











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
Introducción
El género Monalonion (Herrich-Schaeffer, 1853) (Hemiptera: Miridae) de la subfamilia Bryocorinae (tribu Monaloniini) presenta una distribución Neotropical (Namyatova et al. 2016). En Centro y Sur América, algunas especies de Monalonion se han reportado atacando cultivos de cacao Theobroma cacao (L., 1753) (Malvaceae), aguacate Persea americana (Mill., 1768) (Lauraceae) y guayaba Psidium guajava (L., 1753) (Myrtaceae) (Wheeler 2000). Monalonion velezangeli (Carvalho y Costa, 1988) es una plaga polífaga que en Colombia se ha registrado en 19 especies de plantas ornamentales, arvenses, maderables y frutales entre las que se encuentra el café, Coffea arabica L. (Rubicaeae) (Ramírez-Cortés et al. 2008; Giraldo y Benavides 2012; Rodas et al. 2014; Ocampo et al. 2018).
En el cultivo del café, M. velezangeli se conoce como “la chinche de la chamusquina del café”. En Colombia, este insecto se encontró por primera vez atacando este cultivo en 1998, en los municipios de La Plata, La Argentina, Paicol y El Pital del departamento del Huila. Desde entonces, su presencia se ha registrado en las regiones cafeteras de los departamentos de Cauca, Nariño y Valle del Cauca (Benavides et al. 2013). El insecto ataca los brotes nuevos de las plantas y necrosa los tejidos cuando se alimenta, ya que inyecta enzimas hidrolasas tipo poligalacturonasa que licuan el contenido celular del mesófilo para posteriormente succionarlo. Este proceso de digestión extra oral ocasiona el necrosamiento del tejido y la formación de pústulas de color marrón sobre los brotes (Wheeler 2001). El ataque induce a la planta a una producción continua de follaje y escasa formación de fruto y como consecuencia toma una apariencia de achaparramiento con reducción de las floraciones. Las plantas afectadas por este insecto, presentan retrasos en su crecimiento, reducción en la productividad debido al desbalance que sufren como respuesta al daño (Ramírez-Cortés et al. 2008).
Monalonion velezangeli usualmente se encuentra en cafetales localizados en altitudes entre 1.650 y 2.100 msnm, en zonas con baja luminosidad (inferior a 1.400 horas/año) y con promedios de temperatura inferior a 20 °C y humedad relativa superior a 80 % (Ramírez-Cortés et al. 2008; Giraldo et al. 2010). En el departamento del Huila, el 81 % del café se encuentra sembrado entre los 1.400 y los 2.000 msnm, rango altitudinal que por sus condiciones climáticas resulta ideal para el desarrollo de la plaga (Benavides et al. 2013); es por ello que actualmente se reportan cerca de 8.000 ha de café afectadas por el insecto .
En la actualidad, la recomendación técnica para el control de M. velezangeli en el cultivo del café, comprende manejo del cultivo y prácticas de control cultural, si es necesario la aplicación de insecticidas químicos de forma localizada. Sin embargo, en campo se observa que los caficultores realizan aplicaciones generalizadas de insecticidas de amplio espectro, práctica que afecta la fauna benéfica del cafetal (Benavides et al. 2013).
En estudios realizados en Cenicafé, se han observado en campo algunos enemigos naturales que depredan ninfas y adultos de M. velezangeli en particular se reportan arañas e insectos de la familia Reduviidae (Hemiptera) (Giraldo et al. 2011; Laiton et al. 2018). Además, se han encontrado hongos entomopatógenos (Ramírez-Cortés et al. 2008) como Beauveria bassiana (Balsamo) (Vuillemin, 1912) (Hypocreales: Cordycipitaceae) causando mortalidad de M. velezangeli en aguacate (Londoño 2014) y cacao (Pinzón et al. 2008). En ambos cultivos se recomienda su uso para el control del insecto. Cenicafé (2008) reportó un aislamiento del hongo entomopatógeno Clonostachys sp. (Hypocreales: Bionectriaceae), a partir de un adulto de M. velezangeli, cepa que posteriormente mostró ser patogénica sobre ninfas y adultos de la plaga. En Argentina Toledo et al. (2006), encontraron a este hongo como micoparásito de hongos fitopatógenos de importancia económica y como entomopatógeno del cicadélido Oncometopia tucumana (Schröder, 1959). Sin embargo, las pruebas realizadas en Cenicafé mostraron una baja esporulación de esta cepa, lo cual podría dificultar su uso en un programa de control biológico en aplicación por aumentación en campo (Goettel et al. 2005). Además, en el cepario de Cenicafé existen al menos dos aislamientos del hongo B. bassiana provenientes de Monalonion sp., en donde una de las cepas presentó alta virulencia sobre Monalonion dissimulatum en condiciones de laboratorio (datos no publicados) lo que evidencia que dicha cepa podría ser efectiva contra Monalonion velezangeli.
Actualmente se carece de información acerca del efecto de hongos entomopatógenos en el control de M. velezangeli en café en condiciones de campo. Sin embargo, los resultados reportados a partir de pruebas en laboratorio indican que su uso podría ser promisorio (Cenicafé 2018).
El propósito de este trabajo fue evaluar el efecto de una cepa del hongo entomopatógeno B. bassiana sobre el control de M. velezangeli bajo condiciones de laboratorio y campo en el departamento del Huila, en plantaciones de café con la finalidad de encontrar una cepa virulenta que pueda ser utilizada para el control biológico del insecto incorporándolo en una estrategia de manejo integrado de la plaga.
