











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introduction
The African oil palm, Elaeis guineensis Jacq., 1897 (Arecaceae) was the main commercial crop in the Pacific lowlands around Tumaco in Nariño, Department of Colombia, until the arrival of bud rot led to the eradication of about 35,000 hectares of the crop (Corredor Ríos et al. 2008; Preciado et al. 2011). As a solution to the disease problem, oil palm breeders produced a hybrid (OxG) between the native congeneric species, Elaeis oleifera (Kunth) Cortés, 1897 and E. guineensis, with the aim to transfer the disease tolerance of the native species to the hybrid and thus overcome the problem. Since 2010, more than 10,000 hectares in the Tumaco area were replanted with OxG hybrids (Fedepalma 2015) and until now, these seem to resist the disease. However, other problems have arisen, one of these is a lepidopteran root borer, Sagalassa valida Walker, 1856 (Lepidoptera: Brachodidae) that affects the palms mainly during the establishment phase. This insect is known in many countries producing oil palm in South and Central America (Genty 1977; Genty et al. 1978) and is present in all areas producing oil palm in Colombia (Sáenz and Olivares 2008). Particularly in the Pacific area, it is considered a pest of economic importance (Peña Rojas and Jiménez Ochoa 1994; Pinzón Andersson 1995; Aldana de la Torre et al. 2000).
Chemical control of the pest is possible but expensive (Bernal et al. 2015), so other ways of handling the problem were looked for. The role of ants in limiting the root borer was investigated by Coral et al. (2004) who associated two species of Ponerinae, Pachycondyla harpax F., 1804 and P. obscuricornis Emery, 1890 (Hymenoptera: Formicidae) with reduced borer damage. However, the importance of ants as pest control agents has been attributed to dominant or co-dominant ant species (Fernández 1991; Way and Khoo 1992; López and Potter 2000; Gallego Ropero and Armbrecht 2005), and Ponerinae generally have small colonies and an unequal distribution (Baena 1993). Therefore, we were interested in knowing a) whether there was any ground predator species that could play a role in controlling the oil palm root borer; b) the effect of land use on their distribution and population; and c) the effect of rainfall and temperature on their activity and permanence. In case a ground-active predator was identified, further studies would be required to estimate its effect on the root borer and how it could be enhanced.
Materials and methods
Study site.
All collections were performed at the research center “El Mira” (1º32’58”N 78º41’21”W) of the Colombian Corporation of Agricultural Research, located 38 km southeast of the municipality of Tumaco (Nariño, Colombia). The center is located at an altitude of 16 masl, with average temperature of 25.5 °C and mean annual precipitation of 3,067 mm (Reyes 2012). It comprises of 564 ha of land dedicated to research in oil and peach palm (Bactris gasipaes Kunth), cacao Theobroma cacao L. and non-timber forestry products. The whole farm is in an area with high soil water table and prone to flooding after heavy rainfall. Rainfall data were obtained from the weather station at El Mira Research Center and temperature data from the weather station at Tumaco airport.
The collections were made in four areas, three contiguous and one at a distance of about 1 km. Of the contiguous areas, one was a secondary forest regrowth, ten years of age and comprising of 14 ha, mainly populated by Cecropia sp. The neighboring area was a juvenile (3 years at the onset of the collections) hybrid (OxG) oil palm plantation of 14 ha, next to the peach palm germplasm bank (9.9 ha, planted in 1997). The fourth area of 3 ha at about 1 km distance was planted to OxG hybrids seven years of age when the experiment was set up. In the younger oil palm plantation, Engeo TM247SC (thiametoxam and lambda-cyhalothrin) had been applied on February 16, 17/2016 (2.5 ml/l, soil application with knapsack sprayer) three months before the onset of our collections.
Layout of pitfall trapping.
