










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Latinoamericana de Psicología
versão impressa ISSN 0120-0534
rev.latinoam.psicol. v.44 n.1 Bogotá jan./abr. 2012
Faculdade São Bento, Faculdade Santíssimo Sacramento, Brasil
Thomas A. R. Woelz
Pontifícia Universidade Católica de São Paulo, Brasil
Sigrid S. Glenn
University of North Texas, EE.UU.
The experiments were designed and conducted by the first author in partial fulfillment of requirements for the Master of Science degree in behavior analysis at the University of North Texas. The computer program was developed by the second author in collaboration with the first author. We consider the manuscript has benefitted from the collaboration with the first author. We consider the manuscript has benefitted from the thoughtful and detailed suggestions of the journal reviewer and hereby express our appreciation. Please address correspondence regarding this paper to Ana Barbara Neves at ana.babi.neves@gmail.com
Recibido: Septiembre 20 de 2011, Revisado: Diciembre 2 de 2011, Aceptado: Febrero 4 de 2012
Resumen
Se condujeron dos experimentos en una Universidad en la cual díadas de estudiantes participaron en una simulación de cazadores que debían compartir su presa para mantener la adaptabilidad diádica (grupal). El compartir era una contingencia conductual entrelazada que contribuía a la supervivencia de la díada, conceptualizada como un nomoclón de cacería (Harris, 1964). La simulación incluyó seis temporadas de caza consecutivas en las cuales la variable antecedente de la escasez de presas fue manipulada como variable independiente. Los resultados del primer experimento no mostraron diferencias en la adaptabilidad diádica como función de la escasez de presas. En el segundo experimento, la diferencia entre condiciones de pobreza y riqueza se aumentó. En las condiciones de riqueza, todas las díadas se desempeñaron de forma similar a las del primer experimento. Sin embargo, en las condiciones de sucesiva pobreza, las díadas comenzaron menos adaptadas pero aumentaron su adaptabilidad progresivamente. De este modo, compartir contingencias entrelazadas fue más difícil en condiciones menos significativas de escasez, pero las mismas se hicieron más frecuentes con el tiempo. El experimento se discute a la luz de la visión de Skinner (1981) sobre la evolución cultural, la taxonomía de cosas culturales de Harris (1964) y la formulación de metacontingencias de Glenn (1988, 2004).
Palabras clave: análisis cultural, contingencias conductuales entrelazadas, metacontingencias.
Abstract
Two experiments were conducted in a college where students in dyads participated in a simulation of hunters who were required to share prey in order to maintain dyadic (group) fitness. The sharing was an interlocking behavioral contingency contributing to survival of the dyad, conceptualized as a hunting nomoclone (Harris, 1964). The simulation comprised 6 consecutive hunting seasons in which the antecedent variable of prey scarcity was manipulated as the independent variable. Results of the first experiment did not show a difference in dyadic fitness as a function of prey scarcity. In the second experiment the difference between poor and rich conditions was increased. In rich conditions, all the dyads performed similarly to those in the first experiment. However, in successive poor conditions, dyads started out less fit and became increasingly fit. Thus, sharing IBCs were more difficult to form under significant scarcity, but they became more frequent over time. The experiment is discussed in the context of Skinner's view (1981) about cultures evolution, Harris's (1964) taxonomy of cultural things, and Glenn's (1988, 2004) formulation of metacontingencies.
Keywords: Cultural analysis, experimental analysis, interlocking behavioral contingencies, metacontingencies.
In Selection by Consequences, B. F. Skinner (1981) accounted for human behavior in terms of three kinds of selection. He compared operant conditioning, as a second kind of selection, to natural selection and suggested that a third kind of selection was involved in the evolution of cultures. "A culture evolves when [its] practices...contribute to the success of the group in solving its problems. The effect on the group, and not the reinforcing consequences for individual members, is responsible for the evolution of cultures" (Skinner, 1981, p. 502).