Materiales y métodos
I. Patogenicidad y virulencia de Beauveria bassiana cepa B sobre Monalonion velezangeli en laboratorio
La cría de M. velezangeli se realizó sobre plantas de Cissus verticillata (L.) (Vitaceae), siguiendo el método reportadó por Cenicafé (2017a). La cría se hizo con el fin de permitir la sobrevivencia del insecto en todos los estados biológicos y mantener poblaciones a través del tiempo. Ésta se desarrolló en los laboratorios de la disciplina de Entomología de Cenicafé, bajo condiciones controladas de temperatura 20 ± 1 °C, humedad relativa entre 80 % y 90 % y fotoperiodo de 12 horas luz. Las muestras de M. velezangeli para el pie de cría, se recolectaron sobre café en la finca El Divino Niño, Vereda El Billar, Ansermanuevo (Valle del Cauca 4º48̓57.1”N 76º05̓40.2”O) a una altitud de 1.849 msnm. Cada pareja (1♂: 1♀) se dispuso en cajas de acrílico de 32 cm de largo, 20 cm de ancho y 11 cm de alto con dos orificios de ventilación en la tapa, forrados con tela muselina blanca y en su interior se depositaron tres ramas frescas (con tallos lignificados) de Cissus vertillata. En la base de cada caja se colocó una servilleta la cual se humedeció diariamente con agua destilada.
Para la eclosión de los huevos, los tallos de C. vertillata que contenían las posturas de M. velezangeli, se sembraron en discos turgentes de turba canadiense (“jiffys”); por cada “jiffy” se sembraron entre dos y cuatro tallos y cada cinco días las turbas se hidrataron con 50 ml de solución nutritiva de Hoagland (laboratorio PhytoTechnology), en una concentración de 1,63 g/L de agua. Cuando emergieron las ninfas, se dispusieron en cajas de acrílico transparente de 21 cm de largo x 11 cm de ancho y 8 cm de alto, con dos orificios de ventilación en la tapa, forrados con tela muselina blanca, una servilleta húmeda en la base y hojas frescas de C. vertillata (entre 5 y 7 dependiendo el tamaño), en cada caja se colocaron cinco ninfas. Para disminuir los factores de mortalidad, el alimento y las cajas se cambiaron cada dos días.
En el cepario de Cenicafé existían dos aislamientos del hongo Beauveria bassiana aislados en 1990 de Monalonion sp., donde una de las cepas nombrada como cepa B había mostrado alta virulencia sobre Monalonion dissimulatum en laboratorio (datos no publicados) por lo que fue escogida para ser evaluada sobre M. velezangeli. Para evaluar la virulencia de la cepa, ésta fue reactivada del cerapio de Cenicafé y se cultivó en cajas de Petri con medio Agar Papa Dextrosa (PDA) (Merck), se incubó por 15 días bajo condiciones controladas de temperatura 25 ± 2 °C, humedad relativa de 71 ± 5 %, con un fotoperíodo de 12:12.
Antes de evaluar la patogenicidad del hongo, se determinó que el porcentaje de esporulación de las conidias fuera superior a 90 %. Se preparó una solución de la cepa a una concentración de 1 x 107 conidias/ml. Se tomaron 50 ninfas de M. velezangeli que fueron colocadas en una pieza de tela de tul (15 x 15 cm) y se sumergieron por espacio de 30 segundos en 100 ml de la solución de conidias contenidos en un vaso de precipitado de 150 ml. Como testigos se sumergieron otros 50 insectos en agua. Las ninfas tratadas fueron transferidas a las unidades experimentales, que consistieron en una caja de acrílico rectangular de 7,5 cm de alto x 21,5 cm de largo y 10 cm de ancho, la cual contenía ramas con hojas de la planta C. verticilllata y una servilleta húmeda en la base. En cada caja se colocaron cinco individuos de M. velezangeli, en total nueve cajas por tratamiento. Las cajas se mantuvieron a 23 °C y se revisaron diariamente durante ocho días, contabilizando el número de individuos muertos y el número de ninfas que pasaron al estado adulto. Los insectos muertos se sacaron de las cajas y se incubaron en cámara húmeda con el fin de registrar signos de infección causados por el hongo. Los días de evaluación estuvieron determinados por la duración del material vegetal del tratamiento testigo.
La variable de respuesta fue el porcentaje de mortalidad de los insectos al cabo de ocho días, se realizó un análisis de varianza para determinar diferencias en la variable mortalidad en el testigo vs hongo y se determinó la presencia de micelio y conidias del hongo en los insectos.
II. Evaluación de Beauveria bassiana cepa B sobre Monalonion velenzageli en jaulas bajo condiciones controladas similares a invernadero
Para evaluar el efecto del hongo en condiciones controladas, se sembraron estacas de plantas de C. verticillata en materos y luego de cuatro meses de su establecimiento, se infestaron con ninfas y adultos de M. velezangeli. Los individuos de M. velezangeli se recolectaron en campo sobre plantas de café en fincas de la vereda El Billar en Ansermanuevo. Cada planta se introdujo en una jaula entomológica de 50 cm x 50 cm x 50 cm cubierta de tul y se infestó con seis insectos (ninfas y adultos), conformando así la unidad experimental (Fig. 1). Luego, las plantas infestadas dentro de la jaula se asperjaron con dos tratamientos. El tratamiento 1, consistió en la aplicación de la solución de B. bassiana a una concentración de 4 x 1010 conidias /L, aplicando 20 cc por planta. El tratamiento 2, testigo consistió en la aplicación de agua. Se tuvieron 10 repeticiones por tratamiento.

Figura 1 Planta de Cissus verticillata dispuesta en la jaula entomológica para la aplicación de los tratamientos.
Las unidades experimentales, luego de la aplicación fúngica se mantuvieron bajo condiciones controladas (Fig. 2) a una temperatura de 20 °C, 80-90 % de humedad relativa y un fotoperiodo de 12 h.

Diariamente durante cinco días, en cada una de las plantas se cuantificó el número de insectos vivos y muertos. Los insectos muertos se sacaron de las jaulas y se incubaron en cámara húmeda con el fin de registrar signos de infección causados por el hongo. La variable de respuesta fue el porcentaje de mortalidad. Con los promedios de mortalidad se realizó un análisis de varianza de una vía para determinar diferencias estadísticas entre los dos tratamientos.