A transect of 100 m length was established with ten pitfall traps set at 10 m distance in each of the four described areas. The traps consisted of small (150 ml) transparent plastic cups, two of which were stacked and buried to leave the upper rim of the top cup level with the soil surface. Both cups remained in place between the trapping events, the top cup covered with a lid. A roof made of three bamboo skewers (300 mm long) and a 250 mm diameter styrofoam plate was placed over the trap to avoid flooding during heavy rain. For the fortnightly trapping events, the top cup was replaced by a cup filled with 50 ml water with a drop of dish-washing liquid to eliminate the surface tension. After 24 hours the collection cup was removed and replaced by the one with a cover. Collections started in May 2016 and in the course of one year, 22 collections were made with two collections per month except August 2016 and February 2017 with only one collection.
Processing of samples.
The samples were taken to the lab and sorted under a stereomicroscope. The working team had a one-week hands-on training in ant taxonomy by a professional myrmecologist to whom also questionable specimens were sent for confirmation of the identification. Ants were separated into E. ruidum and others, and a sample of E. ruidum and all other ants were kept in 70 % alcohol for later identification. All other arthropods were identified to order and family level wherever possible, recorded directly, and then discarded because of lack of space and containers. All ants except E. ruidum were conserved in numbered and labeled vials in 70 % alcohol.
Data analysis.
The data of the collections did not conform to a normal distribution and were therefore analyzed with the nonparametric Kruskall Wallis test. When significant differences in the dataset were detected, the means were separated following the method of Dunn (1961) with a 5 % level of significance.
El Mira Research Center has a very high soil water table and in addition is occasionally affected by heavy floods. As this can be expected to affect the permanence and number of soil-nesting ants and other terrestrial arthropods, a linear correlation (Pearson 1895) was calculated between rainfall and the captures in all transects. The following scenarios were considered: the accumulated precipitation between subsequent collections, accumulated precipitation during the three days before the collections and rainfall of the day the samples were recovered from the field. In order to assess the impact of the floods, the numbers of all arthropods and ants in the collection before the flood events were compared with the collection immediately afterwards. Floods for this purpose were defined as > 100 mm of rain in a day. Similarly, a linear correlation was calculated between maximum and minimum temperatures the day of placement of the pitfall traps and the average temperature calculated the day of collection of the pitfall traps.
The independence of individual trap catches was analyzed by comparing the numbers of ants captured by all traps within a transect with the hypothesis that the outer traps capture more because of the lack of competing traps on one side. The ant numbers were verified for normal distribution and the differences analyzed with the Kruskall-Wallis test as described before.
Results
Taxa captured.
In 22 pitfall trapping events covering a whole year, a total of 50,603 arthropods were captured (Table 1) of which 18,706 were ants (37.0 %) (Table 2). In addition to the ants, a wide variety of arthropods were collected in the traps. These were grouped according to order (Diptera, Hemiptera, Coleoptera, Orthoptera, Araneae, Acari and Collembola), the remainder was lumped together under “other arthropods”. Collembola were the most abundant (17,929 or 35.4 % of the collected specimens), followed by Acari (5,349 or 10.6 %), Coleoptera (3,538 or 7.0 %), Diptera (1,366 or 2.7 %), Hemiptera (1,262 or 2.5 %) and Araneae (1,259 or 2.5 %) (Table 1). Orthoptera and the remaining arthropods accounted for only 1.1 and 1.2 % of the collected specimens, respectively.
Table 1 Pitfall trap collections of terrestrial arthropods in areas under different land use; El Mira Research Centre, Tumaco, Nariño, Pacific coast of Colombia. From April 2016 to March 2017, 22 collections of ten pitfall traps were made in each area.
