











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae), es una plaga invasiva altamente polífaga originaria del sudeste asiático (Walsh et al. 2011). Esta especie se detectó por primera vez en el sur de Europa (España e Italia) y Norte América (California) en 2008 (Cini et al. 2012; Bolda et al. 2010). En 2012, se hallaba en la costa meridional y sureste de Francia, Portugal, Alemania, oeste de Eslovenia, Suiza, Croacia, Austria y Bélgica. Al presente ocupa prácticamente toda Europa (EPPO 2015) y se ha detectado recientemente en Sudamérica, en Brasil, atacando frutos de fresa (Silva do Santos 2014) y arándano (Ribeiro-Vilela y Mori 2014). A diferencia de otras especies del género, D. suzukii posee un ovipositor aserrado que le permite atacar frutos sanos, en envero y maduros (Walsh et al. 2011). Las larvas se alimentan y desarrollan dentro de los frutos favoreciendo la colonización de hongos y otros patógenos, y ocasionando su devaluación. Además, debido a que los daños iniciales son difícilmente apreciables a simple vista, su presencia puede resultar especialmente preocupante tras la cosecha, en las plantas de procesado, ya que pueden pasar desapercibidos y provocar el rechazo de partidas en destino con la consiguiente pérdida económica y de fiabilidad en el mercado. Este díptero causa importantes pérdidas económicas en la producción de frutos rojos en América del Norte (Goodhue et al. 2011) y Europa (Lee et al. 2011) donde puede alcanzar el 80 % en fresa y el 30-40 % en cultivos de arándano, mora y frambuesa. Se han llegado a registrar pérdidas de producción del 80-100 % debido a D. suzukii en parcelas de fresa y cereza en el sur de Europa, específicamente en España, (Sorribas y Lekinberri 2013). Desde la llegada de D. suzukii a Europa y Norte América, se ha avanzado en el estudio de artrópodos benéficos, tanto comerciales como presentes en los diferentes cultivos, que pueden incidir en las poblaciones de este díptero. Sin embargo, aún se desconoce, en gran medida, el potencial de acción de la mayoría de ellos. Se ha constatado que varias especies de parasitoides pueden actuar sobre la plaga. Los datos que se tienen hasta la fecha apuntan fundamentalmente a tres especies de este grupo de enemigos naturales: por un lado Pachycrepoideus vindemmiae (Rondani, 1875) (Hymenoptera: Pteromalidae) y Trichopria drosophilae Perkins, 1910 (Hymenoptera: Diapriidae) que parasitan pupas, y por otro Leptopilina boulardi Barbotin et al., 1979 (Hymenoptera: Figitidae) que parasita larvas (Sorribas y Lekinberri 2013; Arnó et al. 2015). En Italia se han realizado estudios con L. heteronoma (Thompson, 1862) llegando a índices de parasitismo del 60 % (Arnó et al. 2015). Entre los depredadores, se ha constatado la presencia de Orius laevigatus (Fieber, 1860), Cardiasthetus nazarenus Reuter, 1884 y C. fasciiventris (Garbiglietti, 1869) (Hemiptera: Anthocoridae) en fresón y frambuesa infectados por D. suzukii y Dicyphus tamaninii Wagner, 1951 (Hemiptera: Miridae) en Solanum luteum (Arnó et al. 2013). Además, los estudios preliminares en laboratorio han mostrado una escasa actividad de control de Orius majusculus (Reuter, 1879) y O. laevigatus sobre huevos de D. suzukii, aunque estos depredadores y Anthocoris nemoralis (Fabricius, 1794) pueden atacar pupas, adultos y larvas en laboratorio (Malagnini et al. 2014; Cuthbertson et al. 2014). Asimismo, ensayos preliminares indican que el dermáptero Labidura riparia (Pallas, 1773) (Dermaptera: Labidurdae) puede alimentarse de larvas y pupas de D. suzukii reduciendo significativamente su población (Gabarra et al. 2015). Finalmente, algunos ensayos ponen de manifiesto que el estafilínido Dolatia coriaria (Kraatz, 1856) es capaz de depredar larvas y reducir en un 50 % la infestación en frutos (Renkema et al. 2015). Este trabajo analiza el potencial depredador sobre huevos de D. suzukii de algunas de las especies de hemípteros comerciales más ampliamente utilizadas para el control de plagas en frutos rojos y otros cultivos hortícolas.