III. Evaluación de la Beauveria bassiana cepa B para el control de Monalonion velezangeli en condiciones de campo
Este experimento se realizó en mayo de 2018, en la vereda Las Nieves, en el municipio de Tarqui, Huila, a una altitud de 1.695 msnm (2º07̓03.1”N 75º54̓16.9”O). Se seleccionaron lotes de café variedad Castillo® de 18 meses de edad afectados por la plaga (Fig. 3). Se realizaron dos ensayos para evaluar el efecto de la aplicación de los siguientes tratamientos: T1: hongo B. bassiana cepa B a una concentración de 4 x 1010 conidias/L, T2: insecticida Clorpirifos a una concentración de 4 cc/L y T3: agua, en el tratamiento testigo del insecto.

El hongo se produjo en el laboratorio de Cenicafé usando como sustrato arroz en bolsas de 30 x 20 cm de polipropileno con 60 g de arroz previamente esterilizado. Las bolsas de arroz se humedecieron con 20 ml de agua estéril y se inocularon con 5 ml de una solución de 1 x 107 conidias/ml. Estas bolsas se dejaron en incubación a 25 °C y fotoperiodo regulado de 10 horas luz/día, durante 15 días.
Se produjeron 2,4 kg de arroz con conidias a una concentración de 4,4 x 109 conidias/g. Se calculó la cantidad de hongo en arroz requerida para asperjar 500 árboles a una concentración de 4 x 1010 conidias/L. Se prepararon 35 L de solución con hongo para esto se tomaron 322 g del arroz con conidias, se disolvieron en 5 L de agua y luego se aforó para completar los 35 L. Se asperjaron 70 cc de cada tratamiento por árbol, con el fin de lograr un cubrimiento completo de la planta que alcanzara tanto haz como envés de las hojas, pero con énfasis en el tercio medio superior. Las aplicaciones se realizaron después de las 16:00 horas.
Se realizaron dos ensayos de campo. El primero se hizo con el fin de evaluar el método de aspersión de los tratamientos y observar si las unidades experimentales se verían afectadas por poblaciones vecinas de la plaga que llegaran a atacar las plantas después de las aspersiones. Posteriormente, se realizó un segundo ensayo con el cual se pretendía evaluar a través del tiempo, durante dos meses el efecto del hongo sobre la plaga en un lote de café afectado.
Ensayo 1:
Para este ensayo se escogieron grupos de nueve árboles: un árbol central infestado con una o dos ninfas de M. velezangeli (unidad efectiva) y ocho árboles alrededor, el conjunto conformó la unidad experimental. Los tratamientos se asperjaron sobre todos los árboles que componían la unidad experimental. Se contó con 21, 21 y 17 repeticiones para los tratamientos T1, T2 y T3, respectivamente, distribuidos de forma aleatoria dentro del lote. Se realizaron evaluaciones a los ocho y 15 días después de aplicados los tratamientos, tomando como variable de respuesta la presencia de daño fresco ocasionado por M. velezangeli en las unidades efectivas, es decir, la presencia de lesiones recientes en los brotes del árbol, ocasionadas por la alimentación de un individuo durante el mismo día.
Ensayo 2:
Este ensayo se realizó en un lote con 2.000 árboles a una distancia de siembra de 1,5 x 2 m, que mostraba una distribución homogénea de la plaga. El lote se dividió en tres parcelas separadas por 10 surcos. Cada parcela consistió de 450, 462 y 462 árboles, donde se aplicaron los tratamientos T1, T2 y T3 respectivamente. Se realizaron evaluaciones en forma de censo cada ocho días durante dos meses, sobre todos los árboles que componían las parcelas, tomando como variable de respuesta el número de brotes con daños frescos por árbol. Este dato se promedió para cada fecha de evaluación en cada tratamiento. Con estos datos se determinó el promedio del número total de árboles afectados por lote en cada fecha de evaluación y se calculó el porcentaje de infestación total en los lotes de cada tratamiento: Número de árboles con afectación nueva /Número total de árboles.
Resultados y discusión
I. Patogenicidad y virulencia de Beauveria bassiana cepa B sobre Monalonion velezangeli en condiciones de laboratorio
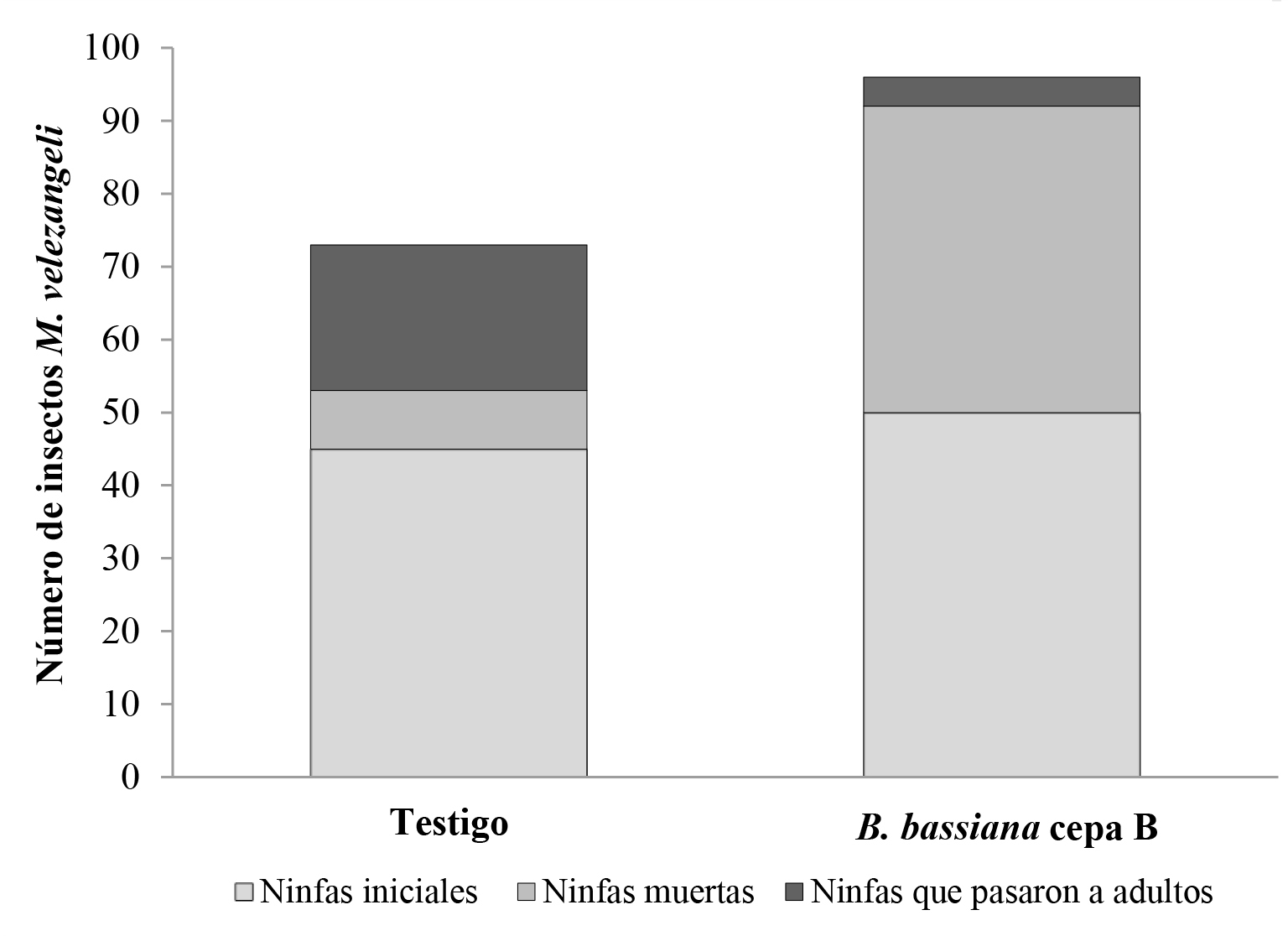
La cepa B mostró ser patogénica y altamente virulenta sobre M. velezangeli. Los resultados de mortalidad se observan en la Tabla 1, y la Figura 4 muestra los signos de infección del hongo que se observaron tanto en ninfas como en insectos adultos y en ninfas que alcanzaron la etapa adulta, pero murieron por efecto del hongo.
Tabla 1 Porcentaje de mortalidad causada por el hongo Beauveria bassiana sobre la población de Monalonion velezangeli.
| Tratamiento | No. ninfas iniciales | No. ninfas muertas | No. ninfas que pasaron a adultos | Porcentaje mortalidad |
|---|---|---|---|---|
| Testigo Agua | 45 | 8 | 20 | 18 |
| Cepa B | 50 | 42 | 4 | 84 |

Figura 4 Individuos de Monalonion velezangeli infectados con Beauveria bassiana cepa B. A. Ninfa. B. Adulto.
