










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Latinoamericana de Psicología
Print version ISSN 0120-0534
rev.latinoam.psicol. vol.39 no.3 Bogotá Sept./Dec. 2007
ARTÍCULOS
EL ESTUDIO DEL CONTRASTE ENTRE INCENTIVOS: LA CONTRIBUCIÓN DE ABRAM AMSEL
THE STUDY OF INCENTIVE CONTRAST: THE CONTRIBUTION OF ABRAHAM AMSEL
LEONARDO A. ORTEGA
y
MAURICIO R. PAPINI1
Texas Christian University, USA
1Correspondencia: MAURICIO R. PAPINI, Department of Psychology, Texas Christian University Box 298920, Fort Worth, TX 76129, USA. Correo electrónico: m.papini@tcu.edu
ABSTRACT
This article describes Abram Amsel's [1922-2006] contributions to the study of learning in situations involving surprising reward omissions. Emphasis is placed on recent developments in the study of consummatory successive negative contrast. Amsel's model of psychobiological research involves an integration of behavioral theory with neurobiological and developmental approaches. It is suggested that such a model is often implicit in current research programs that are advancing our understanding of some of the central phenomena in basic psychological theory, including learning, emotion, and conflict.Key Words: paradoxical reward effects, successive negative contrast, opioid system.
RESUMEN
Este artículo describe las contribuciones de Abram Amsel [1922-2006] al estudio del aprendizaje en situaciones que involucran la omisión sorpresiva de incentivos. Se enfatizan algunos desarrollos recientes en el estudio del contraste sucesivo negativo consumatorio. Amsel propuso un modelo de la investigación psicobiológica basado en una integración de la teoría conductual con enfoques neurobiológicos y del desarrollo. Se sugiere aquí que dicho modelo está implícito en muchas líneas actuales de investigación que están avanzando nuestra comprensión de algunos fenómenos centrales de la psicología, tales como el aprendizaje, la emoción y el conflicto.
Palabras clave: fenómenos paradójicos de reforzamiento, contraste sucesivo negativo, sistema opioide. científica.
PASOS EN LA INVESTIGACIÓN PSICOBIOLÓGICA DEL APRENDIZAJE
La comprensión de los fenómenos psico-lógicos requiere de una investigación dirigida a diferentes niveles causales, comenzando por regularidades de comportamiento (el nivel de análisis tradicionalmente psicológico), y siguiendo por la identificación de los circuitos nerviosos, los sistemas neuroquímicos y los procesos celulares involucrados. Teniendo en cuenta la íntima relación entre comportamiento y biolo-gía, es de especial importancia la colaboración entre la psicología y la neurociencia. A comien-zos del Siglo XXI, este enfoque integrador está en su infancia y su desarrollo depende del esfuerzo de científicos capaces de extender su trabajo en dos o más niveles de análisis. Entre los pioneros se encuentra Abram Amsel [1922-2006], un investigador cuyas contribuciones sirven como modelo y guía al trabajo interdisciplinario que caracteriza el presente de la psicología y que será sin dudas de fundamental importancia en el futuro inmediato.
Amsel (1992) propuso estudiar el aprendizaje asociativo partiendo de hallazgos comportamentales sintetizados en una teoría psicológica, para luego establecer conexiones con niveles biológicos de causalidad. Esta estrategia esta dando sus frutos en una variedad de áreas de trabajo, incluyendo el condicionamiento clásico de miedo (Davis, 2006; Fanselow & Poulos, 2005), el condicionamiento instrumen-tal apetitivo (Balleine & Killcross, 2006) y las bases celulares del condicionamiento en invertebrados (Giles, Rose, & Rankin, 2006). Amsel (1992) propuso una serie de seis pasos que se utilizarán a continuación como esquema organizativo del resto del este artículo.
El primer paso es la observación y descripción de una familia de fenómenos comportamentales aparentemente relacionados. Amsel concentró su esfuerzo en una serie de efectos que involucran la pérdida sorpresiva de recompensas y que colectiva-mente denominó fenómenos paradójicos de reforzamiento (FPR). Estos fenómenos se denominan “paradójicos” pués se caracterizan por producir menor nivel conductual en respuestas que han sido reforzadas más frecuentemente o por recompensas de mayor magnitud. Cada uno de estos FPRs involucra uno o más cambios sorpresivos en el tamaño o calidad de la recompensa. Por ejemplo, en el efecto en la extinción del reforzamiento parcial (EERP), una respuesta reforzada en algunos ensayos y no en otros, mezclados al azar, exhibe un alto grado de persistencia durante la extinción (i.e., cuando ninguna ocurrencia de la respuesta es reforzada), en relación con una respuesta sometida a reforzamiento continuo. Como el organismo carece de señales que le permitan predecir si un ensayo determinado será reforzado, tanto el reforzamiento como su omisión son sorpresivos. El EERP permite estudiar los ajustes de comportamiento en situaciones de incertidumbre. Otro ejemplo de un FPR es el contraste sucesivo negativo (CSN), en el que la respuesta a un incentivo de baja magnitud es mucho menor cuando esta respuesta fue previamente reforzada por un incentivo de gran magnitud, que en un grupo control siempre reforzado con la magnitud baja. El CSN sugiere que el comportamiento actual depende de un proceso de comparación entre expectativas de incentivo e incentivos presentes en una determinada situación.
El segundo paso implica la conceptualización de los fenómenos comportamentales en términos de una teoría psicológica. Una de las contribuciones más significativas de Amsel (1958, 1962, 1967, 1972, 1992) a la psicología fue su teoría de la frustración. Trabajando dentro de un enfoque neoconductista, Amsel (1989) distinguió entre la reacción incondicionada a la pérdida sorpresiva del reforzamiento y la reacción mygdaleorio que involucra el desarrollo de expectativas. Denominó a estas dos reacciones, respectivamente, frustración primaria (FP) y secundaria (FS). En la teoría de la frustración, la FP y FS son variables intervinientes cuyas propiedades conducen a una serie de predicciones originales que vienen estimulando una enorme cantidad de investigación desde la década de los 1960s. En su aplicación al CSN, la teoría de la frustración sugiere que la violación de la expectativa del incentivo de mayor magnitud produce FP, un estado emocional aversivo. En ensayos subsecuentes, los estímulos asociados con la FP pueden inducir FS, es decir, la expectativa de que la respuesta será nuevamente seguida por una reacción emocional aversiva de frustración. La FS induce respuestas de evitación, lo que lleva a un conflicto de acercamiento-evitación y a la consecuente degradación del comporta-miento típica del CSN.
