones de laboratorio durante un año, tiempo requerido para la realización de este estudio.
Las plantas usadas para el mantenimiento de la cría de D. citri se mantuvieron en bolsas plásticas negras de 10 L en una casa de malla, bajo condiciones de humedad y temperatura del ambiente y permanecieron libres de infestación de insectos. Las plantas fueron regadas diariamente y se fertilizaron cada tres semanas con sulfato de amonio (5 gramos por bolsa) y Osmocote (Forth cote, Brasil, proporción 1:1:1 de N-P-K) (3 gramos por bolsa) al inicio del experimento.
Biología de Diaphorina citri en los porta-injertos s Rangpur y Citrumelo ‘Swingle’
Para el estudio de la biología de D. citri en los porta-injertos Rangpur y Citrumelo ‘Swingle’ se utilizaron plántulas de una altura aproximada de 25 cm, con presencia de brotes. Se colocaron las plántulas en vasos de plástico (12 x 3,5 cm) que contenían vermiculita humedecida, con el fin de garantizar la humedad de la plántula. Posteriormente, cada plántula se ubicó dentro de una “jaula” construida a partir de una botella plástica transparente de 2 litros, abierta en la parte superior y cubierta con una tela de organza para permitir la aireación de los insectos. Se utilizaron 12 plántulas para cada tratamiento o porta-injertos y dos parejas de D. citri de 15 días de edad fueron colocadas por plántula. Los adultos fueron sexados bajo un microscopio estereoscópico binocular de acuerdo con la descripción de Mead (1977).
Para homogenizar la disponibilidad de brotes, algunos de ellos fueron removidos, manteniendo apenas uno, con un tamaño ≤ 5 mm. Después de 24 h, los insectos fueron retirados y se contaron los huevos ovipositados en cada plántula con el auxilio de un microscopio estereoscópico binocular. Las evaluaciones incluyeron la viabilidad del huevo, la duración de los estadios huevo y ninfa, y la proporción de sexos de acuerdo al método propuesto por Nava et al. (2007). El experimento se desarrolló en cámaras climatizadas con temperatura de 27 ± 1 °C, 60 ± 10 % de humedad relativa (HR) y fotoperiodo de 14: 10 h luz: oscuridad.
Biología de Diaphorina citri en diferentes combinaciones de porta-injertos s y copas de naranjas
Los tratamientos usados para este experimento fueron plantas de naranja (Citrus sinensis(L.) Osbeck, 1765), de las variedades Hamilim, Valencia y Pera injertadas en Rangpur (Citrus limonia (L.) Osbeck, 1765) y Citrumelo ‘Swingle’ (Citrus paradisi Macfad. cv. Duncan, 1830 x Poncirus trifoliata (L.) Raf., 1838). Se usaron 12 plantas (repeticiones) por hospedero siguiendo el método desarrollado por Alves et al. (2014). Dos parejas de D. citri de 15 días de edad fueron colocadas en cada hospedero. Las plantas fueron cubiertas por una jaula individual confeccionada con tejido de organza (30 x 35 cm diámetro y largo). Después de 24 h individualmente, los insectos se retiraron de la plántula con un aspirador bucal y los huevos se cuantificaron con un microscopio estereoscópico binocular (NSZ 606T/ LED 200x, Coleman, Brasil).
Se hizo un seguimiento diario a las plantas con el fin de evaluar la duración de las etapas de desarrollo de D. citri (huevos, ninfas y ciclo biológico), la fertilidad de los huevos, la supervivencia y la proporción de sexos. Todas las evaluaciones se realizaron en las mañanas (9:00-12:00 m). Este experimento se llevó a cabo dentro de un laboratorio climatizado, con temperatura de 27 ± 1 °C, 60 ± 10 % de humedad relativa (HR) y fotoperiodo de 14: 10 h luz: oscuridad.
Parámetros poblacionales y reproductivos
Para la elaboración de las tablas de vida y la obtención de los parámetros reproductivos de D. citri, se evaluaron los porta-injertos s Rangpur, Citrumelo 'Swingle' y dos combinaciones copa de naranja/ porta-injertos: naranja Valencia/Rangpur y Valencia/Citrumelo ‘Swingle'. El experimento se realizó dentro de un laboratorio con una temperatura de 25 ± 2 °C, humedad relativa de 60 ± 10 % y fotoperiodo de 14: 10 h luz: oscuridad. Las plantas se podaron a una altura de 20 cm para los porta-injertos s y 40 cm para las plantas injertadas. Se utilizaron 20 plantas (repeticiones) por hospedero y cada planta fue infestada con una pareja de adultos de D. citri, con 8-12 h de emergencia. Las plantas junto con la pareja de D. citri se cubrieron con una jaula individual (30 cm de diámetro y 35 cm de largo) confeccionada con malla anti-insectos. Las plantas se revisaron cada tres días, con la ayuda del microscopio estereoscópico, con el fin de evaluar la presencia de huevos y contabilizarlos. En seguida, la pareja de D. citri se trasladaba a una nueva planta con brotes, en donde el conteo de huevos se hizo a los tres días y este procedimiento se repitió hasta la muerte de la hembra. En el caso en que se observaba la muerte del macho de D. citri, éste fue sustituido por otro joven para garantizar la presencia de la pareja durante todo el estudio. Las plantas con huevos se evaluaron diariamente hasta la eclosión de las ninfas. Además, con el fin de reducir el número de plantas requeridas, aquellas en donde se observó la eclosión de las ninfas no se descartaron, sino que se reutilizaron dentro del experimento, luego de una poda para estimular una nueva brotación y garantizar que estuvieran libres de insectos.