La Figura 5 muestra el desarrollo de la población del insecto durante cinco días sometido al tratamiento con el hongo al ser comparado con el testigo. Cabe aclarar que el bioensayo se programó para ocho días, pero se concluyó a los cinco días por la alta mortalidad ocasionada por el hongo durante ese período. El 84 % de la población de M. velezangeli sometida a B. bassiana murió y solo el 8 % pasó al estado adulto, cuatro insectos no fueron encontrados. El ANOVA (F = 43; df = 18; P < 0.01), mostró diferencias estadísticas entre el tratamiento y el testigo. Esto indica que el hongo, además de afectar el desarrollo biológico de M. velezangeli, mostró una alta virulencia sobre la plaga. Por el contrario, en el tratamiento testigo, el 45 % de la población pasó a adulto al cabo de los cinco días.
II. Evaluación de Beauveria bassiana cepa B sobre Monalonion velenzageli en jaulas bajo condiciones controladas similares a invernadero
En la Figura 6 se observan los resultados de mortalidad luego de 8 días de realizadas las aspersiones del hongo sobre los insectos mantenidos en C. verticillata. El ANOVA (F = 148; df = 19; P < 0.05), mostró diferencias estadísticas entre el tratamiento y el testigo.
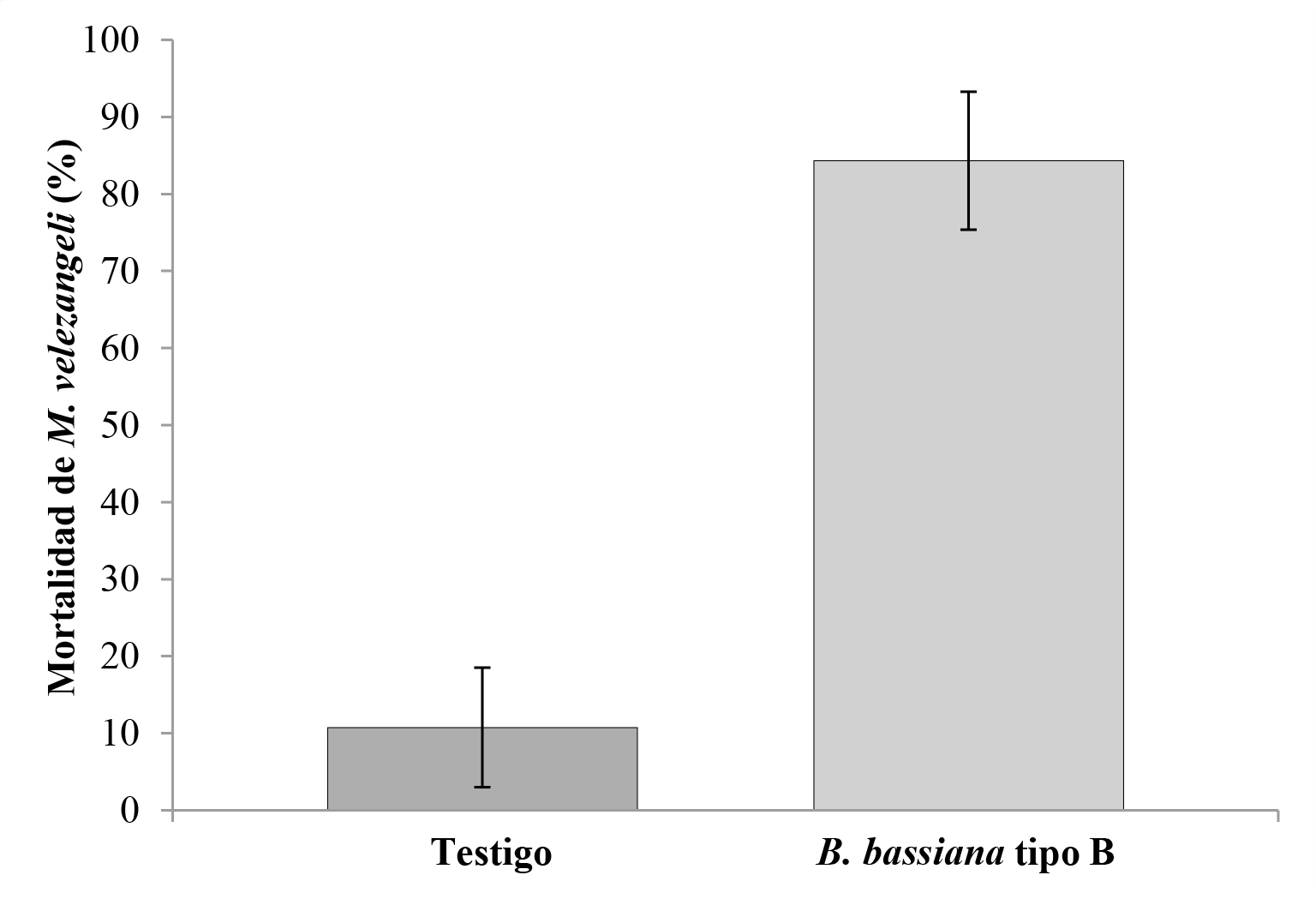
Figura 6 Porcentaje de mortalidad ocasionado por la cepa B de Beauveria bassiana sobre Monalonion velezangeli por tratamiento, en jaulas bajo condiciones controladas. Las barras representan el intervalo de confianza del 95 %.
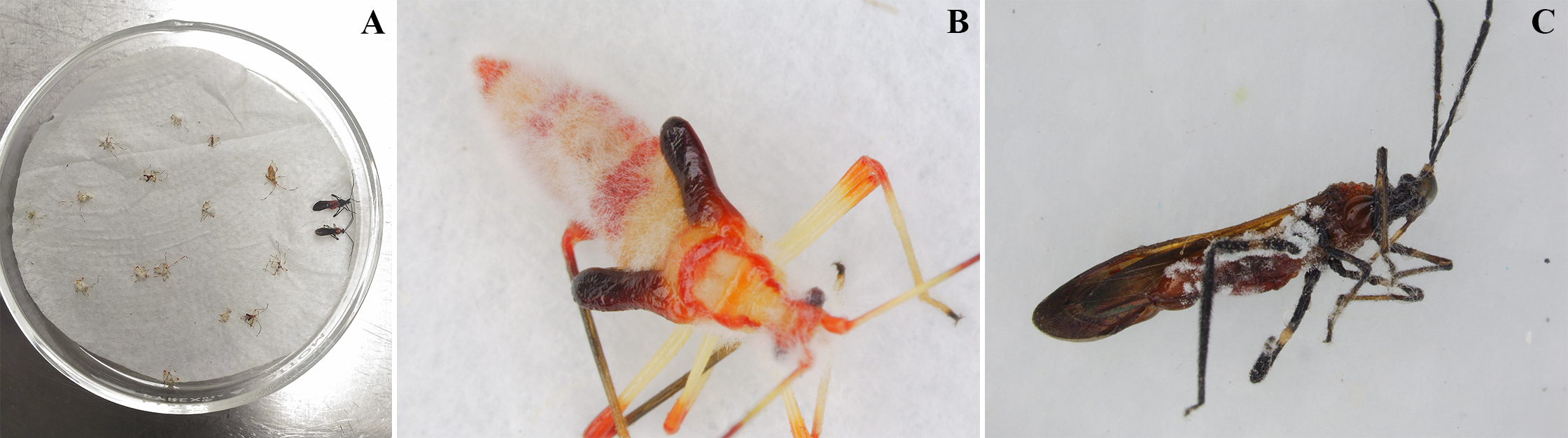
En el tratamiento testigo, la mortalidad natural de la población de insectos fue de 10 % en tanto que, en el tratamiento con aplicación del hongo, se obtuvo el 84 %. Los insectos muertos que fueron colocados en cámara húmeda (Fig. 7A ) mostraron signos de infección por B. bassiana. Se observó crecimiento del hongo en ninfas y adultos del insecto (Figs. 7B y 7C). Esto demuestra que todos los estados del insecto fueron susceptibles al hongo.
III. Evaluación de la Beauveria bassiana cepa B para el control de Monalonion velezangeli en condiciones de campo.
Ensayo 1:
Los resultados de campo de este ensayo se muestran en la Figura 8. Tanto el insecticida como el hongo causaron mortalidad sobre M. velezangeli al disminuir la afectación final en los árboles entre 76 y 86 %, respectivamente. Con estos resultados se continuó con el ensayo 2.