El tercer paso consiste en estudiar el desarrollo ontogenético de los fenómenos comportamentales en cuestión. Este enfoque ontogenético tiene dos metas. Primero, determinar la presencia o ausencia de cada fenómeno en cada estadio del desarrollo, y segundo, establecer el orden en que los diferentes fenómenos emergen durante el desarrollo. Amsel dedicó la última parte de su actividad científica al estudio del desarrollo de los FPRs en la rata (Amsel & Stanton, 1980). El trabajo experimental eventualmente se concentró en el período postnatal que va desde los 10 a los 25 días de vida, cuando el sistema nervioso está madurando rápidamente (en la rata, el sistema nervioso central termina de madurar alrededor de los 45 días de vida). Este trabajo demostró que antes de los 11 días de vida, la rata utiliza mecanismos asociativos de aprendizaje, pero no exhibe FPRs. Tales fenómenos se desarrollan en una secuencia que comienza con el EERP, que aparece a los 12-13 días de vida, y termina con el CSN, que aparece recién a los 24-25 días de vida. Amsel (1992) conceptualizó la primera transición como una que involucra dife-rentes niveles de funcionamiento. Antes de los 11 días de edad, la rata opera en base a un mecanismo no paradójico según el cual el reforzamiento fortalece las asociaciones y el no reforzamiento las debilita. Este nivel opera de una manera similar a lo que describen algunas teorías clásicas, tales como las de Thorndike (1911), Hull (1943), y Rescorla y Wagner (1972). La aparición de los FRs representa el control de la conducta por las expectativas de incentivo, indicando la activación de los mecanismos paradójicos.
El cuarto paso comprende la vinculación entre procesos neurales y los fenómenos comportamentales. Específicamente, los FPRs están relacionados con el funcionamiento de lo que Gray (1987) denominó el sistema de inhibi-ción comportamental. De acuerdo con Gray, diferentes estructuras cerebrales particulares están interconectadas de manera que producen inhibición comportamenteal y facilitan el cambio comportamental en situaciones que involucran estímulos que están en conflicto. Siguiendo esta formulación, Amsel propuso que las interacciones entre la función septo-hipocampal y otras áreas del sistema límbico son las encargadas de modular y modificar los comportamientos en situaciones de cambio sorpresivo en las condiciones de recompensa. Esta idea ha tenido un amplio poder heurístico al permitir el diseño de pruebas que evalúan las bases neurobiológicas del comportamiento en los FPRs. Adicionalmente, como se describirá posterior-mente, ha permitido relacionar el CSN con otros sistemas biológicos, específicamente el sistema opioide.
El quinto paso consiste en relacionar la presencia o ausencia de los fenómenos comportamentales con el estado de maduración del sistema nervioso durante el mismo período de desarrollo. En el caso de los FPRs en la rata, durante el período en el que experimentan un desarrollo (11-25 días de vida) a nivel conductual, algunas áreas del cerebro están también madurando rápidamente. Entre estas áreas esta la formación hipocámpica que contiene dos componentes principales: células piramidales y células granulares. Numerosos estudios demuestran que las células piramidales maduran en el embrión, mientras que las granulares maduran principalmente (aunque no únicamente) durante el período que va de los 5 a los 30 días de vida.
El sexto y último paso es de revisión y modificación de la teoría psicológica utilizada teniendo en cuenta los ajustes sugeridos por los hallazgos de estudios neurobiológicos y ontogenéticos. Este paso es fundamental para el desarrollo de investigaciones interdisciplinarias por lo que se discutirá en detalle utilizando como ejemplo uno de los fenómenos comportamentales de contraste entre incentivos, el CSN. Con este objetivo, se hará una descripción del fenómeno de CSN y su interpretación desde la teoría de frustración de Amsel. Posteriormente, se describirá la literatura más reciente haciendo énfasis en la base biológica de este fenómeno. Para terminar, se revisarán algunos de los aportes de la estrategia de Amsel y las direcciones futuras de este campo de investigación.
TEORÍA DE LA FRUSTRACIÓN Y CONTRASTE SUCESIVO NEGATIVO (CSN)
Los trabajos pioneros de Pavlov (1927) y Thorndike (1911) dieron origen a dos modelos para interpretar los mecanismos psicológicos del aprendizaje: condicionamiento clásico y condicionamiento instrumental. A pesar de que originalmente se intentó hacer una distinción clara entre aprendizajes basados en uno de estos puntos de vista (e.g., Skinner, 1935), algunos experimentos sugerían que la situación es más compleja y que existen una variedad de comportamientos que dependen de la contribución simultánea de mecanismos pavlovianos e instrumentales. Por ejemplo, Elliot (1928) observó que el comportamiento de un grupo de ratas entrenadas en un laberinto complejo con una recompensa preferida se veía afectado negativamente cuando la recompensa era cambiada de manera inesperada a una recompensa menos preferida; este cambio de comportamiento posteriormente se denominaría CSN. Una explicación del CSN en términos de condicionamiento instrumental no era suficiente dado que el trabajo de Elliot (1928) sugería que los incentivos eran codificados en la estructura asociativa formada durante la adquisición (Papini, 2006). Por lo tanto, el aprendizaje no se podía reducir a la relación propuesta por Thorndike entre el comportamiento del animal y una recompensa. Igualmente, una explicación en terminos de aprendizaje de relaciones entre estímulos, más cercana a la idea pavloviana, era insuficiente porque aunque introducía la idea de la adquisición de expectativas hacia el objetivo, los mecanismos de obtención de la recompensa se reducían a tendencias generales de aproximación y evitación. En el experimento de Elliott (1928), las ratas aprendían a recorrer un laberinto sumamente complejo, con una gran cantidad de callejones sin salida, lo que sugería que la conducta iba más allá de la mera tendencia de aproximación a la meta.