Materiales y métodos
Los hemípteros seleccionados para el estudio procedieron de productos comerciales utilizados en el control de importantes plagas agrícolas en el sur de Europa: Thripor-L®, O. laevigatus y Nesibug 500®, Nesidiocoris tenuis de Koppert Biological Systems; Almería, España. Anthocontrol 500®, A. nemoralis y Macrocontrol 500®, Macrolophus pygmaeus de Agrobio S.L.; Almería, España. Se utilizó como soporte de oviposición frutos de arándano comerciales de la variedad “Biloxi” (Fres Doñarosa; Superexport Cia. Agraria S.L.; Huelva, España) ya que este fruto permite un conteo de huevos de D. suzukii más sencillo y fiable que otros frutos rojos (Lee et al. 2011). Se llevaron a cabo tres tratamientos: frutos con puesta + depredador, fruto sin puesta + depredador (control) y fruto con puesta sin depredador (control). Un día antes del comienzo del ensayo (24 h), los frutos de arándanos se ofrecieron a grupos de machos y hembras de D. suzukii en jaulas de 0,3 m3 (BugDorm® 1; Bio-Quip Products Inc., Rancho Rodríguez, CA; EEUU). Los adultos empleados provinieron de una población establecida y mantenida en el Laboratorio de Entomología del centro IFAPA “Las Torres-Tomejil” (Alcalá del Río, Sevilla; España) a partir de larvas recolectadas en frambuesas de cultivos infestados de Huelva (sur de España) en febrero de 2014. Posteriormente, el número de huevos por fruto fue contado con la ayuda de una lupa binocular (x20) y los frutos se repartieron equitativamente entre control y tratamiento (con depredadores) según el número de huevos (un promedio de 7,73 ± 0,76 y 7,95 ± 0,79 huevos para control y tratamiento, respectivamente). Cada uno de los frutos se aisló en placas Petri junto a cuatro adultos de cada especie de depredador seleccionada para el tratamiento y sin depredadores para el grupo control, sellando la placa con película de laboratorio para evitar fugas. Paralelamente, del mismo modo, se estableció un segundo control consistente en un fruto sin puesta con cuatro adultos de cada especie (excepto O. laevigatus, del que no se pudieron obtener datos de supervivencia por no disponer de material biológico). Este segundo control se estableció con el propósito de detectar posibles diferencias en la tasa de supervivencia que podrían indicar que los depredadores consiguen alimentarse de huevos (Malagnini et al. 2014). En total se establecieron 24 repeticiones por tratamiento y especie. Los insectos y frutos se mantuvieron en cámara de ambiente controlado: 22 ± 1 °C, 65 ± 5 % HR, y 16:8h (L:O) de fotoperiodo. Al cabo de 72 h, se retiraron los insectos anotando el número de depredadores supervivientes. Los frutos se incubaron en las condiciones indicadas arriba durante 21 días, registrando el número de adultos emergidos por fruto y tratamiento. Al no cumplirse las condiciones de linealidad de los residuos para el análisis paramétrico, el número de adultos emergidos por fruto se comparó con el del tratamiento control utilizando un modelo lineal generalizado (GLZ) con el paquete estadístico R 3.2.2. GLZ en R utiliza el valor del estadístico de “Wald” (‘‘z’’) y Pr ([|z|]) para analizar el efecto de cada factor sobre la variable respuesta testando la hipótesis de que el parámetro correspondiente (coeficiente de regresión) toma el valor 0 (Crawley 2005). En el GLZ se incluyó al número adultos emergidos como variable dependiente ajustado a una distribución Poisson con una función vínculo log, y el tratamiento (depredador/no depredador) como factor. El número de depredadores vivos y muertos en contacto con frutos con puesta se comparó con el de los depredadores en contacto con fruto sin puesta utilizando nuevamente un GLZ. En el modelo se incluyó al número de depredadores vivos y muertos como variable dependiente ajustado a una distribución binomial con una función vínculo logit y el tratamiento (puesta/no puesta) como factor.