Para calcular la longevidad los adultos fueron evaluados desde la emergencia hasta la muerte del último individuo (Rabinovich 1978, 1980; Begon et al. 1988). Con los datos de fecundidad, longevidad y ciclo de vida de D. citri, se construyó una tabla de vida para cada uno de los cuatro tratamientos y se calcularon los parámetros poblacionales como se describen a continuación: x = edad por unidad de tiempo (días), N
x
= total de individuos observados al comienzo de cada etapa, l
x
= proporción de supervivientes a la edad de inicio (N
x
/ N
0
), d
x
= número de muertes entre las edades (l
x
/ l
x + 1
), q
x
= tasa de mortalidad (d
x
/ l
x
), L
x
= individuos que viven entre l
x
/ l
x + 1
, T
x
= tiempo restante a la altura de la muerte (L
X
revertir Σ), e
x
= esperanza de vida (T
x
/ l
x
), m
x
= número de individuos producidos por unidad de tiempo. Para la construcción de la tabla de vida de fertilidad se seleccionó un intervalo de tres días y se calcularon los siguientes parámetros: tasa reproductiva neta
Análisis estadístico
Todos los análisis estadísticos se hicieron con el programa estadístico SAS (SAS Institute Inc. 2012). Los experimentos fueron analizados mediante un diseño experimental completamente al azar, donde cada genotipo estudiado fue considerado un tratamiento. Los datos fueron transformados para reunir las condiciones de normalidad y homogeneidad de varianzas. Los datos de viabilidad de huevos fueron transformados mediante la raíz cuadrada del arco-seno
Resultados y discusión
Biología de Diaphorina citri en los porta-injertos s Rangpur y Citrumelo ‘Swingle’
La duración de todos los estados de desarrollo de D. citri fue mayor en Citrumelo 'Swingle' comparado con Rangpur (Tabla 1). La especie necesitó 17,54 días para completar su ciclo biológico en Citrumelo 'Swingle' y 16,13 días en Rangpur. Estos resultados indican que la duración de los estados de desarrollo del insecto varía dependiendo de la planta hospedera, lo cual coincide con resultados obtenidos por Teck et al. (2011), quienes mostraron diferencias significativas en la duración del ciclo de vida de este insecto en los tres hospederos estudiados (Murraya paniculata, Citrus reticulata y M. koenigii). Sin embargo, otros autores han mostrado que hospederos como C. limonia, C. sunki y M. paniculata no afectaron la duración de las fases de huevo, ninfa y ciclo biológico de D. citri (Nava et al. 2007).
Tabla 1 Media (± desviación estándar) de la duración del desarrollo de Diaphorina citri criada en diferentes hospederos.
| Hospederos | N | Duración (días) (media± desviación estándar) | ||
|---|---|---|---|---|
| Huevo | Ninfas | Huevo- Adulto | ||
| Citrumelo ‘Swingle’ | 12 | 4,27 ± 0,28 a | 13,45 ± 0,33 a | 17,54 ± 0,49 a |
| Rangpur | 12 | 3,75 ± 0,43 b | 12,48 ± 0,72 b | 16,13 ± 0,62 b |
| t | 5,717 | 10,540 | 8,341 | |
| gl | 22 | 22 | 22 | |
| P | < 0,01 | < 0,01 | < 0,01 | |
* Significativo al nivel de 5 % de probabilidad (P < 0,05). Temperatura 27 ± 1 °C, 60 ± 10 % de humedad relativa (HR) y fotoperiodo de 14: 10 h luz: oscuridad. Dentro de cada columna, medias seguidas por las letras diferentes difieren estadísticamente para la prueba t-Student (P > 0,05).
Las diferencias entre estos resultados pueden ser atribuidas a las diferencias en las metodologías utilizadas por los autores, principalmente, en cuanto a la edad de las plantas y también a la cantidad y tamaño de los brotes foliares usados para la alimentación de los insectos. La mayor tasa de viabilidad de huevos (82,56 %) y de viabilidad desde el estadio de huevo hasta el estadio de adulto (70,27 %) para D. citri fueron observadas sobre el tratamiento Rangpur, siendo significativamente diferente de los resultados obtenidos en el tratamiento Citrumelo ‘Swingle’ (Tabla 2).
Tabla 2 Media (± desviación estándar) de sobrevivencia y proporción de sexos de Diaphorina citri criados en diferentes hospederos.
| Hospederos | N | Proporción de sexos * | Viabilidad** | Fertilidad* |
|---|---|---|---|---|
| Rangpur | 12 | 0,62 ± 0,05 a | 70,27 ± 6,96 a | 82,56 ± 4,47 a |
| Citrumelo ‘Swingle’ | 12 | 0,55 ± 0,07 b | 58,97 ± 9,28 b | 75,34 ± 6,56 b |
| t | 5,105 | 3,627 | 7,353 | |
| gl | 22 | 22 | 22 | |
| P | 0,0117* | 0,0377* | 0,00229* |
*Significativo a nivel de 5 % de probabilidad (P ≤ 0,05) para la prueba de Tukey HSD. Temperatura 27 ± 1 °C, 60 ± 10 % de humedad relativa (HR) y fotoperiodo de 14: 10 h luz: oscuridad. Dentro de cada columna, medias seguidas por las letras diferentes difieren estadísticamente para la prueba t-Student (P > 0,05).
Autores como Westbrook et al. (2011) y Borgoni et al. (2014), reportan en P. trifoliata y en algunos de sus híbridos, incluido Citrumelo Swingle la existencia de una resistencia parcial a D. citri. Borgoni et al. (2014), en cuanto a la oviposición, reportan que P. trifoliata 'Rubidoux' tiene una resistencia del tipo de antixenosis frente D. citri. El mecanismo funcional de esta resistencia está relacionado con un conjunto de características que hacen que la planta sea menos atrayente para el insecto. Estos autores no descartan la posible presencia de resistencia tipo antibiosis en algunos de estos genotipos.