Una aproximación teórica diferente fue la de incorporar de manera paralela los mecanismos de aprendizaje clásicos e instrumentales, tal como se propuso desde las denominadas teorías de los dos procesos (Mowrer, 1960; Overmier & Lawry, 1979). A partir de este tipo de modelo, así como de su extensión del concepto de reacción fraccional anticipatorio de la meta propuesta por Hull (1931), Amsel derivó una explicación general de los FPRs. De esta manera, Amsel (1958, 1962, 1992) propuso una teoría de la frustración que al incluir los aspectos clásicos e instrumentales, así como variables motivacionales, provee una integración teórica para una variedad de fenómenos comportamentales relacionados con la no-recompensa sorpresiva tales como el EERP y el CSN, entre otros. En el resto de esta sección se describe la teoría de la frustración utilizando como modelo el procedimiento de CSN consumatorio (CSNc).
En un experimento típico de CSNc, dos grupos de ratas son expuestas a beber una solución de sucrosa al 4% o al 32% mientras están bajo privación de alimentos. Luego de 10 ensayos diarios de 5 min de acceso a la solución, el grupo que recibió 32% de sucrosa experimenta una reducción en la concentración de la solución a 4% de sucrosa (Grupo 32-4). El otro grupo continúa bebiendo la misma solución de 4% de sucrosa (Grupo 4-4). Este nuevo régimen continúa duran-te 5 ensayos diarios. En cada ensayo de entrena-miento se mide el comportamiento consumatorio, registrado en términos de volumen de ingesta de fluidos, número de lamidas del tubo por donde se dispensa la solución, o tiempo acumulado de contacto con dicho tubo según el experimento. Durante el período posterior a la reducción del incentivo, ambos grupos reciben acceso a la misma solución (4% de sucrosa), pero su comportamiento es muy diferente. Los animales expuestos a una reducción del 32% al 4% de sucrosa exhiben una disminución drástica de la conducta consumatoria comparados con los animales que siempre tuvieron acceso a la solución de 4% de sucrosa. Mientras que la solución de 4% es aceptable para las ratas que sólo recibierón tal solución, la misma solución provoca un rechazo en las ratas que previamente recibieron 32% de sacarosa en esas condiciones (figura 1).
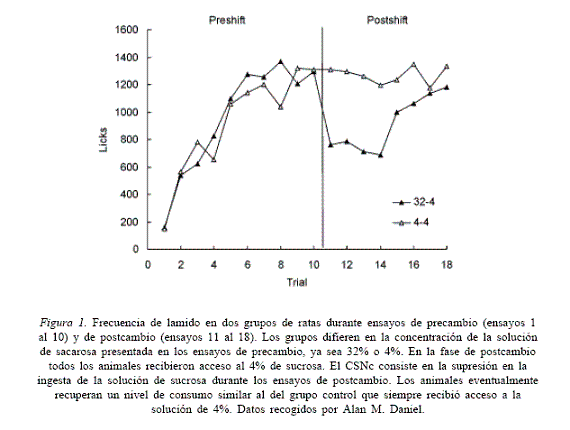
Figura 1. Frecuencia de lamido en dos grupos de ratas durante ensayos de precambio (ensayos 1 al 10) y de postcambio (ensayos 11 al 18). Los grupos difieren en la concentración de la solución de sacarosa presentada en los ensayos de precambio, ya sea 32% o 4%. En la fase de postcambio todos los animales recibieron acceso al 4% de sucrosa. El CSNc consiste en la supresión en la ingesta de la solución de sucrosa durante los ensayos de postcambio. Los animales eventualmente recuperan un nivel de consumo similar al del grupo control que siempre recibió acceso a la solución de 4%. Datos recogidos por Alan M. Daniel.
El modelo teórico de frustración desarrollado por Amsel puede ser aplicado para entender los mecanismos subyacentes al CSNc (Papini, 2003; Papini, Wood, Daniel, & Norris, 2006; Wood, Daniel, & Papini, 2005). La situación de CSNc se relaciona con mecanismos comportamentales que pueden ser descritos en tres estados teóricos generales: FP, FS y contracondicionamiento. Los cambios conductuales observados con posterioridad a la reducción del incentivo en el grupo 32-4 se dividen en aquellos que ocurren durante el primer ensayo de post-cambio (usualmente el ensayo 11) y aquellos que ocurren durante el consiguiente período de recuperación del comportamiento consumatorio (usualmente los ensayos 12-15). La supresión consumatoria que se detecta en el ensayo 11 se relaciona con la inducción de FP provocada por la discrepancia negativa entre una expectativa de 32% sucrosa y la solución del 4% efectivamente encontrada en la situación. La FP induce, a su vez, un estado emocional interno aversivo que provoca un cambio de una conducta consumatoria a una de búsqueda y evitación de la fuente original de reforzamiento. Simultáneamente, los estímulos presentes se asocian al estado interno de FP.
Las consecuencias de este nuevo aprendizaje aversivo se detectan en los siguientes ensayos. En el ensayo 12 (el segundo ensayo post-cambio), los estímulos presentes inducen expectativas opuestas. Por un lado, y debido al aprendizaje de la primera fase (ensayos 1-10), todavía existe una tendencia a acercarse al tubo que dispensa la solución de sucrosa con la expectativa de encontrar la solución original del 32%. Por otro lado, la experiencia frustrante del ensayo precedente induce la expectativa de una nueva frustración, o FS. De esta manera, la expectativa previa de recompensa junto con la nueva expectativa de frustración producen respuestas de aproximación y de evitación, respectivamente, generando un conflicto que conlleva a una competencia conductual entre tales respuestas. Tal conflicto refleja la carga ambivalente de una situación que es a la vez apetitiva y aversiva, y consiguientemente puede denominarse conflicto de acercamiento-evitación (Miller, 1944).