| Total number of collected specimens * | Diptera | Hemiptera | Coleoptera | Araneae | Acari | Collembola | Orthoptera | All other arthropods | |
|---|---|---|---|---|---|---|---|---|---|
| Secondary forest | |||||||||
| Mean ± SD | 1073.7 ± 265.1 B | 53.2 ± 19.8 A | 46.6 ± 13.2 A | 139.8 ± 30.6 A | 19.0 ± 11.5 C | 115.4 ± 104.6 B | 250.8 ± 98.6 D | 24.6 ± 9.0 A | 20.7 ± 9.9 |
| Peach palm | |||||||||
| Mean ± SD | 1252.2 ± 177.4 B | 35.0 ± 13.0 A | 41.2 ± 13.9 A | 104.4 ± 32.5 B | 58.5 ± 34.8 A | 132.2 ± 43.6 A | 505.4 ± 182.7 B | 14.4 ± 5.2 B | 16.4 ± 3.7 |
| Hybrid oil palm, 7 years | |||||||||
| Mean ± SD | 1616,4 ± 625.5 A | 22.9 ± 9.3 B | 27.4 ± 11.1 B | 75.5 ± 23.8 C | 24.8 ± 15.3 B | 148.7 ± 65.7 A | 614.0 ± 281.3 A | 12.1 ± 5.8 B | 17.5 ± 8.9 |
| Hybrid oil palm, 3 years | |||||||||
| Mean ± SD | 1119.1 ± 573.3 B | 22.5 ± 12.9 B | 11.7 ± 3.1 C | 34.1 ± 8.9 D | 23.6 ± 12.7 B | 138.6 ± 48.0 A | 422.7 ± 466.5 C | 6.0 ± 6.0 C | 10.0 ± 5.1 |
| Total | 50,613 * | 1,366 | 1,296 | 3,538 | 1,259 | 5,349 | 17,929 | 571 | 646 |
*Includes 18,812 ant specimens; values in columns with different letters are significantly different at P = 5 % or below
Table 2 Ants collected in 22 pitfall trap collections in a 100 m transect in areas under different land use: El Mira Research Centre, Tumaco, Pacific coast of Colombia, April 2016 to May 2017. Data shown are averages of 10 pitfall traps per transect.
| All ants | Ectatomma spp. * | Labidus spp. ** | |
|---|---|---|---|
| Secondary forest | |||
| Mean ± SD | 403.6 ± 200.4 B | 279.4 ± 139.8 B | 62.5 ± 70.4 A |
| Peach palm | |||
| Mean ± SD | 349.0 ± 113.1 B | 256.7 ± 108.1 B | 53.8 ± 49.1 A |
| Hybrid oil palm, 7 years | |||
| Mean ± SD | 672.8 ± 406.6 A | 638.5 ± 410.2 A | 10.3 ± 30.2 B |
| Hybrid oil palm, 3 years | |||
| Mean ± SD | 458.7 ± 102.5 AB | 424.7 ± 106.0 A | 6.6 ± 13.3 B |
| Total | 18,841 | 15,993 | 1,332 |
* > 95 % Ectatomma ruidum (Roger); ** 94.5 % Labidus praedator (Smith, 1858), 5.4 % Labidus coecus (Latreille, 1802); values in columns with different letters are significantly different at P = 5 % or below.
A breakdown of the main orders to family level gave the following results: in Diptera, 41.2 % were Drosophilidae and 5.5 % Tephritidae, in Hemiptera 20.6 % were Cydnidae and in Coleoptera 78.2 % Bostrichidae, 8.5 % Nitidulidae, and 8.1 % Staphylinidae.
Ectatomma ruidum (Roger, 1860) (Hymenoptera, Formicidae) was the most abundant and consistently present ant species throughout the whole collection period with 15,450 specimens, 84.0 % of all ants and 30.4 % of all specimens collected. It was absent in only four (of a total of 220) traps in the secondary forest, in 13 of the peach palm plantation, and three and none of the younger and older hybrid oil palm plantation, respectively. The second most frequent group were army ants of the genus Labidus with a total of 1,332 (7.1 % of ants and 2.6 % of the whole collection) specimens (Table 2).
Effects of land use.