Skinner's 1981 article was reprinted in 1984 as the subject of commentaries by scholars in numerous disciplines. Among the commentators it was the cultural anthropologist Marvin Harris, who began by acknowledging his "fundamental agreement with Skinner's positivism and materialism and [his] own intellectual grounding in reinforcement principles as taught by William Schoenfeld and Fred Keller..." (Harris ,1984, p. 490). Harris then called the "group" in Skinner's statement quoted at the end of the last paragraph "an epistemological lapse (unoperationalized entity)". In his taxonomy of "cultural things", Harris distinguished among different types of phenomena commonly called "groups" and explained that these various kinds of groups play different roles in the analysis of cultures. Of current interest are those "groups" that Harris (1964) designated as nomoclones.
Nomoclones, like all "cultural things", begin with individual human behavior; but they do not end there. Identifying a nomoclone entails identifying specific actors performing at a particular place, who repeatedly behave with respect to one another's behavior and behavioral products in particular ways. An example of a nomoclone given by Harris is the crew of a fishing boat who repeatedly "meet only for performances of a series of idioscenes of the 'same' scene", i.e., fishing episodes (Harris, 1964, p.115). Harris clarifies that the nomoclone is identified both by its particular individual participants and by the repetition of their interrelated performances. We view these repetitions as constituting the interlocking behavioral contingencies (IBCs) that Glenn (1988, 2003, 2004) suggested as the cultural level units undergoing selection by "metacontingencies". Sometimes, Glenn (e.g., 1988; 2003) viewed the product of the IBCs as the selecting agent; at other times, the product was itself part of the unit undergoing selection (e.g. Glenn, 2004; Glenn & Malott, 2004). The fact that metacontingencies can work in either way is suggested.
Vichi, Andery and Glenn (2009) reported an experiment in which metacontingencies worked in the latter way. In their experiment, specific IBCs were required to achieve a group monetary consequence that was contingent on the product of IBCs. In this metacontingency arrangement, the contingency between IBC products and the group consequence was manipulated. Depending on condition, the group consequence was contingent on either equal or unequal distribution of proceeds in the previous round. The distributions (products of the group IBCs) conformed to the requirements of the metacontingency. No participant reported verbalizing the relation between previous distribution of proceeds and subsequent group earnings.
In the present experiments, products of the interlocking behavioral contingencies are viewed as consequences of the IBCs and the metacontingency is held constant while antecedent environments are manipulated as the independent variable. The experiments draw on the method of Ward, Eastman and Ninness (2009), who developed an experimental analog of 2-person nomoclones. They viewed Harris as suggesting "a negative correlation between production efficiency and resource sharing" (Ward et al, p. 60) and they sought to model that relation by manipulating production efficiency in a simulation of hunter/gatherer dyads in which resource sharing was the dependent variable. Participants obtained "resources" on each trial and were required to expend a resource to continue playing. If a player did not have resources to continue playing, that player "went out" and lost all resources previously obtained. This also made it more likely that the other player would go out. There were six experimental conditions beginning with a condition that provided a steady supply of resources to both members of the dyad. Intense scarcity of resources characterized the second condition, and increasingly less scarcity (greater production efficiency) characterized each successive condition. Opposite to the predicted negative correlation between sharing and production efficiency, sharing occurred a total of only four times for all four dyads combined across all six conditions. Those rare occurrences of sharing were in conditions of less scarcity (greater production efficiency).
The researchers reasoned that allowing participants who "went out" to keep resources earned in conditions where they hadn't gone out did not model hunter-gatherer conditions very well. In a second experiment, three more dyads were exposed to the same conditions with one change in procedure: Restarting after going out resulted in loss of all tokens in a player's possession, including those acquired in past and current conditions. One dyad showed a general trend toward increased sharing across Conditions 3-6, thus a clear positive correlation between resource sharing and production efficiency. One dyad had no sharing/group saves, and one dyad had one sharing/ group save in the last condition.