Resultados y discusión
En todos los casos el tratamiento con depredadores produjo un número menor de adultos emergidos por fruto con respecto a su grupo control aunque sólo en el caso de A. nemoralis (Fig. 1) se detectaron diferencias significativas (Wald = -3,65; P < 0,001) emergiendo < 50 % de moscas (2,48 ± 0,43 moscas/fruto) que en el control (4,52 ± 0,64 moscas/fruto). Para O. laevigatus, N. tenuis y M. pygmaeus no se detectaron diferencias significativas en el promedio de adultos emergidos por fruto (Wald = 1,19, -1,84 y -1,30, respectivamente; P > 0,05). En todos los casos, el número de depredadores vivos fue mayor en el tratamiento (frutos con puesta) que en el control (frutos sin puesta). En términos de supervivencia, sólo se encontraron diferencias significativas entre el número de depredadores vivos y muertos del tratamiento de N. tenuis con respecto al grupo control (Wald = 1,99; P < 0,05; 30,36 ± 4,01 % y 12.5 ± 3,18 %, respectivamente). Para A. nemoralis y M. pygmaeus no existieron diferencias significativas en las tasas de supervivencia (Wald = -1,73; P > 0,05 y Wald = 168; P > 0,05, respectivamente) siendo éstas del 83,3 ± 3,3 %, y el 70,0 ± 6,9 para tratamiento y control, respectivamente, en el caso de A. nemoralis y del 58,33 ± 8,06 % y el 42,50 ± 5,34 % en el de M. pygmaeus (Fig. 2).
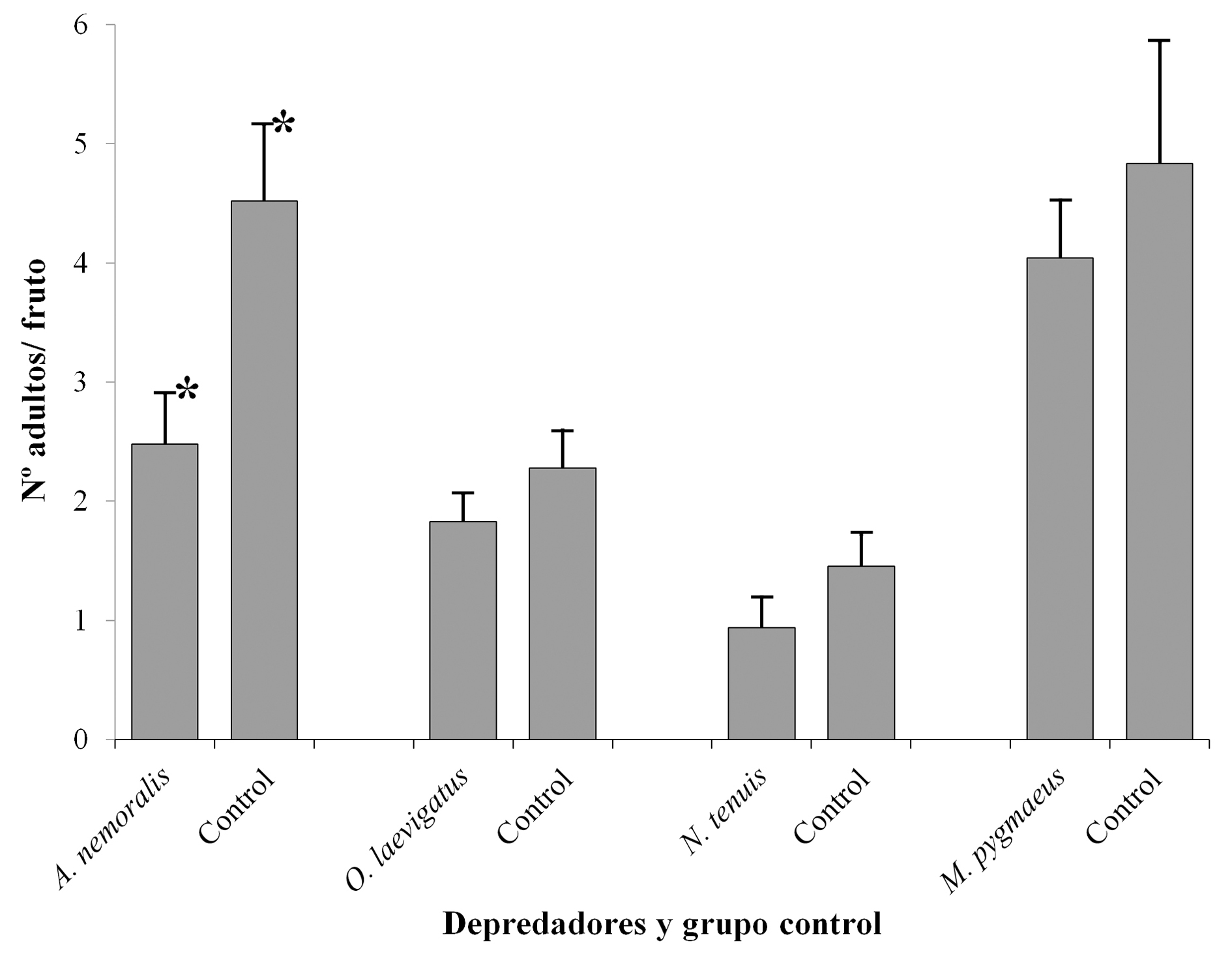
Figura 1 Número medio de adultos emergidos por fruto para cada especie de depredador y su grupo control correspondiente. El símbolo * indica diferencias significativas (P < 0,05).