Nuestros resultados muestran que la planta hospedera puede influir sobre la viabilidad de los estados de desarrollo de este insecto. Otros estudios también muestran diferencias en la viabilidad de D. citri cuando es criada en diferentes hospederos. Tsai y Liu (2000) observaron una mayor viabilidad promedio (84,5 %) cuando los insectos se alimentaron de Citrus paradisi Macfadyen que cuando lo hicieron sobre C. aurantium L. (68,5 %). De la misma manera, Nava et al. (2007) observaron que la viabilidad del insecto entre los hospederos estudiados fue menor sobre Citrus sunki [Hayata], con una sobrevivencia en la fase de ninfa de apenas 44,6 %.
La proporción de sexos también fue influenciada significativamente por el hospedero (Tabla 2), presentando el mayor valor en Rangpur (0,62). Los valores de proporción de sexos por encima de 0,5 muestran que las hembras predominaron en todos los hospederos, evidenciando la alta capacidad reproductiva de D. citri. Alves et al. (2014) también verificaron la predominancia de hembras en todos los hospederos evaluados, a pesar de que no se hallaron diferencias significativas en la proporción de sexos de los insectos entre los genotipos estudiados.
Biología de Diaphorina citri en diferentes combinaciones de porta-injertos y copas de naranjas
La duración del estado embrionario fue menor en la combinación Pera/Rangpur difiriendo significativamente de Hamilim/Citrumelo ‘Swingle’ que presentó la mayor duración para esta fase. También fueron observadas diferencias significativas en la duración de la fase de ninfa y la duración total del ciclo de vida del insecto cuando estos fueron alimentados en los diferentes hospederos. El ciclo total, desde huevo hasta adulto, de D. citri, fue mayor en la combinación Hamilim/Citrumelo ‘Swingle’ (16,76 días), difiriendo significativamente de la duración del insecto criado en naranja Valencia y Pera independientemente del porta-injertos utilizado.
Cuando se compara la duración de las fases del desarrollo de D. citri en las diferentes combinaciones de copa/porta-injerto, se observó que en la combinación Hamilim/C. ‘Swingle’, el insecto necesitó de mayor tiempo para completar todos sus estados de desarrollo, diferenciándose de manera significativa con los períodos de duración del ciclo biológico y del estado de ninfa para las combinaciones de las variedad Pera y Valencia, en cuanto a la duración del estado de huevo fueron observadas diferencias significativas para la combinación Pera/ Rangpur (Tabla 3).
Para completar los estados de huevo y ninfa, D. citri requirió un mayor tiempo en la combinación Hamilim/Citrumelo ‘Swingle’. En contraste, la duración del estado embrionario fue menor en la combinación Pera/Rangpur (Tabla 3). Así mismo, la duración total del ciclo de vida de D. citri, de huevo a adulto fue mayor en la combinación Hamilim/C. ‘Swingle’ (16,76 días), difiriendo significativamente de los valores obtenidos en la variedad Valencia independientemente del porta-injertos utilizado.
Tabla 3 Media (± desviación estándar) da duración do desarrollo de Diaphorina citri criadas en diferentes copas sobre porta-injertos s Rangpur y Citrumelo ‘Swingle’ en condiciones controladas.
| Copa/porta-injertos | N | Duración (días) | ||
|---|---|---|---|---|
| Huevos | Ninfas | Ciclo biológico (huevo-adulto) | ||
| Hamilim/C. ‘Swingle’ | 12 | 4,52 ± 0,473a | 12,25 ± 0,76 a | 16,76 ± 0,42 a |
| Hamilim/Rangpur | 12 | 4,33 ± 0,44 ab | 12,08 ± 0,75 ab | 16,37 ± 0,45 a |
| Valencia/C. ‘Swingle’ | 12 | 4,25 ± 0,45 ab | 11,20 ± 0,78 c | 15,12 ± 0,84 b |
| Pera/C. ‘Swingle’ | 12 | 4,16 ± 0,29ab | 11,33 ± 0,41 bc | 15,50 ± 0,52 b |
| Valencia/Rangpur | 12 | 4.14 ± 0,41 ab | 10,28 ± 0,93 d | 14,90 ± 0,93 b |
| Pera/Rangpur | 12 | 3,91 ± 0,59 b | 11,16 ± 0,37 c | 15,09 ± 0,79 b |
| F | 2,37 | 12,38 | 14,31 | |
| gl | 5,66 | 5,66 | 5,66 | |
| P | < 0,01 | < 0,01 | < 0,01 | |
* Significativo al nivel de 5 % de probabilidad (P ≤ 0,05) para la prueba de Tukey HSD. Médias seguidas por la misma letra no difiereen estadísticamente. Temperatura 27 ± 1 °C, 60 ± 10 % de humedad relativa (HR) y fotoperiodo de 14: 10 h luz: oscuridad.
El intervalo total de viabilidad de los huevos fue de 82,03 a 78,96 %, siendo mayor en Valencia/Rangpur, con diferencias significativas para los valores en Valencia/Citrumelo ‘Swingle’ y Hamilim/Citrumelo ‘Swingle’ (Tabla 4). Con respecto a la viabilidad de huevo a adulto, se observó que la combinación Pera/Rangpur fue la combinación donde D. citri tuvo la mayor supervivencia (71,07 %), con diferencias significativas cuando los insectos fueron criados en Valencia/Citrumelo ‘Swingle’, Hamilim /Rangpur y Hamilim/Citrumelo ‘Swingle’ (Tabla 4).