Varios procesos se desencadenan durante esta segunda fase de recuperación (ensayos 12-15). Por un lado, las expectativas de reforzamiento comienzan a ajustarse a las nuevas condiciones. Una expectativa cercana al 4% de sucrosa induce cada vez menos FP y por consiguiente debilita la respuesta aversiva. Pero además, el estado interno de FS comienza a ser asociado con la ingesta de 4% de sucrosa, que aunque no es tan alta como la de 32%, todavía constituye un incentivo de considerable valor, a juzgar por la conducta del grupo control. Esta asociación entre un estado aversivo y un incentivo apetitivo conducen al contracondicionamiento de la FS, la que pierde el poder de inducir respuestas de evitación. La consecuencia de estos procesos (ajuste a las nuevas condiciones de incentivo y contracondicionamiento de la FS) es la recupe-ración gradual de niveles normales de conducta consumatoria. Este mecanismo de contracondicionamiento fue de particular importancia para Amsel (1992) dado que proveyó la base para plantear el aprendizaje de disposiciones, es decir, de tendencias generales de comportamientos idiosincrásicos que se mantienen durante la vida del organismo y que le permiten desarrollar un estilo conductual independientemente de los detalles situacionales.
BASES BIOLÓGICAS DEL CONTRASTE SUCESIVO NEGATIVO CONSUMATORIO (CSNC)
Esta sección describe algunos de los trabajos que exploran los mecanismos biológicos del CSNc. En particular, se describen algunas áreas cerebrales identificadas como parte del circuito subyacente al CSNc y el trabajo que identifica al sistema opioide como uno de los substratos neuroquímicos de este fenómeno psicológico. Siguiendo la estrategia de investigación psicobiológica de Amsel y utilizando la teoría de la frustración como guía, uno esperaría que la distinción entre FP y FS sugerida con base en datos puramente conductuales, tuviera un paralelo en términos de funcionamiento cerebral. La predicción más general es que el comportamiento consumatorio que se observa durante el primer y el segundo ensayo luego de la reducción del incentivo (usualmente los ensayos 11 y 12) debiera estar regulado por diferentes circuitos o sistemas neuroquímicos, uno activado principalmente por la FP y el otro involucrado principalmente en la FS y el conflicto de acercamiento-evitación.
Identificación de las Áreas Cerebrales Involucradas en el CSNc
En lo que concierne específicamente al CSNc, la identificación del circuito cerebral subyacente comienza por notar las áreas cuyo daño afecta (o no) este fenómeno. La interpretación de los efectos de lesiones cerebrales, como cualquier otra manipulación fisiológica, está sujeta a una serie de inconvenientes. Por ejemplo, las lesiones generadas por el pasaje de una corriente eléctrica (lesiones electrolíticas) dañan no sólo las neuronas del sitio en el que se efectúa la lesión, sino también las fibras de paso. Esto complica la interpretación. De todas maneras, el método de lesionar áreas discretas del sistema nervioso y determinar su efecto en el comporta-miento es una herramienta valiosa como primer abordaje al problema de identificar los componentes del circuito cerebral.
Estudios de lesión cerebral muestran que no todas las áreas son igualmente importantes. Dos áreas en particular sobresalen porque si bien su participación en fenómenos de contraste instru-mental ha sido demostrada en varios estudios, su lesión no tiene efectos en el CSNc: el área septal y el hipocampo (Flaherty, 1996). Tres áreas cuya lesión perturba el normal desarrollo del CSNc han sido identificadas: (1) el núcleo parabraquial, (2) el tálamo gustativo y (3) la amígdala. Dado que el CSNc involucra la con-ducta de lamer un tubo que provee de una solución de sucrosa, tanto la vía gustativa como la coordinación motriz del lamido son relevantes. El estudio de la respuesta de lamido indica que el circuito cerebral puede visualizarse como un generador central de patrones organizado a nivel del tallo cerebral, con una importante contribución de la formación reticular medial y lateral (Travers, Dinardo, & Karimnamazi, 1997). Los mecanismos de contraste de incentivos pueden, a la vez, conceptualizarse como influencias de varias áreas cerebrales sobre este generador central, incluyendo el tálamo gustativo y la amígdala mencionados previamente.
La lesión del núcleo parabraquial del puente cerebral elimina el CSNc (Grigson, Spector, & Norgren, 1994). Este núcleo es parte de la vía gustativa y se activa cuando la rata esta consumiendo soluciones de sucrosa (Tabuchi et al., 2002). Además, este núcleo provee parte de la información que alimenta el generador central que coordina los patrones de lamida en la rata (Travers et al., 1997) y recibe información tanto de los receptores gustativos periféricos (vía núcleo del tracto solitario) como de la amígdala. A su vez, proyecta fibras tanto a la amígdala como al tálamo gustativo. Las lesiones parabraquiales no eliminan la habilidad de las ratas de registrar la diferencia entre una solución del 32% y una del 4% de sucrosa. Por lo tanto, el efecto de la lesión no se debe a una incapacidad de detectar el cambio, sino que refleja una reacción atenuada al mismo.
Parte del output del núcleo parabraquial alcanza el tálamo gustativo, el que a la vez envía información a la corteza gustativa y a la amígdala (Lundy & Norgren, 2004). No sorprende, entonces, que lesiones del tálamo gustativo in-terfieran con el CSNc (Reilly & Trifunovic, 1999, 2003). Sin embargo, el efecto de esta lesión es selectivo para el CSNc y no involucra el Contraste Sucesivo Negativo Instrumental (CSNi) (Sastre & Reilly, 2006). Dado que el CSNi se induce por medio de la reducción en la magnitud de alimento balanceado sólido, la implicación es que las lesiones talámicas afectan selectivamente el procesamiento de soluciones de sucrosa.
Una buena cantidad de datos confirman un papel relevante de la amígdala en CSNc. Por ejemplo, lesiones del núcleo basolateral de la amígdala reducen el CSNc sin eliminarlo completamente, mientras que lesiones del área corticomedial (que incluye el núcleo central) previenen la ocurrencia del CSNc (Becker et al., 1984). Los animales con este tipo de lesiones parecen detectar la diferencia entre las distintas concentraciones de sucrosa y responden al cam-bio de 32% a 4% disminuyendo la conducta consumatoria, pero sin demostrar contraste. La amígdala contiene una alta concentración de receptores benzodiazepínicos y opiáceos, dos clases de fármacos que reducen el CNSc (Flaherty, 1996). Por lo tanto, el efecto de las lesiones es concordante con el efecto de estos fármacos. Además, Liao y Chiang (2003) demostraron que la infusión de diazepam (un ansiolítico benzodiazepínico) en la amígdala reduce el CSNc, mientras que la misma infusión en el hipocampo carece de efectos detectables.