In these experiments, the selection of a hunting nomoclone in which a specific IBC is required to produce dyadic fitness (ability to hunt) was modeled. This reflects the earlier metacontingency formulation in which the product of the IBCs selects its properties and thus differs from Vichi et al. (2009), who modeled the later formulation of metacontingency. These experiments differ also in that the antecedent environment while the relation between IBCs and product is held constant was manipulated. Vichi et al. manipulated the cultural consequence.
The experiments herein are based on the method used in Ward et al. (2009). Their procedures to study the effect of prey scarcity on dyadic fitness when sharing requirements for survival were the same under all conditions and it was adapted during procedures. Besides, the procedures of the Ward et al (2009) study and automated them for computer delivery were simplified.
General Method
Overview
Two experiments were conducted in which subjects participated in dyads functioning as nomoclones. The members of each nomoclone "hunted" independently, each returning with a quantity of rabbits on each trial. Each participant was required to "eat" a rabbit to continue to the next trial. Remaining rabbits could be "stored" for one trial, "traded" for money, or "shared" (given to the other participant). Dyads hunted for six "seasons". Each season was identified as offering either a "rich" environment or a "poor" environment, determined by the average number of rabbits the experimenters allowed the two hunters. Thus, environmental scarcity was manipulated as the independent variable.
The dependent variable was dyadic fitness, compared in Rich and Poor conditions. Dyadic fitness was defined as both participants being able to hunt and it was measured by the proportion of trials in a season that both participants hunted. The program was designed so that maintaining dyadic fitness through each season required both participants to give at least three rabbits to the other participant during the course of the season. That is, interlocking behavioral contingencies were necessary for survival of the nomoclone.
Experiment 1
Method
Participants and Setting
Five females and one male, ages between 18 and 26, participated in three dyads. Three participants were undergraduate students, taking an introductory course in behavior principles, and three were behavior analysis graduate students. One dyad included two female undergraduates, ages 20 and 21; another dyad included two female graduate students, ages 23 and 26; and the remaining dyad included one 18-year-old, male undergraduate and one 26-year-old female graduate student. The experiment took place in a small research room equipped with a table, three chairs and a computer.
Apparatus
The experimental procedure was implemented by a computer program written in Visual Basic.NET and using Microsoft Office Excel. The computer keyboard was altered by labeling the active keys. The altered keyboard is depicted in Figure 1.

Experimental Design
Each dyad participated in one experimental session of 360 trials, divided into six "hunting seasons" of 60 trials each. Poor and rich conditions were quantified by proportion of hunts that each player came home with no rabbits (p = .36 in rich conditions and p = .23 in poor conditions).
For the first dyad participating in the experiment, the conditions for two seasons before changing conditions (A-A-B-B-A-A design) were repeated. Seeing little difference in performance in the Rich and Poor conditions, the design to A-B-A-B-A-B to allow the participants more experience with changing conditions was changed. In all cases we began with a Poor condition.
Procedure
A trial began when either of the two participants pressed the key labeled "Hunt." In each trial, pictures of rabbits and a pair of numbers then appeared at the top of the screen representing how many rabbits each hunter brought home (Figure 2). Each player then was required to distribute his or her resources (by pressing appropriate keys) in one or more of the following ways: eat a rabbit in order to continue hunting; place rabbit(s) in his/her store to be used within the current or next trial; trade rabbit(s) for tablets/coins to be exchanged for money at the end of the experiment; give a rabbit to the other participant.