Tabla 4 Media (± desviación estándar) de viabilidad (%) y la razón sexual (RS) de D. citri criados en diferentes hospederos.
| Hospederos | N | Proporción sexos* | Viabilidad (%)* | ||
|---|---|---|---|---|---|
| Huevos | Huevos- adultos | ||||
| Pera/Rangpur | 12 | 0,65 ± 0,03 a | 81,69 ± 5,40 a | 71, 07 ± 6,05 a | |
| Hamilim /Rangpur | 12 | 0,64 ± 0,05 ab | 78,96 ± 7,60 abc | 62,93 ± 7,91 bc | |
| Valencia/Rangpur | 12 | 0,59 ± 0,06 b | 82,03 ± 4,27 a | 66,11 ± 6,10 ab | |
| Pera/Citrumelo ‘Swingle’ | 12 | 0,59 ± 0,06 b | 79,92 ± 4,10 ab | 64,52 ± 7,06 ab | |
| Valencia/Citrumelo ‘Swingle’ | 12 | 0,50 ± 0,05 c | 74,73 ± 4,81 bc | 60.53 ± 5,20 bc | |
| Hamilim/Citrumelo ‘Swingle’ | 12 | 0, 48 ± 0,06 c | 73,66 ± 4,18 c | 56,16 ± 6,50 c | |
| F | 23,95 | 5,534 | 7,22 | ||
| gl | 5,66 | 5,66 | 5,66 | ||
| P | < 0,01 | < 0,01 | < 0,01 | ||
* Significativo al nivel de 5 % de probabilidad (P ≤ 0,05) para la prueba de Tukey HSD. Médias seguidas por la misma letra no difiereen estadísticamente. Temperatura 27 ± 1 °C, 60 ± 10 % de humedad relativa (HR) y fotoperiodo de 14: 10 h luz: oscuridad.
Los insectos que fueron criados en naranja Hamilim injertada en Citrumelo ‘Swingle’ presentaron los menores valores de viabilidad de huevos y para el período de huevo a adulto (Tabla 4). Los resultados muestran que esta combinación, fue la que menos favoreció el desarrollo y la reproducción de D. citri y son semejantes a los registrados por Alves et al. (2014). Estos autores encontraron que la naranja Hamilim fue el tratamiento que menos favorecía el desarrollo y la viabilidad de D. citri. Además, Souza et al. (2012) destacaron que las diferencias en el tiempo de desarrollo de D. citri y la viabilidad en diferentes variedades pueden estar relacionadas con la concentración de nitrato en la savia da planta. Estos autores observaron diferencias mayores al 20 % en el contenido de nitrato en la savia de la naranja Hamilim en comparación con la de naranja Valencia. Otros autores, Teck et al. (2011), también sugieren las diferencias encontradas con respecto a la duración de las fases de desarrollo de D. citri pueden estar relacionadas con la calidad nutricional de la planta hospedera, principalmente en la calidad de la savia y la concentración de aminoácidos disponibles. Las diferencias en la sobrevivencia y el desarrollo de los insectos también pueden ser causadas por la ingestión de compuestos tóxicos y/o por la insuficiencia nutricional de la planta hospedero (Vendramim y Guzzo 2012).
La proporción de sexos de D. citri presentó diferencias significativas entre los hospederos. La combinación que más favoreció la emergencia de las hembras fue Pera/Rangpur, lo que influyó a su vez en el incremento da tasa neta de reproducción (R 0) de D. citri en este hospedero. Para la combinación Hamilim/Citrumelo ‘Swingle’, se observaron valores de la proporción de sexos, inferiores a 0,5, lo que indicó que este hospedero disminuyó la emergencia de hembras (Tabla 4).
Longevidad y fecundidad de los adultos de Diaphorina citri
Los valores de la longevidad media de las hembras presentaron diferencias significativas (F = 6,32; gl = 3,76; P < 0,01) entre los hospederos evaluados La menor longevidad media de la hembra de D. citri fue observada en Citrumelo ‘Swingle’ (34,25 días), diferenciándose significativamente de los valores de longevidad obtenidos para las hembras de D. citri en los otros tratamientos (Fig. 1). Tsai y Liu (2000) observaron que los valores medios de longevidad de D. citri fueron de 39,7 días a 25 °C en M. paniculata. Además, otro estudio sobre la longevidad media de los adultos fue obtenida por Chavan y Summanwar (1993) en limón mexicano, Citrus aurantifolia(Christm.), en donde se observó una longevidad aproximada de 20 días en el verano y 190 días para la época de inverno. Similarmente, Fung y Chen (2006) observaron que la longevidad de adultos de D. citri sobre M. paniculata se afecta por cambios en la temperatura, con reducción de un 50 % cuando la temperatura aumentaba desde los 20 ºC hasta los 28 ºC.