Además del procedimiento de lesión cerebral, existen otros métodos que permiten identificar áreas que se activan selectivamente durante un determinado fenómeno conductual y algunos de ellos han sido utilizados en el estudio del CSNc, aunque de manera restringida. Por ejemplo, el procedimiento conocido como microdiálisis permite determinar la concentración de ciertas sustancias químicas (e.g., neurotransmisores) en áreas específicas del sistema nervioso central. Aplicando esta técnica al núcleo accumbens, Genn, Ahn, y Phillips (2004) detectaron un au-mento en la secreción de dopamina en las ratas del grupo control sujeto siempre a la concentración del 4% de sucrosa. Este aumento en dopamina no fue detectado en animales que experimentaban una degradación del 32% al 4% de sucrosa. Dado que la presencia de dopamina en el núcleo accumbens se interpreta generalmente como una indicación del valor apetitivo de un incentivo, se puede concluir que la degradación del 32% al 4% va acompañada por una reducción en el valor apetitivo de la solución del 4% de sucrosa. Esta conclusión concuerda, obviamente, con la evidencia conductual.
La actividad neuronal en áreas específicas puede analizarse por medio de una técnica conocida como inmunorreactividad de c-Fos. Esta técnica utiliza un anticuerpo específico que se liga preferencialmente a la proteina c-Fos, que a su vez es un factor de transcripción genética temprana inducida por exposición a distintos estímulos (Nestler, Hyman & Malenka, 2001). Ratas ex-puestas siempre a 32% o a 4% de sucrosa, y ratas expuestas a una degradación del 32 al 4% de sucrosa fueron sacrificadas inmediatamente después del primer o segundo ensayo después del cambio y sus cerebros fueron procesados para determinar las zonas que estaban activas (Pecoraro & Dallman, 2005). Sorprendentemente, el análisis reveló una significativa elevación de la actividad neuronal en varias áreas luego del primer ensayo de reducción del incentivo, incluyendo la corteza anterior, la corteza frontal orbital, núcleo accumbens, el área septal lateral, el área medial de la amígdala, y el hipotálamo lateral, entre muchas otras áreas. Sin embargo, este análisis reveló sólo un aumento de la actividad en el área gris central del puente cerebral luego del segundo ensayo posterior al cambio de incentivo. Es claro que el análisis de c-Fos revela la activación de una multitud de áreas cerebrales, pero queda por determinar que papel juegan en la producción del fenómeno de CSNc.
En resumen, la identificación de áreas cerebrales involucradas en el CSNc está comenzando a proveer los primeros resultados del tipo de selectividad predicha por la teoría de la frustración. Esta selectividad no se ha observado, hasta ahora, en términos de lesiones cerebrales discretas, cuyos efectos tienen a reflejarse tanto en términos del impacto inicial de la degradación de incentivo, como en términos de la recuperación de la conducta consumatoria. De todas maneras, los estudios de lesión cerebral son muy pocos y cubren un número de áreas muy pequeño, a juzgar por la cantidad que se activan durante la experiencia de reducción del incentivo (Pecoraro & Dallman, 2005).
Identificación de los Sistemas Neuroquímicos Involucrados en el CSNc
Otra estrategia psicobiológica ampliamente utilizada en el estudio del CSNc es la psicofarmacológica. La literatura sobre los efectos de drogas en el CSNc es bastante amplia gracias al trabajo de Charles Flaherty y colaboradores (Flaherty, 1996). Uno de los aspectos más interesantes de esta literatura es que los efectos son selectivos. Dos clases de selectividad han sido descubiertas: (1) Sólo algunas drogas tienen efectos en el CSNc, principalmente los ansiolíticos, la corticosterona, y los opioides. (2) Además, estas drogas son también selectivas en el sentido que, al menos algunas de ellas, tienen efectos en el primer ensayo posterior a la degradación del incentivo, mientras que otras tienen efecto en el segundo ensayo (por supuesto, hay drogas que afectan ambos ensayos también). Los hallazgos mencionados en (2) serán descritos de acuerdo con la lógica de la teoría de la frustración. En general, los efectos de estas manipulaciones han iluminado los mecanismos relacionados con la FP y FS, por lo que su estudio se ha centrado en la evaluacion de distintos efectos farmacologicos en los ensayos 11 y 12 del procedimiento estandar de CSNc.
Un buen ejemplo de esto son los efectos de los ansiolíticos sobre el CSNc. Manipulaciones que involucran distintas benzodiazepinas han resultado en efectos dependientes del ensayo en las cuales son aplicadas. Aplicadas antes del ensayo 11, las benzodiazepinas no tienen efectos detectables en el CSNc. El tamaño del efecto luego de una reducción del 32% al 4% de sucrosa es virtualmente idéntico entre animales que recibieron varias dosis de benzodiazepinas o el vehículo de la droga como control. A diferencia de estos resultados, la aplicación de las mismas drogas antes del ensayo 12 reduce el tamaño del CSNc, aumentando selectivamente el consumo de sucrosa en el grupo que experimento una reducción del incentivo, sin afectar la conducta del grupo que siempre consumió la solución del 4% (e.g., Flaherty, Clark, & Coppotelli, 1996; Flaherty, Grigson, & Rowan, 1986). En términos de la teoría de la frustración, estos resultados sugieren que el efecto ansiolítico de las benzodiazepinas se observa predominantemente cuando la FS alcanza una intensidad alta, cosa que uno esperaría que tomara una cantidad mínima de experiencia con la reducción del incentivo.
Los estudios que evaluaron los niveles de corticosterona en plasma en ratas expuestas a la reducción de incentivo son también congruentes con la disociación entre la FP y FS propuesta por la teoría de la frustración. Por ejemplo, los niveles de corticosterona en plasma aumentan de manera significativa después del ensayo 12, pero no después del ensayo 11 (Flaherty, Becker, & Pohorecky, 1985; Mitchell & Flaherty, 1998). Esto sugiere que la corticosterona, una hormona que se asocia a la respuesta de estrés emocional, está vinculada a la FS y al conflicto, más que a la reacción inicial al cambio. Sugestivamente, los niveles de corticosterona aumentan también an-tes del ensayo 12, es decir, en anticipación al segundo ensayo después de la reducción del incentivo (Mitchell & Flaherty, 1998). Esto sugiere que la corticosterona podría estar involucrada en el proceso de condicionamiento aversivo que da lugar, teóricamente, al desarrollo de la FS. De acuerdo con esta hipótesis, Bentosela, Ruetti, Muzio, Mustaca, y Papini (2006) demostraron que la aplicación de corticosterona inmediata-mente después del ensayo 11 (pero no 3 h después de este ensayo) prolonga el CSNc durante los ensayos siguientes.