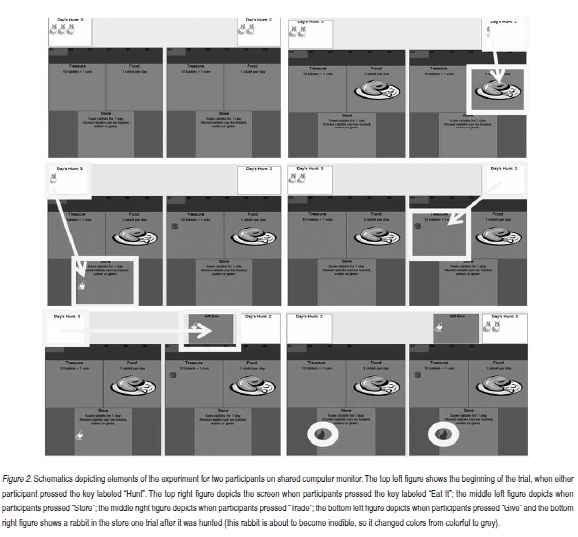
When a participant pressed the "give" button, a gift box containing the gifted rabbit appeared on the receiver side of the screen. Any time after the hunt, participants could move rabbits in their store or gift box. Rabbits could remain in the store for only one more hunt after the trial they were obtained. A stored rabbit turned to grey at the beginning of the trial after it was stored, signaling it was about to become inedible. If participants did not move the grey rabbit during that trial it disappeared when the next trial was initiated.
The money participants accumulated in each season was displayed at the top of the screen in boxes labeled S1 to S6. If a participant had no rabbit to eat, or did not eat one, on any trial, a panel labeled "OUT" covered that participant's side of the screen. When a participant went out, that participant lost all resources he or she had accumulated up to that point in the season, while the remaining player continued hunting. If the second player went out, also losing all resources accumulated for that season, both hunters could return to hunt until the season ended. The program was designed to insure that no hunter could hunt indefinitely without the other.
If a season ended while one participant was out and the other participant was hunting alone, then the participant who continued hunting kept the points he or she had accumulated during that season. Both participants could hunt again in the following season.
The experimental program included two pre-set sequences of 60 pairs of numbers (rabbits obtained from the 60 hunts). One pre-set number sequence was designed to represent "Poor" hunting conditions and the other sequence to represent "Rich" conditions. When the "hunt" key was pressed, each participant received 0, 1, 2, or 3 rabbits. The number obtained was independent of the participants' performance and overall was equalized for participants.
In each season, there were 10 trials in which each participant received only 1 rabbit. During seasons of "Poor" conditions there were 22 trials in which one or the other participant received 0 rabbits at different times, 14 trials in which one received 2 rabbits and 14 in which one received 3 rabbits. In "Rich" conditions participants received 0 rabbits in 14 trials, 2 rabbits in 18 trials, and 3 in the remaining 18 trials.
An Excel program was constructed to program the sequence of rabbits obtained in the experimental conditions. The sequence was designed to insure that in all conditions, each participant was required to give a rabbit to the other at least 3 times in order for both to continue hunting throughout the entire 60-trial season. A situation where one participant had to give a rabbit in order for the other to continue hunting an "opportunity to save" was labeled.
Data Collection
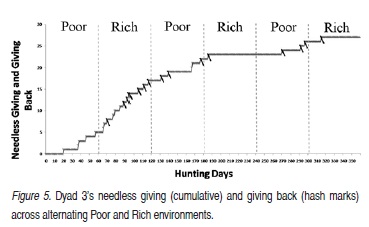
The program was designed to collect data on several variables so that it could be used by several experimenters. The variable of interest in the current experiment was how many cycles in a season both participants were fit to hunt on the next trial. For dyad 3, the "needless giving" and "giving back" responses also became variables of interest, due to particularities of that dyad's interlocked behavioral contingencies. All data were collected using the Visual Basic. NET and graphed using Microsoft Office Excel.
Results
Figure 3 compares dyadic fitness in Rich and Poor conditions for Dyads 1, 2 and 3 when all Rich conditions for each dyad are combined and compared to Poor conditions for that dyad. There is no difference in overall fitness as a function of environmental scarcity, with the possible exception of Dyad 2. Dyad 1 was fit 204 of the 240 days (85%) in four poor seasons and 109 of the 120 (90.8%) hunting days in two rich seasons. Dyads 2 and 3 hunted 180 days when environmental resources were scarce (poor) and 180 days when environmental resources were more abundant (rich.); Dyad 2 was fit for 153 of 180 days (85%) in poor seasons and 180 of 180 days (100%) in rich seasons; Dyad 3 was fit for 168 of 180 days (93.3%) days in both poor and rich seasons.