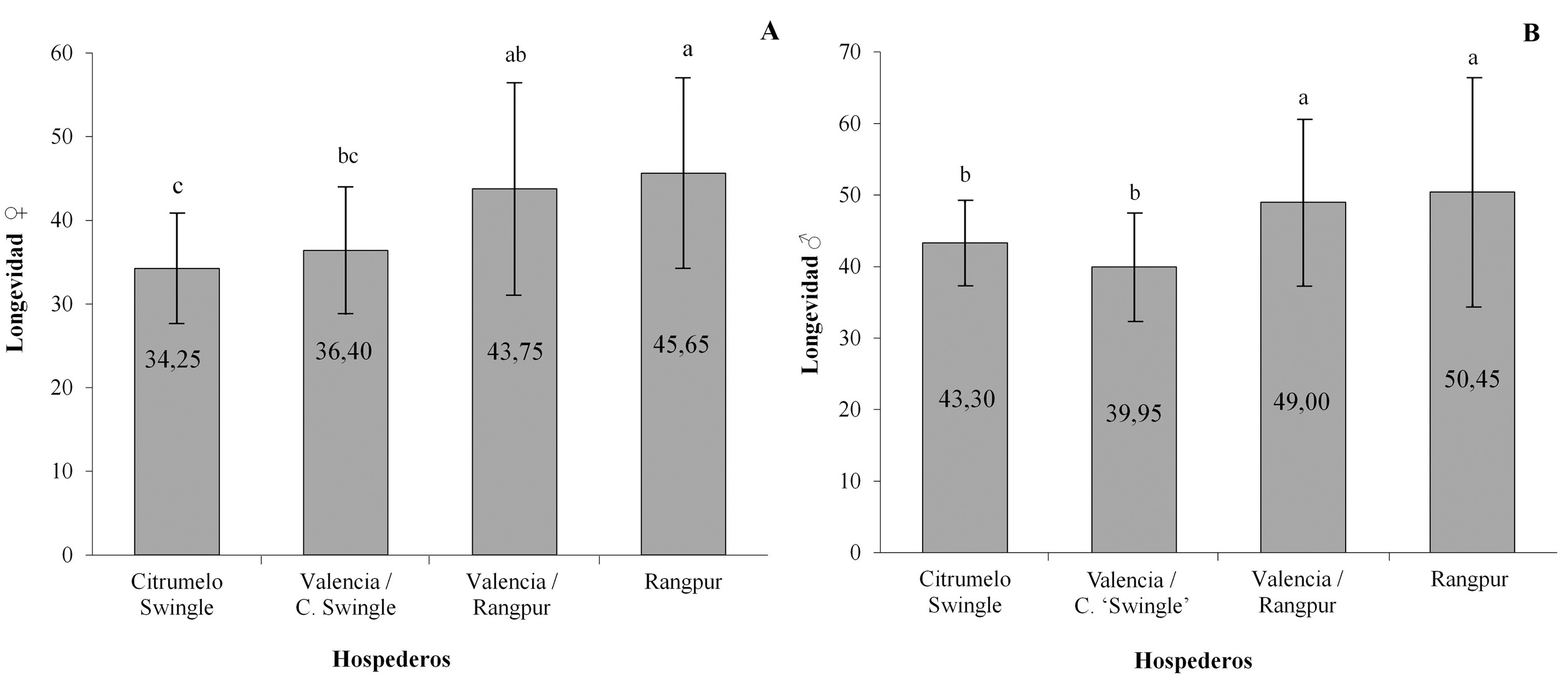
Figura 1 Longevidad media (días) de (A) hembras (L ♀) y (B) machos (L ♂) de Diaphorina citri criados en diferentes hospederos. Temperatura de 25 ± 2 °C, 60 ± 10 % de humedad relativa (HR), y fotoperiodo de 14: 10 h luz: oscuridad. Medias seguidas por la misma letra en la columna no difieren entre sí (Prueba de Tukey, P > 0,05).
El mayor periodo de pre-oviposición fue observado en Citrumelo ‘Swingle’ (14,10 días), con diferencias significativas respecto a los otros hospederos evaluados (F = 8,07; gl = 3,76; P < 0,01).
Para el período de oviposición se observaron diferencias significativas (F = 8,92; gl = 3,76; P < 0,01), en las combinaciones Valencia/Rangpur y Valencia/ Citrumelo ‘Swingle’, donde la naranja Valencia injertada en Citrumelo ‘Swingle’, D. citri presentó un período de oviposición más corto. De los hospederos evaluados, Citrumelo ‘Swingle’ es el hospedero que menos favoreció la reproducción de D. citri, observándose en él periodo de pre-oviposición más largo y de oviposición más corto (Fig. 2), así como la menor oviposición y una tasa líquida de reproducción menor que 100, sugiriendo que entre los hospederos evaluados este hospedero es el menos indicado para la reproducción de D. citri.
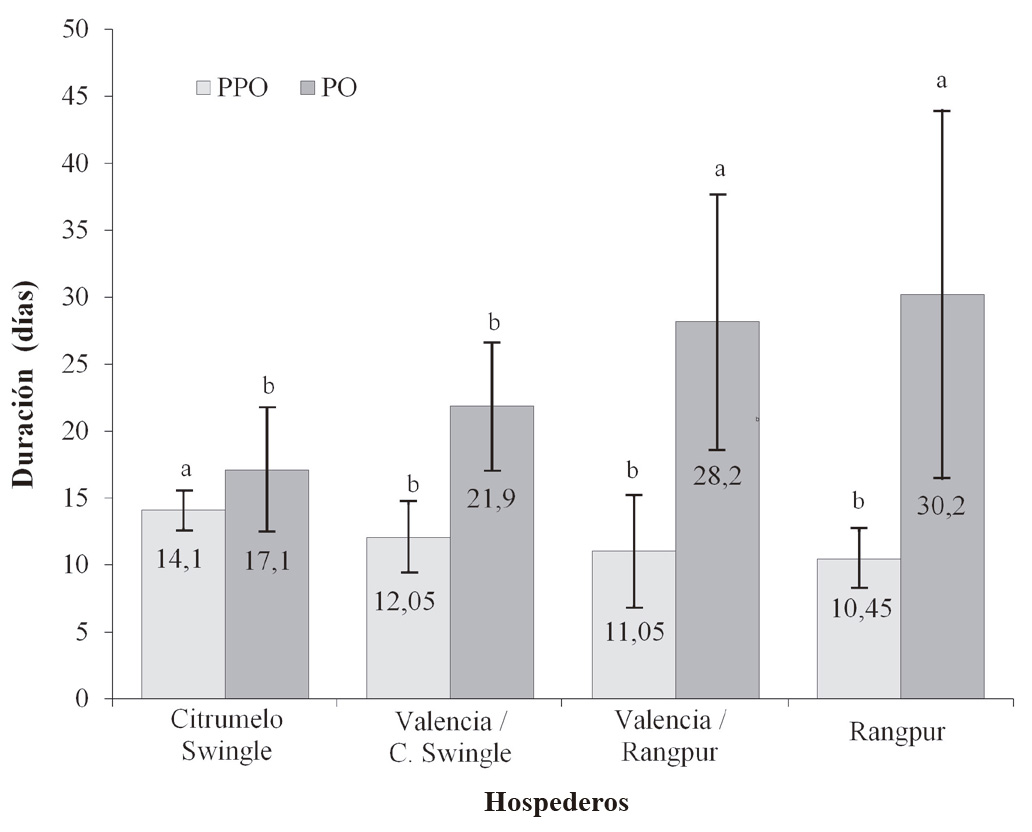
Figura 2 Duración de los períodos de pre-oviposición (PPO) y del período de oviposición (PO) de D. citri criados en diferentes hospederos. Temperatura de 25 ± 2 °C, 60 ± 10 % de humedad relativa (HR), y fotoperiodo de 14: 10 h luz: oscuridad. Medias seguidas por la misma letra en la columna no difieren entre sí (Prueba de Tukey, P > 0,05).