The highest total number of specimens originated in the older hybrid oil palm plantation (χ2 = 9.0111, df = 3, P = 0.02914). Overall specimen numbers were higher in the palm plantations than in the secondary forest, but the differences were only significant for the older hybrid plantation (Table 1). Captures of Diptera and Hemiptera were significantly higher in the secondary forest and the peach palm plantation than in the oil palm transects (χ2 = 17.646, df = 3, P < 0.0005 and χ2 = 25.326, df = 3, P = 1.32e-05, respectively) (Table 1). Coleoptera and Orthoptera numbers were significantly more numerous in the secondary forest than in the other transects (χ2 = 28.388, df = 3, P < 3.01e-06 and χ2 = 23.397, df = 3, P < 3.337e-05, respectively). The highest captures of Acari were recorded from the three palm transects (χ2 = 5.9925, df = 3, P = 0.112, not significant), the highest record of Araneae was registered in peach palm with the forest and oil palm transects significantly lower (χ2 = 15.526, df = 3, P < 0.0014). Collembola were recorded in only 19 capture events because of lack of manpower for the counts of the last three collections. Nevertheless, they were the group with the second highest record and with large differences between older oil palm transects (χ2 = 17.947, df = 3, P = 0.00045) followed by peach palm, younger oil palm and forest transects, all differences significant.
Diptera and Coleoptera were captured in all traps and collections, spiders and mites were also present in most collections. In contrast, Collembola numbers fluctuated widely (between zero and 1,327 in one single trap), and total capture was much higher in all palm plantations than in the secondary forest (Table 1).
The total number of arthropods captured showed several distinct peaks, particularly in September 2016 (all transects), November 2016 (peach palm) and December 2016 (older oil palm hybrid). The greatest source for this variability were Collembola: on September 6th (767 in peach palm, 1,393 in the older oil palm and 1,542 in the younger oil palm transects); on November 11th (1,208 in peach palm) and December 14th (1,434 in the older oil palm). When these were removed from the records, the variability in the whole dataset was much reduced.
While E. ruidum was present in almost all traps and collection events, Labidus spp. were highly concentrated in nine raid episodes (Table 3). Army ant raids (defined as events with 50 or more Labidus sp. in any one pitfall trap) occurred on nine occasions, most frequently in the secondary forest transect (five occasions), twice in the peach palm plantation and once each in the hybrid oil palm plantations. The highest number of army ants caught in any single trap were 160 L. praedator in the peach palm transect and 289 if the collection of three neighboring traps of the same date are pooled.
Table 3 Army ant raids recorded during one year in pitfall traps in four transects in areas under different land use. El Mira Research Center, Tumaco, Nariño, Pacific coast of Colombia, May 2016 - April 2017.
| Date | Trap position | Army ant species | Number of ants caught |
|---|---|---|---|
| 12-may-2016 | Secondary forest, traps 2,4,7,8 | Labidus coecus | 100 |
| 26-may-2016 | Secondary forest, traps 7,8,9 | Labidus praedator | 111 |
| 30-jun-2016 | Hybrid oil palm plantation, traps 8,10 | Labidus praedator | 62 |
| 30-jun-2016 | Peach palm plantation, traps 5,6 | Labidus praedator | 50 |
| 14-jul-2016 | Secondary forest, trap 2 | Labidus praedator | 48 |
| 06-sep-2016 | Peach palm plantation, traps 4-9 | Labidus praedator | 401 |
| 06-sep-2016 | Secondary forest, traps 4, 5, 6 | Labidus praedator | 153 |
| 25-oct-2016 | Hybrid oil palm plantation, trap 9 | Labidus praedator | 51 |
| 14-mar-2017 | Secondary forest, traps 4,5 | Labidus praedator | 88 |
The composition of the ant fauna was also significantly different (χ2 = 14.91, df 3, P < 0.01) between the secondary forest (69.2 % E. ruidum) and peach palm (73.6 %) and the two oil palm transects (94.6 % E. ruidum older and 92.6 % younger plantation).