El papel del sistema opioide en el CSNc es sugerido por un paralelo entre, por un lado, el dolor físico y el miedo, y por el otro, la FP y la FS (Gray, 1987; Papini, 2003). El dolor físico es el que un organismo experimenta, por ejemplo, cuando hay un corte en la piel o cuando se administra una descarga eléctrica en las manos o pies. Este tipo de estímulos activan el sistema opioide, cuya función es la de regular la intensidad de la aversividad asociada con la experiencia de dolor físico. En la situación de CSNc no existe ninguna fuente de dolor físico, pero el organismo es expuesto a una experiencia de pérdida. Vista como tal, la reducción del 32% al 4% de sucrosa es para la rata una experiencia de pérdida de una recompensa de valor apetitivo que tiene consecuencias emocionales análogas a experiencias similares de pérdida en primates, tales como la separación entre madre y cría (Papini & Dudley, 1997). Es posible, entonces, reconceptualizar el CSNc como una situación que involucra una especie de dolor no-físico que puede denominar-se dolor psicológico (Papini, Wood, Daniel, & Norris, 2006).
El sistema opioide regula este dolor psicológico, tal como se estudia en la situación de CSNc, de una manera similar a la regulación del dolor físico. Los opioides son una familia de péptidos neuroactivos que actúan sobre al menos tres tipos de receptores denominados ì, ä y ê. Estos receptores están ampliamente distribuidos en el sistema nervioso central, pero su concentración es diferencial (Mansour, Fox, Akil & Watson, 1995). La predicción central, tal como en los ejemplos descritos previamente para otras drogas y para estudios de lesión y actividad neuronal, es que diferentes clases de receptores opiáceos jugarán un papel selectivo en el CSNc, aumentando o disminuyendo el tamaño del efecto conductual y teniendo estos efectos selectivamente en el ensayo 11 o el 12.
Rowan y Flaherty (1987) hallaron que la administración de morfina produce una atenuación de CSNc cuando es aplicada inmediatamente antes de los ensayos 11 ó 12. Además, el efecto atenuante de la morfina en el ensayo 12 fue eliminado al inyectar de manera concurrente naloxone. Ninguna de estas dos drogas tuvo efecto sobre la conducta consumatoria de los grupos que sólo recibieron 4% de sucrosa. La morfina es un agonista no selectivo de los tres receptores opioides, mientras que el naloxone es un antagonista no selectivo. Estos datos constituyen la evidencia inicial que el sistema opioide es capaz de regular la magnitud del efecto de CSNc. Posteriormente, el efecto independiente del antagonista naloxone fue abordado por Pellegrini, Wood, Daniel, y Papini (2005). En una serie de experimentos encontraron que el naloxone administrado durante los ensayos posteriores a la reducción del incentivo prolonga el CSNc, sin afectar la conducta consumatoria de los animales expuestos sólo a la solución de menor magnitud.
Como antagonista, el naloxone no produce actividad neuronal por si mismo, sino que impide que los opioides activen las neuronas que poseen receptores. Por lo tanto, estos efectos del naloxone sugieren que el sistema opioide es normalmente activado por una experiencia de reducción del incentivo (dolor psicológico), tal como ocurre durante el condicionamiento de miedo inducido por descargas eléctricas (dolor físico). Una com-prensión más completa del papel del sistema opioide en el CSNc requiere una evaluación del efecto de drogas más selectivas que afectan sólo una clase de receptores.
DPDPE es un agonista especifico del receptor ä opioide que atenúa el efecto del cambio de recompensa en el ensayo 11 pero carece de efectos detectables en el ensayo 12 (Wood, Da-niel & Papini, 2005). Por su parte, la administración del antagonista selectivo del receptor ä opioide, naltrindol, aumenta la magnitud del CSNc en el ensayo 11, pero no en el 12 (Pellegrini et al., 2005). Estos datos sugieren que el sistema ä opioide se activa predominantemente con la experiencia inicial de reducción del incentivo (ensayo 11), pero no con el condicionamiento aversivo que da lugar a la FS y al conflicto (ensayo 12). Estos efectos pueden derivarse de al menos dos posibilidades teóricas (Wood et al., 2005). Por un lado, es posible que el receptor d opioide regule la intensidad del estado incondicionado aversivo de FP. Por otro lado, es posible que este receptor facilite la adquisición del condicionamiento pavloviano que resulta en el desarrollo de la FS. Si el receptor d opioide facilitara el condicionamiento aversivo, entonces la administración posterior al ensayo 11 debería modular el proceso de consolidación de esta memoria. Datos no publica-dos sugieren que el tratamiento con DPDPE o naltrindol luego de finalizado el ensayo 11 no afecta el curso subsiguiente del CSNc, lo que sugiere que el receptor d opioide no está vinculado al condicionamiento (Daniel, Ortega, & Papini, en preparación).
Otra serie de estudios aún no publicados se basa en la administración del agonista U-50,488H, que selectivamente activa el receptor k opioide (amygda, Norris, Daniel, & Papini, en preparación). La administración de U-50,488H antes del ensayo 11 no tiene efectos detectables sobre el CSNc, pero la administración antes del ensayo 12 tiene efectos bidireccionales que dependen de la dosis administrada. Una dosis baja de U-50,488H (1 mg/kg) reduce el CSNc, mientras que una dosis alta (3 mg/kg) lo prolonga. Es interesante notar que la administración de la dosis alta (pero no la baja) inmediatamente después del ensayo 11 prolonga el CSNc de manera similar a la administración de corticosterona descrita previamente. Estos resultados sugieren que el receptor k opioide esta vinculado a la consolidación de la memoria aversiva de la reducción del incentivo.