La media del período de pre-oviposición en el presente estudio fue superior a los diez días en todos los hospederos evaluados lo cual concuerda con lo encontrado por Nava et al. (2007), quienes obtuvieron valores cercanos a los diez días para el período de pre-oviposición en todos los hospederos analizados. En contraste, Alves et. al. (2014) registraron valores menores para el periodo de pre-oviposición que los encontrados en este trabajo, y que para los registrados por Nava et al. (2007). Alves et al. (2014) atribuían estas diferencias a la presencia o ausencia de brotes nuevos y/o de hojas maduras en los hospederos usados en los experimentos, ya que según los trabajos de Uechi y Iwanami (2012), la presencia de brotes nuevos favorece a maduración de los ovarios de D. citri.
La fecundidad promedio (huevos/hembra de D. citri) fue significativamente diferente entre los hospederos evaluados (F = 3,601; gl = 3,76; P < 0,01), siendo Rangpur el hospedero donde D. citri presentó mayores valores de la fecundidad (254,15 huevos). Los menores valores de fecundidad se observaron en copa de Valencia injertada en Citrumelo Swingle, pero sin diferencias significativas con la combinación Valencia /Rangpur (Fig. 3). Estos resultados concuerdan con lo reportado por Borgoni et al. (2014), quienes encontraron que Citrumelo Swingle presentó la viabilidad más baja de los huevos y una reducción en la fecundidad de las hembras criadas sobre este hospedero.
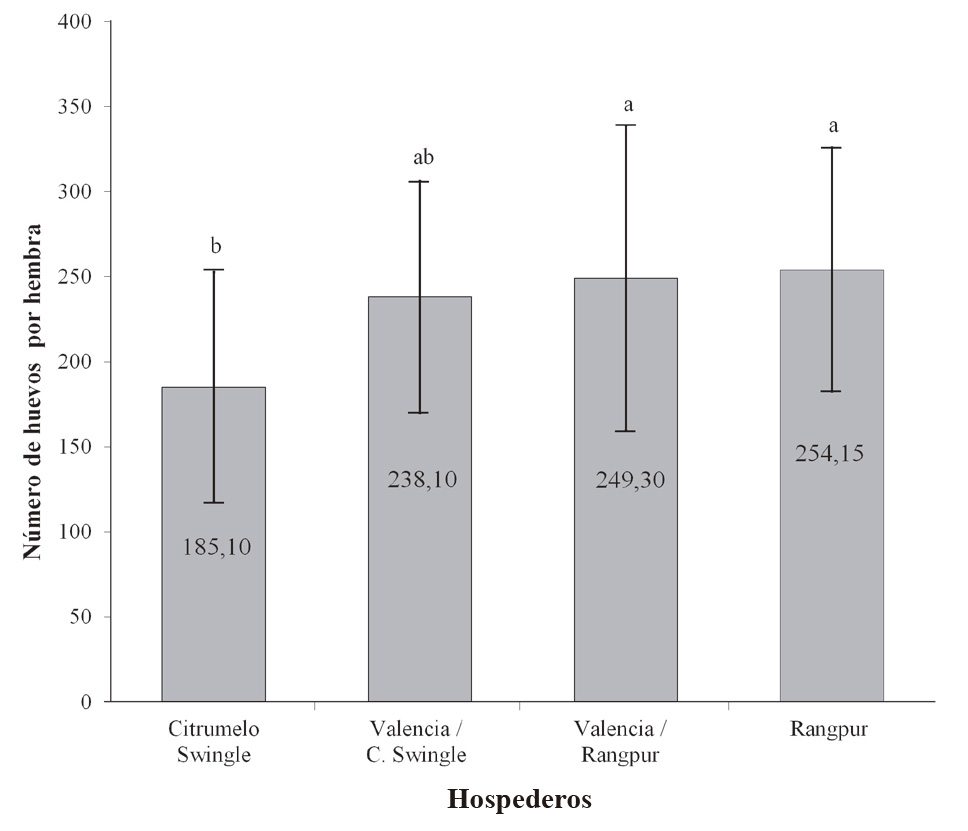
Figura 3 Media de oviposición de D. citri en diferentes hospederos. Temperatura de 25 ± 2 °C, 60 ± 10 % de humedad relativa (HR), y fotoperiodo de 14: 10 h luz: oscuridad. Medias seguidas por la misma letra en la columna no difieren entre sí (Prueba de Tukey, P > 0,05).
El valor promedio de huevos por hembra obtenido en todos los hospederos estuvo dentro del intervalo de 185 a 300 huevos/hembra encontrado por Pande (1971). Nava et al. (2007) mostraron diferencias significativas en la fecundidad media de las hembras de D. citri y reportaron medias de oviposición en Rangpur de 265,1 huevos/hembra, valores semejantes a lo obtenido en este trabajo. Tsai y Liu (2000) sugieren que las variaciones en la fecundidad y la longevidad de este insecto están influenciadas por los hospederos, así como también por los biotipos de D. citri.