Of a total of 880 pitfall traps deployed, only six did not yield results; four were damaged in transport (14 July 2016, P20T1, T2, T4, T5) and two (27 July 2016, P18T9, P18T10) were destroyed by hoofprints of horses.
Within transect variation.
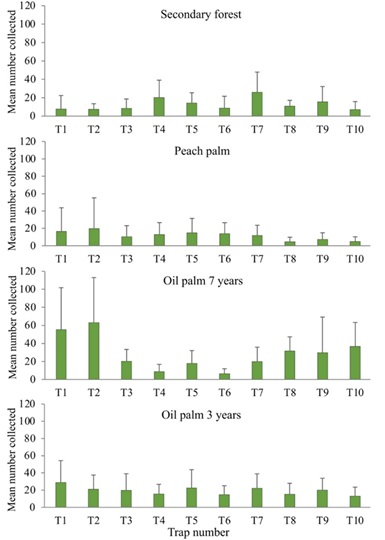
A statistical analysis of the variation of ant captures within the transects (Fig. 1) revealed differences between individual traps in the secondary forest (χ2= 46.28, df = 9, P < 0.000006), peach palm (χ2= 19,859, df = 9, P < 0.019), oil palm of seven years (χ2= 93.578, df = 9, P < 3.109 e-16), with the exception of the oil palm of three years (χ2 = 12.406, df = 9, P < 0.1914). However, these differences were trap-specific and did not depend on trap position as the first and last trapping positions did not consistently capture higher numbers despite not having competing traps on one side.
Influence of weather conditions on captures.
Total rainfall during the collection period was 3,491 mm and the average temperature was 26.3 °C (Fig. 2). Exceptional rainfall, followed by flooding of all transects, was registered on three occasions: May 14th 2016 (118.6 mm), July 21st 2016 (104 mm) and January 20th 2017 (100.2 mm). Correlation coefficients between rainfall and arthropod captures in all transects were low and did not surpass 0.53, no matter which rainfall period before or on the day of collection was tested. Even though some of the correlation coefficients were significant, the variability in the whole dataset does not allow for a conclusive statement on the relation between accumulated rainfall and arthropod captures. Even three events of total flooding of the fields failed to conclusively impact arthropod captures. While arthropod captures in the secondary forest were significantly reduced after the flooding in two of three and the E. ruidum captures in all three occasions, the effect in the palm plots was anything but uniform (Table 4). In addition, similar and even considerably higher variability between two subsequent collections occurred in all transects in absence of rainfall extremes. This was registered on eleven occasions, two of which in the secondary forest and with larger differences than those observed after flooding events.
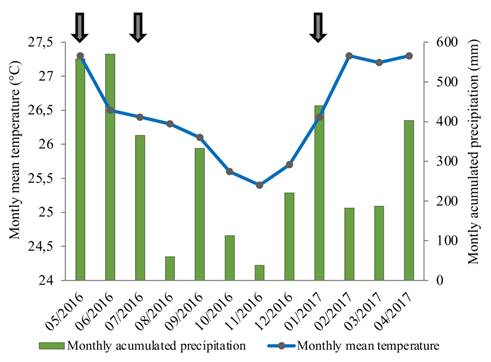
Figure 2 Monthly rainfall and mean temperatures at El Mira Research Center, Tumaco, Nariño, Pacific coast of Colombia (May 2016 - April 2017). Rainfall in excess of 100 mm per day marked with arrows.