Los diferentes resultados de las manipulaciones farmacológicas en los ensayos 11 y 12 del procedimiento de CSNc apoyan la idea original de Amsel (1992) de la existencia de dos mecanismos comportamentales diferentes. Uno de ellos es el un estado emocional aversivo inducido por el cambio sorpresivo del incentivo que él denominó FP. El otro es el desarrollo de una expectativa de ese estado emocional aversivo que él denomino FS. La hipótesis de trabajo basada en los resultados de estos experimentos con opioides puede describirse en estos términos. La experiencia del cambio sorpresivo de incentivo de 32% a 4% de sucrosa en el ensayo 11 activa selectivamente el subsistema ä opioide, cuya función es la de regular la intensidad de la reacción emocional de FP. Una reacción de FP intensa inducida por la reducción del incentivo activa el subsistema ? opioide, cuya función es la de facilitar la formación de una memoria asociativa del evento aversivo. La reactivación de esta memoria durante el ensayo 12, combinada con la privación de alimentos y la disponibilidad de la solución del 4% inducen la eclosión de un conflicto de acercamiento-evitación. Los receptores benzodiazepínicos regulan la expresión de este conflicto, posiblemente afectando el componente de evitación del mismo (Gray & McNaughton, 2000).
COMENTARIOS FINALES
De acuerdo con Albright, Jessell, Kandel y Posner (2000), el problema fundamental para las neurociencias del Siglo XXI es el de encontrar estrategias que combinen de manera exitosa las aproximaciones moleculares y las holísticas. Por ejemplo, para comprender la memoria será necesario construir un puente entre estas dos aproximaciones (Kandel, 2001). Sin embargo, la manera en que tal tipo de colaboración pudiera implementarse no ha sido explícitamente desarrollada, especialmente desde las neurociencias. Dado que el comportamiento es en última instancia el resultado del funcionamiento del diversos sistemas neuronales, se hace cada vez más claro que el papel de la psicología, la ciencia del comportamiento, será el de proveer las herramientas para lograr un modelo integrador. Abram Amsel fue un pionero en la construcción de tal modelo y su propia investigación fue realizada siguiendo estos lineamientos.
REFERENCIAS
Albright, T. D., Jessell, T. M., Kandel E. R. & Posner M. I. (2000). Neural science: A century of progress and the mysteries that remain. Cell, 100, S1–S55. [ Links ]
Amsel, A. (1958). The role of frustrative nonreward in noncontinuos learning. Psychological Bulletin, 55, 102-119. [ Links ]
Amsel, A. (1962). Frustrative nonreward in partial reinforcement and discrimination learning; some recent history and a theoretical extension. Psychological Review, 69, 306-28. [ Links ]
Amsel, A. (1967). Partial reinforcement effects on vigor and persistence: advances in frustration theory derived from a variety of withinsubjects experiments. In K.W. Spence & J. T. Spence (Eds.), The psychology of learning and motivation; advances in research and theory (pp. 51-69). New York: Academic Press. [ Links ]
Amsel, A. (1972). Behavioral habituation, counterconditioning, and a general theory of persistence. In Black A. H. & Prokasy, W. F. (Eds.), Classical conditioning II: Current research and theory (pp. 409-26). New York: Appleton-Century-Crofts. [ Links ]
Amsel, A., & M. Stanton (1980). Ontogeny and phylogeny of paradoxical reward effects. In J. S. Rosenblatt, R. A. Hinde, C. Beer, M. Busnel (Eds.), Advances in the Study of Behavior, Vol. 2, (pp. 227-274). New York: Academic. [ Links ]
Amsel, A. (1989). Behaviorism, neobehaviorism, and cognitivism in learning theory: Historical and contemporary perspectivas. Hillsdale, NJ: Erlbaum. [ Links ]
Amsel, A. (1992). Frustration theory. An analysis of dispositional learning and memory. New York: Cambridge University Press. [ Links ]
Balleine, B.W. & Killcross, S. (2006) Parallel incentive processing: an integrated view of mygdale function. Trends in Neuroscience, 29, 272–279. [ Links ]
Becker, H. C., Jarvis, M. F., Wagner, G. C. & Flaherty, C. F. (1984). Medial and lateral amygdalectomy differentially influences consummatory negative contrast. Physiology and Behavior, 33, 707-12. [ Links ]
Bentosela, M., Ruetti, E., Muzio, R. N., Mustaca, A. E. & Papini, M. R. (2006). Administration of corticosterone after the first downshift trial enhances consummatory successive negative contrast. Behavioral Neuroscience, 120, 371-376. [ Links ]
Davis, M. (2006). Neural Systems Involved in Fear and Anxiety Measured With Fear-Potentiated Startle. American Psychologist, 61, 741-756. [ Links ]
Dudley, R. T. & Papini, M. R. (1995). Pavlovian performance of rats following unexpecved reward omissions. Learning and Motivation, 26, 63-82. [ Links ]
Elliot, M. H. (1928). The effect of change of reward on the maze performance of rats. University of California Publications in Psychology, 4, 19-30. [ Links ]
Fanselow, M.S. & Poulos, A.M. (2005). The neuroscience of mammalian associative learning. Annual Review of Psychology, 56, 207– 234. [ Links ]
Flaherty, C. F. (1996). Incentive relativity. Cambridge, UK: Cambridge University Press. [ Links ]
Flaherty, C. F., Becker, H. C. & Pohorecky, L. (1985). Correlation of corticosterone elevation and negative contrast varies as a function of postshift day. Animal Learning and Behavior, 13, 309-314. [ Links ]
Flaherty, C. F., Clark, S. & Coppotelli, C. (1996). Lack of tolerance to the contrast-reducing actions of chlordiazepoxide with repeated reward reductions. Physiology and Behavior, 60, 645-652. [ Links ]
Flaherty, C. F., Grigson, P. S. & Rowan, G. A. (1986). Chlordiazepoxide and the determinants of contrast. Animal Learning and Behavior, 14, 315-321. [ Links ]