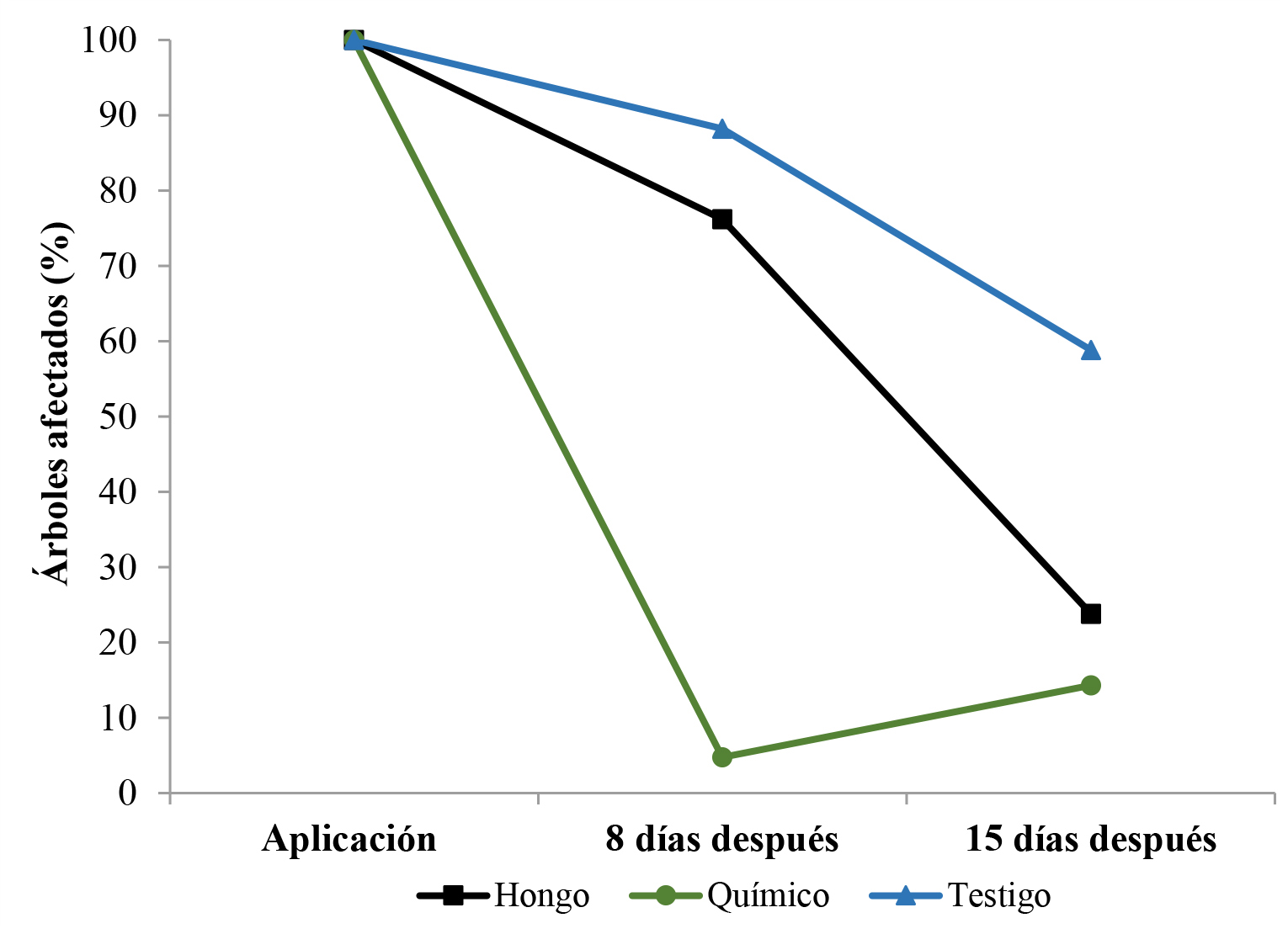
Figura 8 Porcentaje de árboles de café afectados por Monalonion velezangeli en cada evaluación durante 15 días. El tratamiento hongo (T1) corresponde a las aspersiones con Beauveria bassiana cepa B, el tratamiento químico (T2) a las aspersiones con Clorpirifos y el Testigo (T3) a las aspersiones con agua.
Ensayo 2:
El promedio del total de árboles de café afectados por M. velezangeli a través del tiempo, después de la aplicación de los diferentes tratamientos, se muestra en la Figura 9.
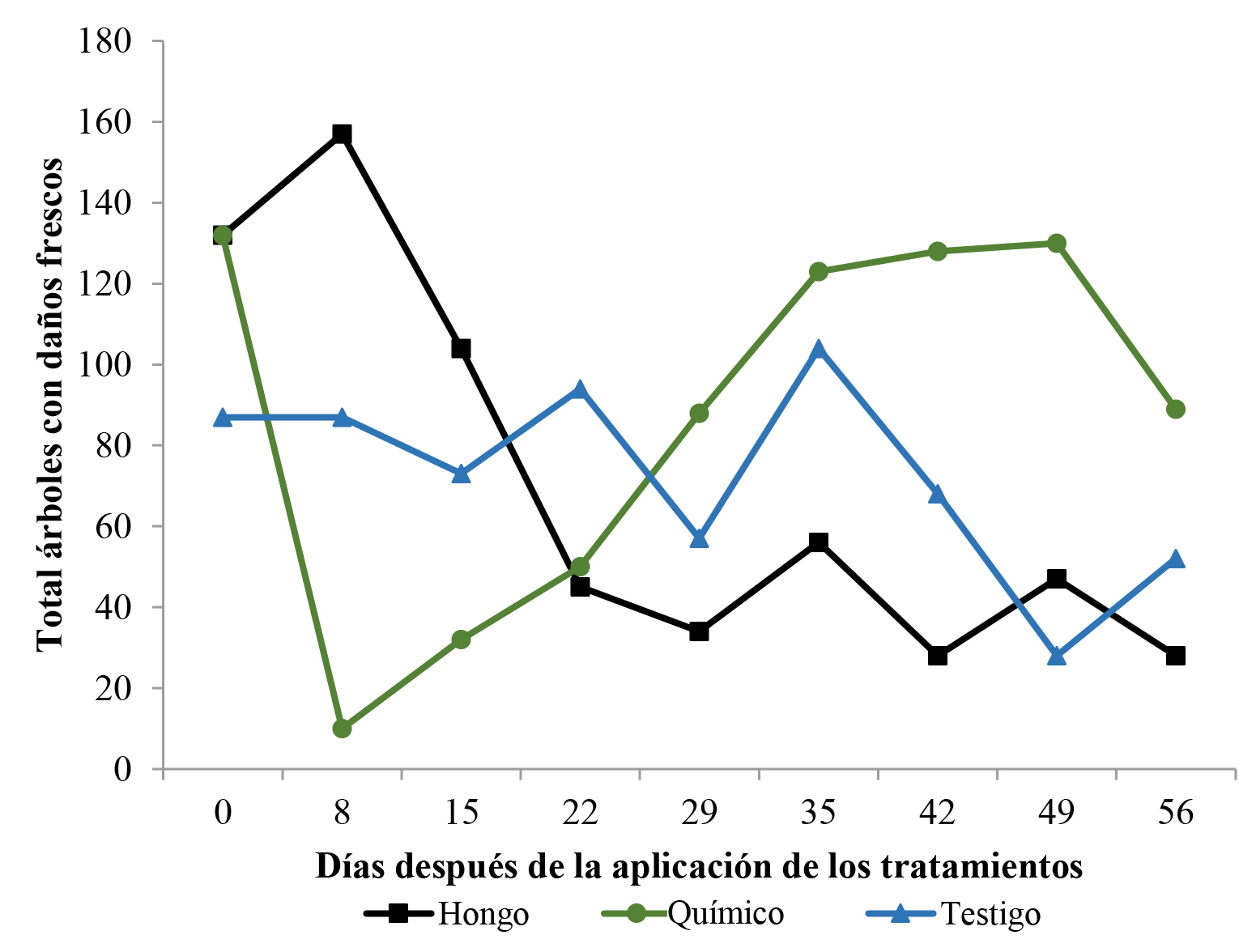
Figura 9 Número total de árboles afectados por Monalonion velezangeli a través del tiempo, después de la aplicación de los tratamientos. El tratamiento hongo (T1) corresponde a las aspersiones con Beauveria bassiana cepa B, el tratamiento químico (T2) a las aspersiones con Clorpirifos y el Testigo (T3) a las aspersiones con agua.