Table 4 Effect of flooding on arthropod captures in pitfall traps in four land-use systems at the Colombian Pacific coast.
| All arthropods | Ectatomma ruidum | |||
|---|---|---|---|---|
| Date of flooding | Before flooding | After flooding | Before flooding | After flooding |
| Secondary forest | ||||
| 14-mar-2016 | 689 | 287 | 283 | 129 |
| 21-jun-2016 | 452 | 493 | 194 | 39 |
| 20-jan-2017 | 547 | 477 | 205 | 136 |
| Peach palm plantation | ||||
| 14-mar-2016 | 348 | 256 | 237 | 54 |
| 21-jun-2016 | 890 | 550 | 465 | 40 |
| 20-jan-2017 | 345 | 583 | 67 | 210 |
| Hybrid oil palm 3 years | ||||
| 14-mar-2016 | 475 | 521 | 396 | 421 |
| 21-jun-2016 | 360 | 543 | 204 | 277 |
| 20-jan-2017 | 408 | 386 | 77 | 205 |
| Hybrid oil palm 7 years | ||||
| 14-mar2016 | 742 | 525 | 595 | 248 |
| 21-jun-2016 | 874 | 615 | 502 | 178 |
| 20-jan-2017 | 382 | 445 | 209 | 272 |
Temperature throughout the whole year of collections fluctuated between 32.9 °C and 21.9 °C, well within the optimal range for tropical insects, and thus had very little effect on the activity of all arthropods. This is reflected in the correlation coefficients between temperature and arthropod numbers captured in the pitfall traps. The highest value calculated was 0.29 (Collembola, minimum temperature, secondary forest) and almost all correlation coefficients were not significant.
Discussion
Arthropods are key to the maintenance of ecosystem functions (Weisser and Siemann 2004), such as pollination, leaf litter and dung decomposition or biological control of pests (Letourneau et al. 2009). Changes in terrestrial arthropod fauna following changes in land use showed that the reduction of plant diversity has serious consequences for arthropod diversity (Siemann et al. 1998; Goehring et al. 2002). More specifically, the conversion of primary forest to oil palm plantations generally has a negative impact on the richness and diversity of arthropod taxa including beetles (Chung et al. 2000; Davis and Philips 2005) and ants (Pfeiffer et al. 2008).
It is difficult to draw conclusions about species diversity from our collections because our records are of abundance of specimens not species. Nevertheless, some insight can be gained from our data. Even though the highest number of Coleoptera were captured in the secondary forest, the differences to the more disturbed areas were moderate. Differences were almost entirely due to a single family, Bostrychidae, borers and decomposers of woody material, which comprised between 84.9 and 66.3 % of the Coleoptera collection in the four transects. The second most abundant family was Nitidulidae, decomposers of fallen fruits or in the case of young oil palms, of fruits not harvested; followed by Staphylinidae, a family of small predatory beetles. The almost total absence of Carabidae, a typical predatory species on the soil surface, is surprising with only 21 specimens in all collections combined.
Diptera, Hemiptera, and Orthoptera were more numerous in the secondary forest, whereas Acari, Araneae, and Collembola were more numerous in the palm transects. Particularly the latter were exceptionally abundant during several peaks, the cause of which is unclear. Another significant difference was the lower numbers of E. ruidum in the secondary forest, probably an effect of the higher canopy density and the resulting shade in the forest (McGlynn et al 2010). The reduced dominance of E. ruidum among the ants and the higher abundance of Coleoptera, Diptera, Hemiptera as well as Orthoptera in the forest transect might be interpreted as a partial return of the original diversity. Nevertheless, ten years of forest regrowth appears to be insufficient to reestablish a terrestrial arthropod community very distinct from the more disturbed areas.
The activity of the terrestrial arthropods as documented by the captures in the pitfall traps did not vary with the amount of rainfall. This is in stark contrast to observations of Nunes et al. (2011) and Delsinne et al. (2008), where ant numbers in semi-arid areas (Brazilian Caatinga and Paraguayan Chaco, respectively) were drastically reduced by rainfall. However, this could be expected, as our study area is part of the Chocó biogeographic region with rainfall records of up to 12,000 mm. Furthermore, Lachaud (1990) stated that foraging activity of E. ruidum never completely stopped and resumed immediately after heavy rain. In our case, even complete flooding of the study area at three occasions did not produce higher levels of variation in the captures than those observed between captures without excessive rainfall. Temperature could also not be related to the variations in activity of any of the arthropods and the range was optimal for E. ruidum (Bestelmeyer 2008).