Genn, R. F., Ahn, S., Phillips, A. G. (2004). Attenuated dopamine efflux in the rat nucleus accumbens during successive negative contrast. Behavioral Neuroscience,118, 869-873. [ Links ]
Giles, A. C., Rose, J. K., & Rankin, C. H. (2006). Investigations of learning and memory in Caenorhabditis elegans. International Review of Neurobiology, 69, 37-71. [ Links ]
Gray, J. A. (1987). The Psychology of Fear and Stress. Cambridge, UK: Cambridge University Press. [ Links ]
Grigson, P.S., Spector, A.C. & Norgren, R. (1994). Lesions of the pontine parabrachial nuclei eliminate successive negative contrast effects in rats. Behavioral Neuroscience 108, 714-723. [ Links ]
Hull, C. L. (1931). Goal attraction and directing ideas conceived as habit phenomena. Psychological Review, 38, 487-506. [ Links ]
Hull, C. L. (1943). Principles of Behavior. New York: Appleton-Century-Crofts. [ Links ]
Kandel, E. R. (2001). The molecular biology of memory storage: A dialogue between genes and synapses. Science, 294, 1030-1038. [ Links ]
Liao, R. M. & Chuang, F. J. (2003). Differential effects of diazepam infused into the mygdale and hippocampus on negative contrast. Pharmacology, Biochemistry and Behavior, 74, 953-960. [ Links ]
Lundy Jr., R. F. & Norgren, R. (2004). Gustatory system. In Paxinos, G. (Ed.), The Rat Nervous System, Third edition. San Diego: Academic Press. [ Links ]
Mansour, A., Fox, C. A., Akil, H. & Watson, S. J. (1995). Opioid-receptors mRNA expression in the rat CNS: Anatomical and functional implications. Trends in Neurosciences, 18, 22-29. [ Links ]
Millar, N. E (1944). Experimental studies of conflict. In Hunt , J. (Eds.), Personality and the Behavior disorders (pp. 431-465). Oxford, UK: Ronald Press. [ Links ]
Mitchell, C. & Flaherty, C. F. (1998). Temporal dynamics of corticosterone elevation in successive negative contrast. Physiology and Behavior, 64, 287-292. [ Links ]
Mowrer, O. H. (1960). Learning theory and behavior. New York: Wiley. [ Links ]
Mustaca, A. E. & Papini, M. R. (2005). Consummatory successive negative contrast induces hypoalgesia. International Journal of Comparative Psychology, 18, 333-339. [ Links ]
Nestler, E. J., Hyman, S. E., Malenka, R. C. (2001). Molecular Neuropharmacology: A Foundation for Clinical Neuroscience. New York: McGraw Hill. [ Links ]
Ovemier, J. B. & Lawry, J. A. (1979). Pavlovian conditioning and the mediation of behavior. Psychology of Learning and Motivation, 13, 1-55. [ Links ]
Papini, M. R. (2003). Comparative psychology of surprising nonreward. Brain, Behavior and Evolution, 62, 83-95. [ Links ]
Papini, M. R. (2006). Role of surprising nonreward in associative learning. The Japanese Journal of Animal Psychology, 58, 35-54. [ Links ]
Papini, M. R. & Dudley, R. T. (1997). Consequences of surprising reward omissions. Review of General Psychology, 1, 175-197. [ Links ]
Papini, M. A., Wood, M., Daniel, A. M. & Norris, J. N. (2006). Reward loss as psychological pain. International Journal of Psychology and Psychological Therapy, 6, 189-213. [ Links ]
Pavlov, I. P. (1927). Conditioned reflexes. Oxford, England: Oxford University Press. [ Links ]
Pecoraro, N. & Dallman, M. F. (2005). C-Fos after incentive shifts: expectancy, incredulity, and recovery. Behavioral Neuroscience, 119, 366-387. [ Links ]
Pellegrini S., Wood, M., Daniel A. M. & Papini M. R. (2005). Opioid receptors modulate recovery from consummatory successive negative contrast. Behavioural Brain Research, 164, 239–249. [ Links ]
Pellegrini S., Muzio, R. N., Mustaca, A. E, & Papini M. R. (2004). Sucessive negative contrast after partial reinforcement in the consummatory behavior of rats. Learning and Motivation, 35, 303–321. [ Links ]
Reilly, S., & Trifunovic, R. (1999). Gustatory thalamus lesions eliminate successive negative contrast in the rat. Behavioral Neuroscience, 113, 1242-1248. [ Links ]
Rescorla, R. A.& Wagner, A. R. (1972) A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In Black, A. H. & Prokasy W. F. (eds.), Classical Conditioning: II. Current research and theory (pp 64–99). New York: Appleton-Century-Crofts. [ Links ]
Rowan, G. A. & Flaherty, C. F. (1987). Effect of morphine on negative contrast in consummatory behavior. Psychopharmacology, 93, 51-58. [ Links ]
Sastre, A. & Reilly, S. (2006). Excitotoxic lesions of the gustatory thalamus eliminate consummatory but not instrumental successive negative contrast in rats. Behavioural Brain Research, 170, 34-40. [ Links ]
Skinner, B. F. (1935) Two types of conditioned reflex and a pseudo type. Journal of General Psychology, 12, 66-77. [ Links ]
Stout, S. C., Boughner, R. L. & Papini, M. R. (2003). Reexamining the frustration effect in rats: Aftereffects of surprising reinforcement and nonreinforcement. Learning and Motivation, 34, 437–456. [ Links ]
Tabuchi, E., Yokawa, T., Mallick, H., Inubushi, T., Kondoh, T., Ono, T. & Torii, K. (2002). Spatio-temporal dynamics of brain activated regions during drinking behavior in rats. Brain Research, 951, 270-9. [ Links ]
Thomas, B. L. & Papini, M. R. (2003). Mechanisms of spaced-trial runway extinction in pigeons. Learning and Motivation, 34, 104– 126. [ Links ]
Thorndike, E. L. (1911). Animal Intelligence: Experimental studies. New York: Macmillan. [ Links ]
Travers, J. B., Dinardo, L. A. & Karimnamazi, H. (1997). Motor and premotor. Mechanisms of licking. Neuroscience and Biobehavioral Reviews, 21, 631-647. [ Links ]
Wood, M., Daniel, A. M. & Papini, M. A. (2005). Selective Effects of the Opioid receptor agonist DPDPE on consummatory successive negative contrast. Behavioral Neuroscience, 119, 446–454. [ Links ]
Recepción: Mayo de 2007
Aceptación final:Septiembre de 2007