El número de árboles afectados inicialmente difirió en los tres tratamientos, siendo menor en el testigo; sin embargo, la aplicación del hongo logró disminuir significativamente las poblaciones de los insectos a partir de los 15 días, alcanzando un número menor de árboles afectados que en el testigo desde los 22 días (Fig. 9). Los porcentajes de infestación en las plantas de café en los tres lotes se observan en la Figura 10, el porcentaje de árboles afectados con los que se inició el ensayo difirió en los tres tratamientos, siendo menor en el testigo, en donde el porcentaje de infestación inicial fue del 19 % y al final del experimento fue del 11 %. Para el caso del insecticida químico, el lote inició con infestación del 29 % y al final del experimento la infestación estuvo cercana al 20 %. Este tratamiento mostró que la aplicación causó una disminución significativa de la población del insecto llegando a niveles de 2,16 % de infestación; sin embargo, tras la aplicación, la población de insectos se recuperó prontamente e incrementó a niveles casi iguales a los iniciales en aproximadamente un mes, de tal forma, que al final del experimento el mayor daño fue el observado en el lote tratado con insecticida químico. Este fenómeno es llamado resurgencia de plagas, y hace referencia al incremento repentino de las poblaciones del insecto plaga, posterior a una aplicación de insecticida de síntesis química (Harris 2000).
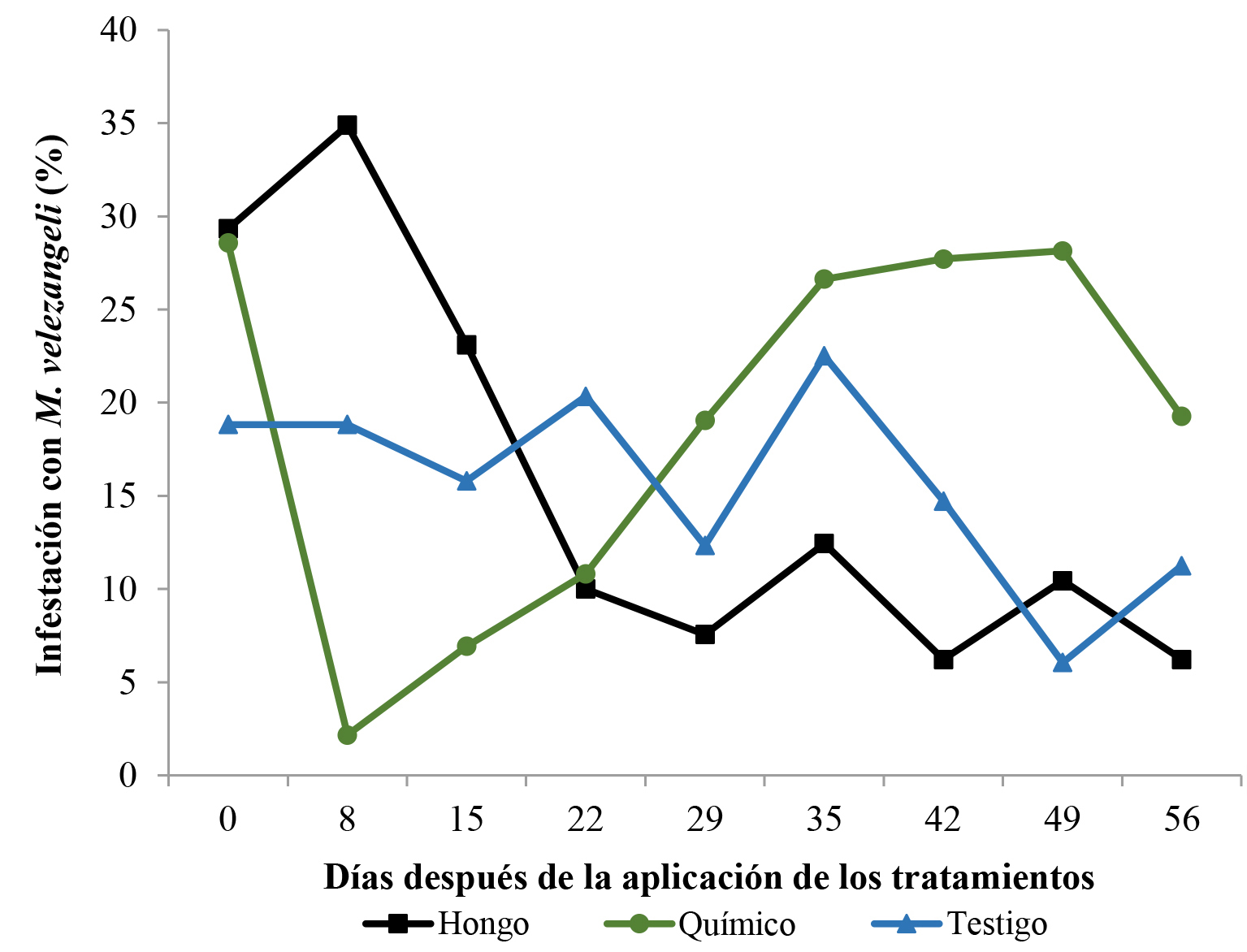
Figura 10 Porcentajes de infestación causado por Monalonion velezangeli en árboles de café durante el tiempo de observación del lote donde se aplicaron los tratamientos: hongo (T1) corresponde a las aspersiones con Beauveria bassiana cepa B, químico (T2) corresponde a las aspersiones con Clorpirifos y el Testigo (T3) a las aspersiones con agua.