What stood out most in our collections, are the predatory ants. The absolute dominance of E. ruidum surprises, and the fact that this species was absent in only 20 out of 880 capture events confirms its ubiquity and the stability of its populations over time. The results of our study indicate that E. ruidum is a dominant species an important attribute of ants as agents for the natural control of insect pests (Majer 1972, 1976; Leston 1973, 1978; Way and Khoo 1992).
Ectatomma ruidum is mentioned as the most frequent and active predatory ant in oil palm cultivation (Zenner de Polania 1994). In the same study, Ectatomma quadridens (F. 1793) is mentioned as a predator of S. valida. In Colombia, E. ruidum has also been associated with predation of spittle bugs in pastures, insect pests of citrus (Abadia Lozano et al. 2013) and various pests of coffee (Mera Velasco et al.2010). Velázquez et al. (2006) identified E. ruidum among the important predators of the agave weevil, Scyphophorus acupunctatus Gyllenhal, 1838 (Coleoptera: Curculionidae) in Venezuela with presence in almost all the sampled areas. The species has also been proposed as biological control agent of the banana weevil, Cosmopolites sordidus Germar, 1824 (Coleoptera: Curculionidae) in plantations of Musa spp. in Miranda State of Venezuela (Goitia and Cerdá 1998).
In previous studies in oil palm plantations of the Pacific region, Coral et al. (2004) associated two species of hunting ants, P. harpax and P. obscuricornis, to a low level of damage by S.valida. However, in our collections that covered four different environments and a considerably longer sampling time, only 25 specimens of this genus were collected. Apparently, Pachycondyla spp. depend on the microhabitat of large amounts of organic matter to nest, which was not available in our plots and in its absence, the species did not establish in high numbers. In addition, there are doubts about the preference of this genus for S. valida larvae (Sarmiento et al. 2005).
Ectatomma ruidum is a species with a wide geographical distribution, preferentially inhabiting the moist neotropical forests. This ant has a high trophic versatility that might be the reason for its wide distribution (Riera-Valera and Pérez-Sanchez 2009). Ectatomma ruidum workers have foraging habits for live prey, where they sting and carry their prey to the nest (Schatz et al. 1996).
Ectatomma ruidum has been categorized as a forest species by some authors, while others claim that it is a species adapted to disturbed areas (Fernández 1991; Zenner de Polania 1994). Our data indicate that the numbers are consistently higher in the most disturbed areas, which would support the second claim. The species is also known as a thermophile, since light-exposed areas generally have higher colony densities than adjacent shaded areas or forests with closed canopies (Lachaud 1990; Schatz and Lachaud 2008; Santamaría et al. 2009; McGlynn et al. 2010). This would explain its high abundance in the oil palm plantations and the relative scarcity in the secondary forest.
Wang and Foster (2015) have indicated that species composition of ground-foraging ant communities varies with different ages of oil palm, where older plantations show greater species homogeneity, with specialist predatory ants more abundant, as observed with E. ruidum in our study.
The comparison of individual ant catches within each transect was made to demonstrate the independence of trap catches. Only two outer traps of eight (at the end of the older oil palm transect) but also four interior traps had higher captures, which appears to confirm the independence of each trap.
Conclusions
In the present study, we show that the ants are the prevalent and most stable of the terrestrial arthropod communities and that Ectatomma ruidum is the dominant species in the four areas studied. Therefore, we can follow Lachaud who stated “E. ruidum is mainly carnivorous and considering the high density of nests of this species, its role as an element of biological control seems to be undeniable”. We are convinced we have identified a terrestrial predator with the potential to provide the required pest control services in oil palm plantations at the Pacific coast. Further studies are necessary to elucidate the effectiveness of E. ruidum in removing root borer larvae and ways and means of its conservation and stimulation. We can also conclude that ten years of secondary forest recovery lead to measurable differences in arthropod diversity as compared to managed areas.

