











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Raspberry weevil Aegorhinus superciliosus (Guérin- Méneville, 1830) (Coleoptera: Curculionidae) is an insect native to Chile and part of Argentina. Its north-to-south distribution in Chile extends from the Maule (35°25’36”S, 71°40’18”W) to the Los Lagos regions (41°28’18”S, 72°56’12”W) (Parra et al. 2009).
Exotic fruit trees in this part the country have recorded the highest land use growth over the last decades generating, an agro-ecosystem imbalance, favoring the colonization of A. superciliosus (Ellena et al. 2012). This insect pest has affected the whole shrub fruit group in south-central Chile and particularly European hazel (Corylus avellana L.) and blueberry (Vaccinium corymbosum L.) (Quintana et al. 2011; Zavala et al. 2011; Rebolledo et al. 2012).
Most of the damage produced by A. superciliosus occurs during its larval stage, as it feeds mainly on roots, causing dwarfism and weakening and during intensive attacks, larvae might possibly cause the death of the plant (Kuschel 1951; Elgueta 1993; Cisternas et al. 2000; France et al. 2000; Mutis et al. 2010; Medel et al. 2013; Tampe et al. 2016).
Maitén (Maytenus boaria Mol.) (Celastrales: Celastraceae) is a native tree from Chile and Argentina (Rodríguez et al. 1983), with important morphological variations along its wide distribution, extending in Chile from 29ºS to 49ºS latitude (from Atacama to the Magallanes regions). For this reason, and in relation to its wide environmental adaptability, this tree has been catalogued as a plastic species (Donoso 1993).
Characterized as a tree without forest used and despite being found in most forest types in Chile, forest-level research in maitén species has not generated interest (Donoso and Wendler 1985). Nonetheless, the medicinal use of its, foliage has prompt interest in chemical studies to prove the anti-inflammatory and antipyretic effect of their glycosidic components (Muñoz et al. 1995). Céspedes et al. (2000) studied the inhibitory effect of sesquiterpenes in M. boaria seeds in photosynthesis and in subsequent studies, the mechanisms associated with insecticidal effects of the same chemical compounds (Céspedes et al. 2001).
The large morphological variability found in A. superciliosus in its natural distribution has allowed us to consider these as subspecies (Aguilera and Rebolledo 2001). However, it is now estimated that the morphological differences are associated with the color and distribution of the exoskeleton scales (Elgueta 1993).
Taking into consideration the great genetic diversity found among insects species and between populations of the same species (Gadelhak and Enan 2005), the use of molecular tools has allowed for the generation of DNA “fingerprints”, used in the identification of individuals, species, subspecies, in genetic rank of populations and in green applications (Korpelainen et al. 2007).
The use of ISSR (Inter Simple Sequence Repeats) markers appears to be an efficient tool for the detection of genetic variations (Tikunov et al. 2003). This technique also allows for the examination of a large variety of genomic loci, using only one oligonucleotides sequence from 16 to 25 pb making the amplification of areas located between microsatellite DNA sequences, through PCR possible. Microsatellites are distributed through the genome of eukaryotic organisms and are flanked by highly conserved sequences (Chambers and MacAvoy 2000). The polymorphisms can be detected between individuals from the same population, mainly because this analysis is sensitive to the presence or absence of the genomic element recognized for the primer, and to length of the amplified intermediary sequence (Zietkiewicz et al. 1994).
A high DNA concentration is not required in the use of ISSR and no previous knowledge in the genome sequence of the studied individual is needed when designing the primers (Joshi et al. 2000), leaning on the high grade of polymorphism and huge distribution of microsatellites to detect low levels of differentiation, generating several polymorphic band (Yua et al. 2011).
Although A. superciliosus is considered as an important species, its biology background focuses mainly in behavior, growth and development (Aguilera and Rebolledo 2001). Studies associated with this curculionid in Chile provide important information regarding its geographic distribution (Kuschel 1951; Vergara et al. 2006), morphological description, characteristics of their different stages of development, life cycle, hosts (Parra et al. 2009), types of control (Mutis et al. 2009) and economic relevance (Aguilera 1994). Carrillo et al. (2002) addresses reproductive traits of oviposition and the taxonomic aspects are treated by (Elgueta 1993; 2000). Phylogenetic relationship, through its mitogenome sequence, allowed corroborating the monophyletic origin of A. superciliosus to the Aterpini tribe (Cabrera and Gaitán 2015).