La aplicación del insecticida químico en este ensayo al parecer genera un desbalance en el agroecosistema, debido a que, a pesar de que este tratamiento logró una disminución significativa de las poblaciones del insecto, por ser un insecticida de amplio espectro, también pudo haber afectado las poblaciones naturales de posibles biocontroladores de la plaga presentes en el lote, ya que, luego de ocho días de aplicación del insecticida, la población de la plaga incrementó rápidamente a niveles superiores a los observados en el lote testigo, sin ningún agente de control natural que lo impidiera. Este tipo de situaciones han sucedido en campos de alfalfa en Estados Unidos, donde aplicaciones constantes de insecticidas químicos para el control del áfido Therioaphis trifoli (Monell, 1882) (Hemiptera: Aphididae) eliminaron al depredador Hippodamia spp. (Coleoptera: Coccinellidae), lo que generó que la plaga se presentara a niveles mayores a los iniciales (Stern et al. 1959; Nicholls 2008).
Cenicafé (2017b), reportó seis especies de Reduviidae depredadores cohabitando con M. velezangeli dentro de cafetales afectados por la plaga en varios municipios del departamento del Huila. Entre esas especies se encuentran los depredadores Zelus vespiformis (Hart, 1987) y Arilus gallus (Stål, 1872), los cuales fueron observados durante el montaje de estos experimentos (Fig. 11). Estos depredadores pueden estar ejerciendo el control de las poblaciones de la plaga dentro de los cafetales y su posible supresión mediante la aplicación del insecticida químico de amplio espectro, pudo haber creado el desbalance en el agroecosistema de la parcela donde se aplicó dicho tratamiento.

Figura 11 Especies de Reduviidae depredadores de la chamusquina del café observados durante el experimento en campo. A. Zelus vespiformis. B. Arilus gallus.
En el caso del tratamiento con B. bassiana, la infestación inicial del lote fue de 30 %, la cual se incrementó hasta 35 % ocho días después de la aplicación; sin embargo, debido a su acción, luego de 15 días se observó un decrecimiento de la población hasta llegar al final del experimento con una infestación cercana al 5 % (Fig. 10). Este lote mostró el menor nivel de infestación de M. velezangeli.
En este tratamiento, la disminución de la población y muerte de M. velezangeli tardó más tiempo en comparación con el insecticida; al menos ocho días más, lo que indica que la mortalidad no es tan rápida como la observada con el insecticida químico. Sin embargo, una posible explicación de este suceso bajo las condiciones evaluadas es que el hongo actuó de manera específica al afectar solo la plaga y provocar un menor efecto en los depredadores, lo que, en conjunto, ejerció un control efectivo y constante sobre M. velezangeli, manteniendo una baja población durante el periodo de observación. Varios autores han reportado el efecto del uso de métodos de control selectivos vs. métodos de amplio espectro. Cuando Whalen et al. (2016) usaron insecticidas selectivos para el control de plagas de lepidópteros en soya, observaron que éstos no afectaron la población de depredadores de la plaga. Sin embargo, cuando usaron insecticidas de amplio espectro, el efecto tanto sobre la plaga como sobre los depredadores fue significativo.
En este estudio, el entomopatógeno se comportó como un agente de control selectivo, que no afectó a los depredadores, lo que mantuvo una reducción de la plaga hasta dos meses después de su aplicación. Estudios anteriores han reportado que el entomopatógeno B. bassiana no afecta la sobrevivencia de enemigos naturales como los depredadores Coccinella septempunctata (Linnaeus, 1758) (Coleoptera: Coccinellidae), Chrysoperla carnea (Stephens, 1836) (Neuroptera: Chrysopidae), Dicyphus tamaninii (Wagner, 1951) (Hemiptera: Miridae) (Thungrabeab y Tongma 2007), ni Harmonia axyridis (Pallas, 1773) (Coleoptera: Coccinellidae) (Cottrell y Shapiro-Ilan 2003; Roy et al. 2008) lo que confirma la especificidad de este hongo. De esta manera, es posible suponer que los depredadores presentes en la parcela tratada con B. bassiana y que no fueron afectados por el mismo, pueden seguir ejerciendo un control sobre la plaga una vez haya terminado el efecto del hongo sobre esta.
La cepa B afectó a M. velezangeli y causó mortalidades de alrededor de 85 % en condiciones controladas de laboratorio, en tanto en condiciones de campo, la cepa también causó mortalidad sobre los insectos al disminuir los porcentajes de infestación y los daños en los árboles. El uso de esta cepa en condiciones de campo controló el insecto de una manera más efectiva que el insecticida de síntesis química, convirtiéndose en una nueva estrategia para ser usada en el control de este insecto. Además, debido a que M. velezangeli afecta otros cultivos agrícolas como es el caso de aguacate, cacao y guayaba, el uso de esta cepa podría ser evaluado en estos cultivos con el propósito de realizar un mejor control del insecto.