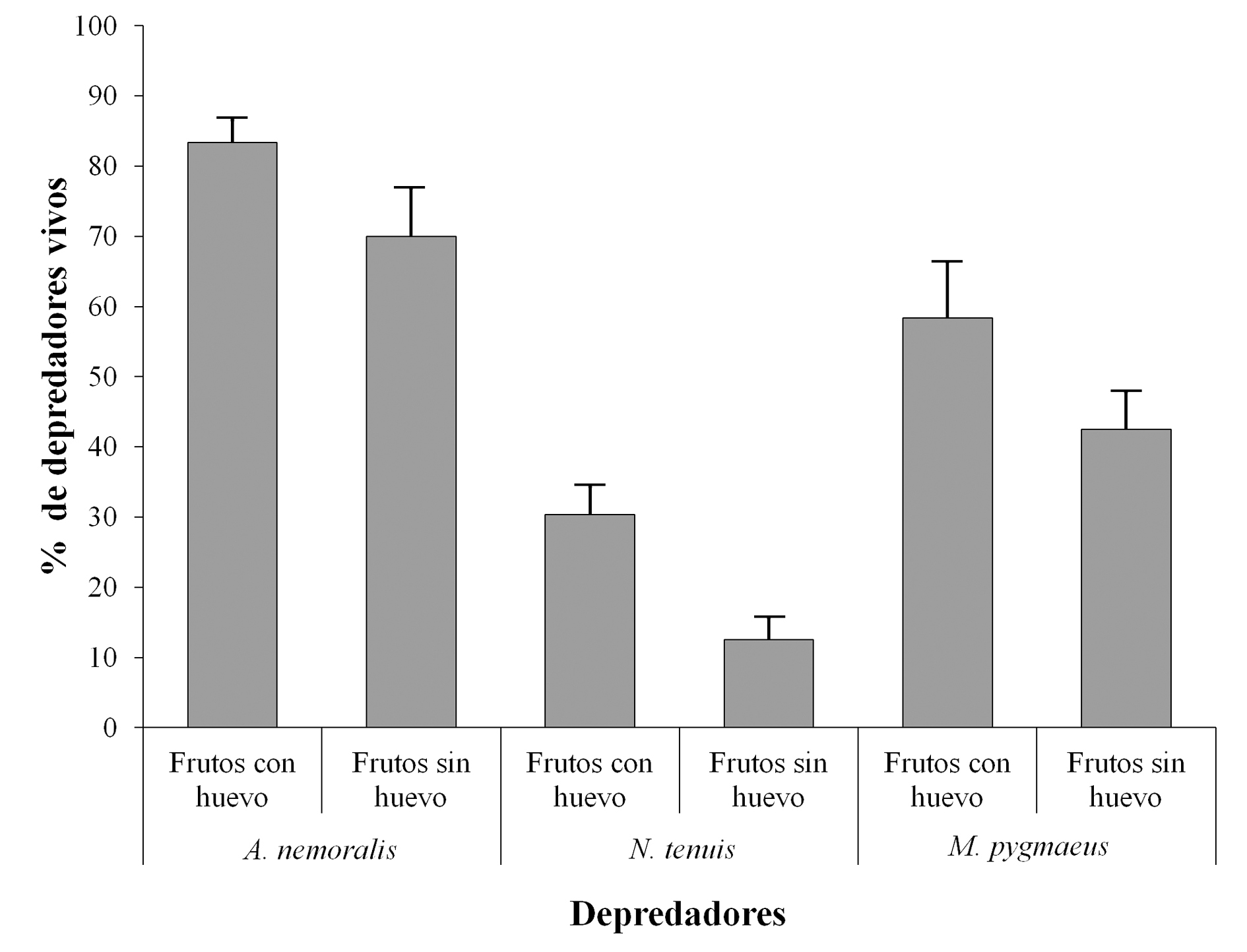
Figura 2 Porcentaje medio de depredadores vivos tras 72 h de exposición a frutos infestados (tratamiento) y a frutos no infestados (control). El símbolo * indica diferencias significativas (P < 0,05).
Previamente, se documentó la baja o nula actividad de O. laevigatus sobre frutos con puesta de D. suzukii en laboratorio (Malagnini et al. 2014), aunque según Gabarra et al. (2015) O. laevigatus sí sería capaz de alimentarse de huevos. Nuestros resultados confirmarían el bajo impacto de este tipo de depredadores sobre la puesta de D. suzukii. Sin embargo, estas especies podrían alimentarse de otros estados de desarrollo como pupas, adultos y larvas (Cuthbertson et al. 2014) por lo que no se puede descartar que la acción de estos hemípteros tenga algún tipo de impacto sobre las poblaciones de D. suzukii en el cultivo. No existen datos sobre la actividad de N. tenuis y M. pygmaeus sobre la puesta de D. suzukii. Estos dos depredadores están presentes y son utilizados en el control biológico de plagas importantes de invernaderos del sur de Europa con resultados satisfactorios (Desneux et al. 2010). Las dos especies pueden alimentarse de huevos de algunas especies plaga (e. g. lepidópteros) aunque muestran preferencia por larvas (Urbaneja et al. 2009). Los resultados indican que ninguno de los dos depredadores inciden de manera significativa sobre la puesta de D. suzukii. Ensayos previos demuestran que A. nemoralis puede depredar sobre adultos de