In spite of the relevant contributions produced by the authors described previously, there is currently there is a lack of information associated with the degree of genetic variability in different populations along their geographic distribution in Chile. With the purpose of detecting possible similarities and differences at genetic level in A. superciliosus populations, this research analyzed the genetic variability among populations from southern Chile, associated with Maytenus boaria, through ISSR molecular markers.
Materials and methods
Collection of biological material
Adults individuals of Aegorhinus superciliosus were manually collected from barks of M. boaria in four different areas of Los Rios (39º48’30”S, 73º14’30”W) and Los Lagos (41º28’18”S, 72º56’12”W) regions, in the south of Chile. Calvertius tuberosus, was used as control group, due to its large genetic differences with A. superciliosus, adult individuals were collected in La Araucanía region (38º54’00”S, 72º42’00”W) (Table 1).
Table 1 Geographic location of the sampling areas.
| Region | City | Coordinates |
|---|---|---|
| Los Ríos | Máfil | 39°38’41,1”S, 72°57’08,7”W |
| Los Ríos | Futrono | 40°06’49,8”S, 72°27’18,6”W |
| Los Ríos | La Unión | 40°18’16,5”S, 73°04’20,1”W |
| Los Lagos | Frutillar | 41°07’21,5”S, 73°03’33,8”W |
| Araucanía* | Araucanía | 38°54’00”S, 72°42’00”W |
*Geographic location of the control group.
Homogenized tissue and total DNA extraction of A. superciliosus
The insects were stored at -80 °C in the Animal Biotechnology Research Laboratory (LINBA), at the Universidad de La Frontera Temuco, Chile, directly after their collection. Individuals were homogenized through disintegration in a previously ultraviolet irradiated (10 min) porcelain mortar and cooled at -80 °C. A total of 1-2 mg of homogenized tissue from each individual was obtained.
DNA extraction from homogenized tissue was carried out through the phenolic extraction protocol adapted from Sambrook and Green (2012), by addition of the previous homogenized individual mix into a 2 ml cell lysis tampon (Tris-HCl 25 mM pH: 8.4, EDTA 2 mM, SDS 1 %) supplemented with 20 µl of proteinase K (10 mg/ml). Incubation was performed in a thermoregulated bath at 55 °C for a minimum of 2 hours. Recovery of 0.7 ml of the lysed in sterile tube (1.5 ml) and application of 0.7 of phenol/chloroform/isoamylic alcohol 25:25:1 (pH: 8.4). Invert to mixed and centrifugation at 10.000 rpm for 10 minutes. Recovery of 0.5 ml from aqueous phase and transfer to a sterile tube. Addition of 0.3 ml of ammonium acetate (10 M) and 1 ml of cold pure ethanol. Incubation of the sample at -20 °C for 20 min. After the incubation period, DNA was extracted through a sterile glass rod to an eppendorf tube (1.5 ml), where it was washed with 1 ml ethanol (75 %) and centrifuged at 5.000 rpm for 1 min. The latter two steps were repeated three consecutive times. To finish, samples were oven drying at 40 °C for 10 min (open lid tube) and DNA sediments suspended with 100 µl of TE buffer.
Primers design to ISSR markers amplification
Based on the methodology used by Korpelainen et al. (2007), 17 ISSR primers were selected (Table 2) and seven were designed in the LINBA (AC-T, CA-G, GA-C, AG-C, AC-C, CA-A, CAG) and synthesized by the Integrated DNA Technologies (IDT), USA. These starters were used to amplify ISSR to PCR from A. superciliosus DNA.
Table 2 Primers used in ISSR analyses.
| Primer’s name | Sequence | Primer’s name | Sequence |
|---|---|---|---|
| AC-T | [AC]8T | CAA | [CAA]5 |
| CA-G | [CA]9G | CA-A | [CA]9A |
| GA-T | [GA]9T | CAG | [CAG]6 |
| GA-C | [GA]8C | ATG | [ATG]5 |
| GA-A | [GA]9A | GA-C | [GA]9C |
| AG-T | [AG]8T | AC-G | [AC]9G |
| AG-C | [AG]8C | CA-T | [CA]9T |
| AG-G | [AG]9G | GA-C | [GAC]5A |
| AC-C | [AC]8C |
Amplification conditions of A. superciliosus were adapted and modified according to the protocol described by Pérez de la Torre et al. (2012). Terminal amplification conditions were as follows: (a) denaturation for 1 min at 94 °C and for 30 sec at 98 °C (30 cycles); (b) annealing for 30 sec at 55 °C (30 cycles); (c) initial extension for 40 sec at 72 °C (30 cycles) and (d) final extension for 40 sec at 72 °C. The reaction mix was carried out as follows: SapphireAmp Fast PCR Master Mix (2X) 10 µl, DNA (200 ng) 2 µl, Primers ISSRs 2 µl (17 primers) and ultra pure H2O 6 µl (final volume of 20 µl). The amplification reaction was carried out using a MultiGene Gradient Thermocycler (Labnet International Inc.).