Oviposición y curva de sobrevivencia de Diaphorina citri
La Figura 4 muestra el comportamiento de la oviposición de D. citri y la curva de supervivencia de D. citri en cada hospedero. El pico de oviposición se alcanzó entre los 27 y los 33 días a partir del inicio de la oviposición (Fig. 4). De acuerdo con los principales tipos de curvas de supervivencia que son propuestos en la literatura, la curva obtenida para D. citri, en las condiciones de este experimento, se asemeja una curva de tipo I (Fig. 4), caracterizada porque la mayor mortalidad natural ocurre en los individuos más viejos de la población (Southwood y Henderson 2000; Rabinovich 1978). Durante la época de reproducción, no se observó mortalidad durante los primeros 12 días y a partir de este día comienzan a morir con un porcentaje muy bajo.
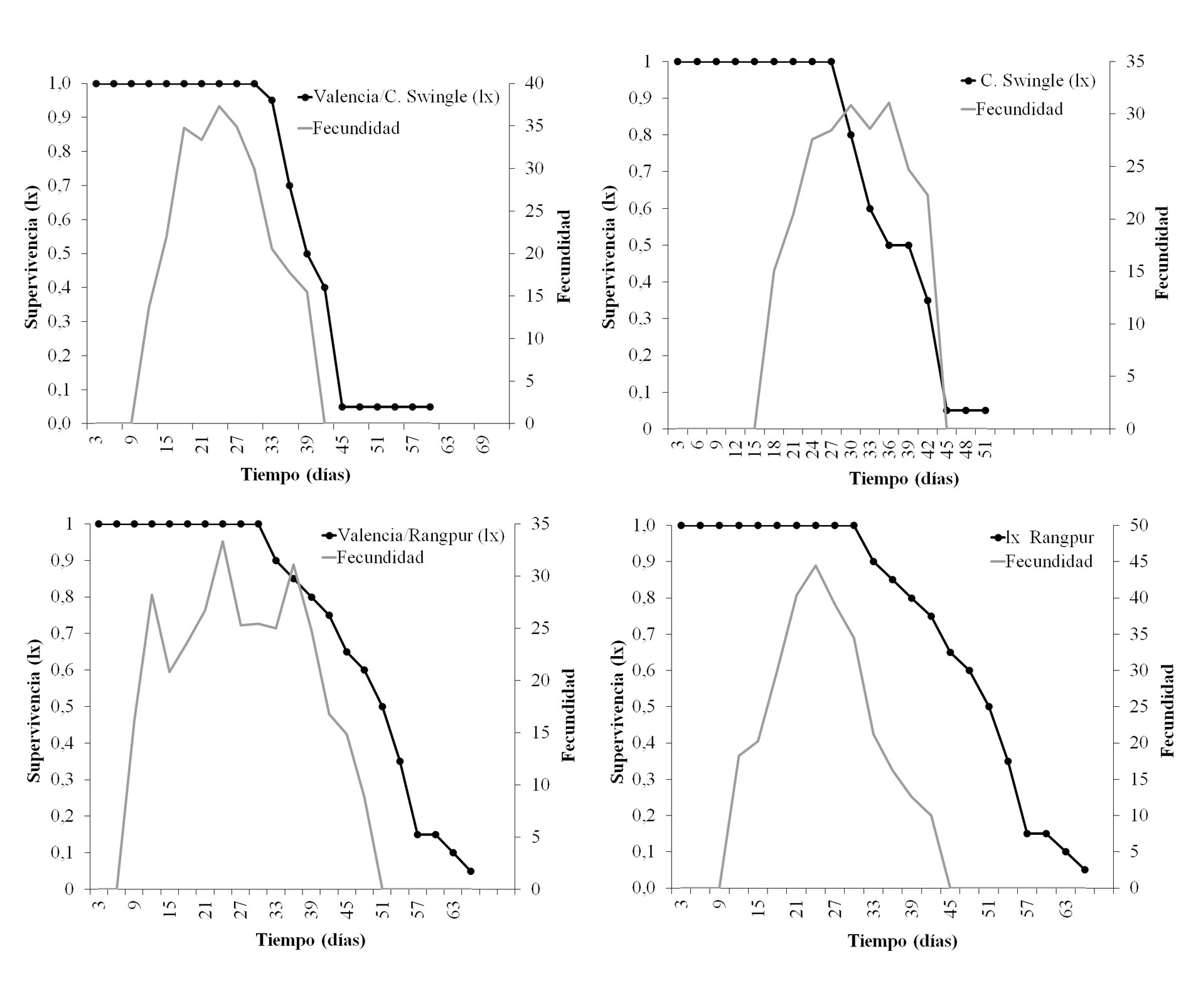
Figura 4 Oviposición y curva de sobrevivencia de Diaphorina citri criadas en diferentes hospederos. Temperatura de 25 ± 2 °C, 60 ± 10 % de humedad relativa (HR), y fotoperiodo de 14: 10 h luz: oscuridad.
Palomares-Pérez et al. (2015) obtuvieron una curva de sobrevivencia tipo II para las poblaciones de D. citri en el verano, pero en ella se incluyeron los datos de los estados inmaduros de D. citri, resultados que difieren del presente trabajo, en donde solamente se incluyeron los datos de los insectos adultos. Para las poblaciones observadas durante la época de invierno, los autores describieron una curva de sobrevivencia do tipo IV, donde la mortalidad de las primeras fases de desarrollo se mantuvo relativamente constante. Rabinovich (1980) plantea que la curva tipo II afecta fundamentalmente a individuos jóvenes de una población.
Correlación de la fecundidad total con periodos de pre-oviposición, oviposición y longevidad
La fecundidad total de las hembras (oviposición total) presentó una correlación positiva y significativa con la longevidad y con el período de oviposición. El período de pre-oviposición estuvo correlacionado de forma negativa con el período de oviposición, mostrando que las hembras que tienen un mayor período de pre-oviposición, tenían el menor período de oviposición. Sin embargo, no se observó correlación significativa del periodo de pre-oviposición con la longevidad (Tabla 5). Estos resultados difieren de los resultados obtenidos por Ma y Wang (2001) en China, quienes observaron que hembras con un período de pre-oviposición más corto presentaban menor longevidad.