D. suzukii, preferentemente sobre machos (Cuthbertson et al. 2014). Este antocórido es considerado generalmente un depredador específico de psílidos, aunque también se ha demostrado su potencial de control contra áfidos (Meyling et al. 2003). En las condiciones del presente estudio, esta especie produjo una reducción significativa de la emergencia de adultos de D. suzukii cuando está en contacto con frutos infestados, lo que indicaría cierto potencial de acción contra esta especie. Sin embargo, aunque los ensayos en laboratorio suponen un primer paso, son necesarios estudios de campo y bajo condiciones más reales para dilucidar el verdadero potencial de control de esta especie sobre D. suzukii. En este sentido, es necesario abordar aspectos tan importantes como si, en las condiciones de los cultivos afectados por D. suzukii (e.g. frutos rojos), este depredador sería capaz de incidir y buscar huevos en los frutos o cual sería el nivel de depredación sobre éstos en presencia de presas alternativas.
Conclusiones
De las cuatro especies de hemípteros incluidos en el estudio, sólo A. nemoralis produce una reducción significativa en la emergencia de adultos por fruto, sugiriendo que este antocórido podría producir un impacto sustancial en la puesta de D. suzukii. Sin embargo, son necesarios estudios bajo condiciones de campo que confirmen los resultados obtenidos en laboratorio y analizar el verdadero potencial de esta especie, así como las posibilidades de su inclusión en programas de manejo integrado.