DNA amplification of A. superciliosus via PCR using ISSR
A total of 17 primers were evaluated and on the basis of their polymorphic expression patterns, five primers ([AC]8-C, [GA]8-C, [GA]9-A, [CAA]5 y [GAC]5A) were selected. The PCR reaction conditions were assessed as follows: SapphireAmp Fast PCR Master Mix (2X) 10 µl, DNA (200 ng) 2 µl, ISSR primers 2 µl for each and H2O ultra-pure 6 µl (final volume of 20 µl). The DNA amplification was carried out using a MultiGene Gradient Thermocycler (Labnet International Inc.) (Fig. 1).
Data analysis
Amplified bands, different and reproducible markers were classified using a value of “1” (presence) and “0” (absence). The data generated were arranged in a binary matrix used for the genetic variability analysis through the Nei’s model (Nei 1972) and the construction of a dendrogram by the UPGMA (Bootstrap de 1000) method. Furthermore, the analysis took into account the estimations of the Genetic Differentiation Coefficient (Gst); the Genetic Flow Estimation (Nm) from Gst (McDermott and McDonald 1993) and the Calculation of the Nei’s Genetic Diversity Index (h) (Nei 1973). Through POPGEN version 1.32 program (Yeh et al. 1999).
The Mantel test (Mantel 1967) with 9999 permutations was carried out using the Gene AlEx6 program. And then establishing the relation among the genetic distance and geographic distance, and estimate isolation patterns that could be exists in this population (Peakall and Smouse 2006).
Results
The primers selected generated 28 amplified bands 100 % polymorphic, ranging in size between 250 and 1,100 pb (Table 3).
Table 3 Primers used in ISSR amplification and number of generated bands for each primer.
| Primer | Sequency | Total number of bands | Polymorphic Bands (%) | Amplicon’s size (pb) |
|---|---|---|---|---|
| AC-C | [AC]8C | 6 | 100 | 500-150 |
| CAA | [CAA]5 | 6 | 100 | 1000-400 |
| GAC | [GAC]5A | 8 | 100 | 1000-300 |
| GA-A | [GA]9A | 5 | 100 | 1100-400 |
| GA-C | [GA]8C | 3 | 100 | 1100-250 |
| TOTAL | 28 | 100 |
Genetic variability analyses showed substantially high identity values between geographically neighboring populations; Máfil-Futrono (95 %), Futrono-La Unión (99 %), La Unión-Frutillar (97 %). The genetic distance values between populations were: Máfil-Futrono (0.05), Futrono-La Unión (0.002), La Unión-Frutillar (0.03), indicating that the studied populations present practically identical allele frequencies (Fig. 2B).
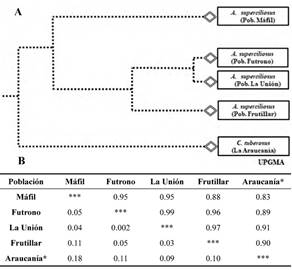
Figure 2 A. UPGMA dendrogram based on genetic distance of Nei (1978) between A. superciliosus populations. B. Genetic similarity (on the diagonal) and Genetic distances (under the diagonal) (Nei 1978) on studied populations. (Control: C. tuberosus, Araucania region).
The average genetic differentiation value between populations was low (Gst = 0.14). The estimated number of individuals migrating between populations in each generation reached an average value of 2.9 (Nm). Nei’s genetic diversity index (h) provided identical values to the populations of Futrono and La Unión (h = 0.11) and lower to those for to Máfil and Frutillar populations (0.07 and 0.08 respectively), which indicates a low genetic variability grade within the studied populations.
A recent cluster that groups Futrono and La Unión populations was possible to be observed through the dendrogram generated by the UPGMA method, collaterally presenting a common ancestor to the Frutillar population. Máfil population, however, is separated from the other three, which would indicate that it apparently derives from a more ancient clade (Fig. 2A).