Tabla 5 Correlación de la fecundidad (Pearson) total de Diaphorina citri con periodos de pre-oviposición, oviposición y longevidad.
| Factor de correlación | PO | LF | LM | Fecundidad total |
|---|---|---|---|---|
| Periodo de pre-oviposición (PPO) | -0,38189** | -0,20516 | -0,10846 | 0,10269 |
| Período de oviposición (PO) | 1,00 | 0,77979** | 0,66518** | 0,39011** |
Para cada variable analizada en la primera fila se presentan los valores del índice de correlación (r). Valores de r en itálicas corresponden a correlaciones estadísticamente significativas. (**): Altamente significativo (P ≤ 0,01). Longevidad de la hembra (LF); Longevidad del macho (LM).
Parámetros poblacionales
Utilizando la tabla de vida de fertilidad, fue posible determinar el potencial de crecimiento de D. citri en diferentes hospederos. La mayor tasa neta de reproducción (R 0) fue obtenida en Rangpur (160,57), este valor indica que el insecto puede incrementar su población en 160,57 hembras en cada generación cuando es mantenido en este hospedero. Los valores de la tasa neta de reproducción (Ro) presentaron diferencias significativas entre los tratamientos Valencia/Citrumelo ‘Swingle’ y Valencia/ Rangpur (Tabla 6), sugiriendo que el porta-injertos podría estar influenciando en este parámetro. Los valores R 0 alcanzaron valores mayores a 100 en los tratamientos Valencia/Citrumelo ‘Swingle’, Valencia/ Rangpur y Rangpur, indicando un crecimiento de la población en esos tratamientos, según consideraciones teóricas de Southwood y Henderson (2000) y Rabinovich (1980). En contraste, el valor Ro en el tratamiento Citrumelo ‘Swingle’ fue el menor al resto de hospederos, lo que sugiere que este hospedero es menos favorable para la reproducción de D. citri. El tiempo medio de una generación (T) de D. citri fue menor en Rangpur (31,54 días) y en Valencia injertada en Rangpur (31,89 días), diferenciándose de los tratamientos Citrumelo ‘Swingle’ y da naranja Valencia injertada en Citrumelo ‘Swingle’ (Tabla 6).
Tabla 6 Parámetros poblacionales de D. citri criados en diferentes hospederos en condiciones controladas. Temperatura 25 ± 2 ºC, Humedad relativa (HR) 70 ± 10 % y fotoperiodo de 14: 10 h luz: oscuridad.
| Hospederos | Parámetros | ||||
|---|---|---|---|---|---|
| R o | T(G) | r m | λ | DT | |
| Citrumelo ‘Swingle’ | 93,27a* | 35,78a** | 0,1268b* | 1,135b* | 5,46a* |
| Valencia/Citrumelo ‘Swingle’ | 114,65b* | 34,54c** | 0,1396a* | 1,149a* | 4,95b* |
| Valencia/ Rangpur | 126,7c** | 31,89b** | 0,1517c** | 1,163ac** | 4,56b** |
| Rangpur | 160,57d** | 31,54b** | 0,1469d** | 1,158d** | 4,71b** |
** Altamente significativo (P ≤ 0,01) y * significativo al nivel de 5 % de probabilidad (P ≤ 0,05). En las columnas las medias seguidas por la misma letra no diferen estadísticamente para la prueba t- Student.
Los valores de lambda (λ) fueron superiores a la unidad en todos los hospederos, lo que implica un crecimiento de población. En Citrumelo ‘Swingle’ fue donde la población tuvo el menor crecimiento, encontrándose el menor valor de lambda (Tabla 6). El tiempo mínimo para duplicar la población fue de 4,56 días en Valencia/Rangpur y el tiempo máximo de 5,46 días en Citrumelo ‘Swingle’. En estudios realizados por Liu y Tsai (2000) en Florida, EUA, se obtuvieron menores tiempos de duplicación para poblaciones de D. citri, con un valor de 3,5 días a una temperatura de 28 °C. En general, los parámetros poblacionales en condiciones controladas representan el potencial reproductivo de una especie para enfrentar el ambiente y, por tanto, constituyen elementos básicos para a interpretación da dinámica de una población (Rabinovich 1980).
Conclusiones
Los porta-injertos estudiados influyeron en los parámetros poblacionales y reproductivos de D. citri, especialmente los parámetros relacionados con el período de oviposición y la fecundidad. El menor tiempo para completar el ciclo biológico de D. citri fue de 15,08 días en la combinación Pera/ Rangpur. El limón Rangpur y la combinación Valencia/Rangpur fueron los hospederos que favorecieron la reproducción de D. citri, con los más altos valores de tasa neta de reproducción de 160,57 y 126,7, respectivamente. La mayor tasa intrínseca de crecimiento fue obtenida en Valencia/Rangpur (0,1517). Citrumelo 'Swingle' y Hamilim/ Citrumelo 'Swingle' fueron los tratamientos que afectaron el desarrollo y reproducción de D. citri, con los menores valores de supervivencia en la combinación Hamilim/Citrumelo 'Swingle' de 56,16 %. La longevidad de la hembra (0,23440; P = 0,03649) se correlacionó de forma positiva y significativa con la oviposición total. Las plantas hospederas estudiadas mostraron tener efecto sobre el desarrollo, longevidad y la reproducción de D. citri. Estos estudios son importantes en la búsqueda de genotipos o combinaciones de porta-injertos s y copas de interés comercial, que sean poco favorables a desarrollo de D. citri, para disminuir la población del insecto vector y en consecuencia la propagación de la enfermedad.