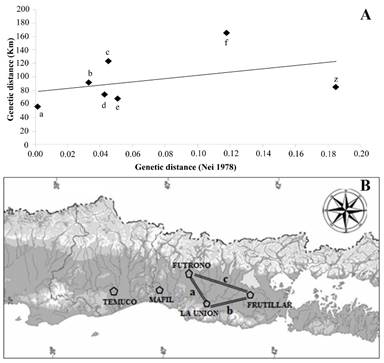
A direct relationship between genetic and geographic distance matrices (Fig. 1) was provided by the Mantel test. Therefore, the greater geographic distance between two populations, smaller their genetic distance. The Máfil population was more independent of the other three as supported by the UPGMA method results. The control group was the farthest when compared to all the studied populations (Table 4).
Table 4 Relationship between genetic and geographic distance (Mantel test).
| Points | Between- populations | Genetic distance (Nei 1978) | Geographic Distance (Km) |
|---|---|---|---|
| A | Futrono - La Unión* | 0.00 | 57 |
| B | La Unión - Frutillar* | 0.03 | 91 |
| C | Futrono -Frutillar* | 0.05 | 123 |
| D | Máfil - La Unión | 0.04 | 74 |
| E | Máfil -Futrono | 0.05 | 67 |
| F | Máfil - Frutillar | 0.12 | 165 |
| Z | Mafil - Temuco (control) | 0.18 | 86 |
Discussion
The high genetic identity values observed between the populations of Futrono, La Unión and Frutillar may partly be explained by both the geographic proximity between them and the presence of M. boaria, the main host across all the studied territory. The principle of isolation by distance (Wright 1943) the geographically neighbors populations are more similar at the genetic level than the populations that are farthest, phenomenon that is possible to observe within the populations in study.
Southern Chilean landscape (38°54’00”S, 72°42’00”W to 41°28’18”S, 72°56’12”W) is characterized by a continuous vegetation cover along wild and rural areas, paths and roads, where M. boaria is a frequent tree (Hoffmann 1998). Maytenus trees grow in association with Rubus ulmifolius (Schott), Salix viminalis (L.), Rubus idaeus (L.) species, all of which are cataloged as usual hosts of A. superciliosus (Zavala et al. 2011). Therefore, all this vegetation plays an important role in the distribution and dispersal of this pest.
This vegetation cover in the studied area, allowing the genetic flow, thus generating lesser variability among the studied populations, as supported by the low genetic differentiation value of the populations (Gst = 0.14).
The genetic flow estimator Nm provided an average value of 2.9, indicating little genetic differentiation between the studied populations, as also observed by McDermott and McDonald (1993), where the average migrant individuals is sufficient to prevent a substantial differentiation between these populations. Slatkin (1985) proposed that the low genetic distance values are mainly due to the genetic flow existence, which was clearly observed in the Aegorhinus populations under study. Therefore, the isolation by distance phenomenon proposed by Wright (1943), in which individuals tend to associate with the ones who are geographically closest, generate low genetic variability in their populations.
Mantel test is centered in producing matrices of genetic similarity and geographic distance derived from original data. It is interpretation makes the genetic similarity of neighboring populations clear (López and Olano 2006). The positive correlation found between geographic distribution and genetic variability (Fig. 1A) of neighboring populations (Futrono-La Unión-Frutillar) supports the understanding of the genetic flow and the constant presence of Maytenus along the studied territory.
Additionally, Mantel test results not only showed that neighboring populations had a high genetic similarity, but also that geographically distant populations displayed a greater genetic distance between them. This was the case of the Máfil population (the northernmost) in comparison to the populations of Futrono, La Unión and Frutillar (Table 4). Therefore, the positive correlation observed is solid proof to discard chance (Castellano and Balletto 2002).
The low genetic distance values found in A. superciliosus populations in this study (Fig. 2B), are supported by the results obtained by Elgueta (1993). The author describes that the morphological differences observed are correlated with the color and the scales distribution on the exoskeleton. These characteristics may be possibly associated with the age of the adult individuals or with the effect of the high phenotypic plasticity exhibited by the coleopterans, which allows them to adapt to many and different environmental conditions (Moczek 2010).
The genetic distance between the Máfil population (the northenmost) and the Frutillar population (the southernmost) is the greatest compared with the other populations. Two possible explanations have been provided: (a) the distance between these populations (165 km) is the longest among the four under study, (b) the different weather conditions associated with these populations. The Máfil population has the lowest average rainfall in the Los Rios region (1,200 to 1,600 mm per year), in contrast to the Frutillar population (in the Los Lagos region), with an annual average up to 3,000 mm (INE 2015).
Conclusions
The low levels of genetic variability among A. superciliosus populations in southern Chile, is probably to be associated to the large number of vegetal hosts found in agricultural and forest ecosystems. These conditions contribute to increase the genetic flux between their populations and they allow that A. superciliosus to be considered one of the most important and most harmful blueberry pests in Chile.