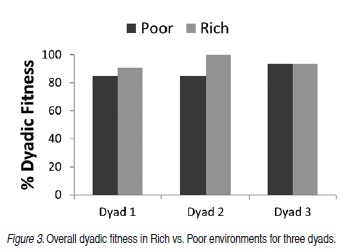
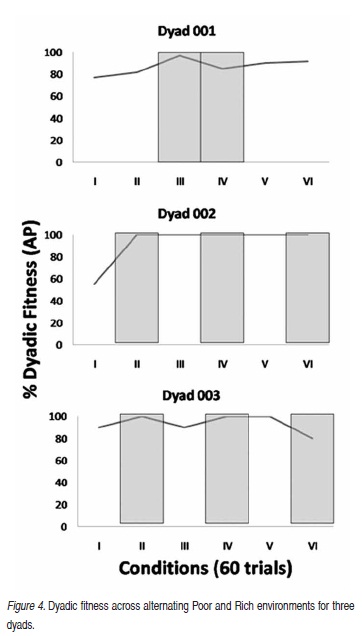
Figure 4 displays the fitness data for the three dyads across alternating Rich and Poor seasons showing trends over time in fitness relative to environmental scarcity. Dyad 1 was fit for 46 of the 60 days of Season I (Poor), 49/60 days of Season II (Poor), 58/60 days of Season III (Rich), 51/60 days of Season IV (Rich). In the last two Poor conditions, Dyad 1 was fit 54 and 55 days respectively. Dyad 2 was fit for 33 days of Season I (Poor) and all 60 days in all subsequent seasons. Dyad 3 was fit for 54 days of Season I (Poor), 60 days in Season II (Rich), 54 days in Season III (Poor), 60 days in Season IV (Rich), 60 days in Season V (Poor), and 48 days in Season VI (Rich).
In Dyad 3, one participant or another sometimes gave a rabbit to the other when the recipient did not need one, and sometimes the recipient of this needless giving immediately returned the unneeded rabbit. In Figure 5, the cumulative record displays "needless giving" in Dyad 3 and the hash marks on the record represent the "giving back" in Dyad 3. The participants needlessly gave four rabbits in Season I (Poor) and no rabbits were returned. In Season II (Rich), participants needlessly gave 12 rabbits and seven of them were returned. In Season III (Poor), participants gave five rabbits and returned three. In Season IV (Rich), a participant gave one rabbit and it was returned. Three rabbits were given in Season V (Poor) and one was returned. One rabbit was given, and returned, in Season VI (Rich).
Discussion
Figure 3 shows that overall dyadic fitness was not very different in Rich and Poor conditions. But when data are displayed over time it is possible to see that, in general, dyadic fitness was high in all Rich conditions, suggesting that sharing was easily achieved. The lower dyadic fitness for all three dyads in the first Poor condition cannot be unequivocally attributed to scarcity because Poor conditions were always experienced first. So the lower dyadic fitness in the first condition for all dyads probably represents the development of the IBC as the individuals in the dyad learned to behave with respect to relevant stimulus conditions. However, the changes in dyadic fitness over time for Dyad 1, and perhaps Dyad 3, suggest that the IBCs representing dyadic performance emerged and strengthened across all Poor conditions.
In the first experiment, the probability of getting 0 rabbits in the Poor conditions was 0.36 and the probability of getting 0 in the Rich conditions was 0.23. Therefore, there was not much difference in scarcity in the Rich and Poor conditions. In the second experiment the difference in the values of Rich and Poor conditions was increased, so that the probability of getting 0 in the Poor conditions was 0.46 and the probability of getting a 0 in the Rich condition was 0.16. In order to allow for that discrepancy, it was necessary to extend the seasons to 100 trials.
Experiment 2
Method
Participants and Setting
Three females and one male, ages between 18 and 45, participated in two dyads. All were undergraduate students, taking an introductory course in Behavior Principles. All were naïve with respect to concept of metacontingencies. One dyad included two female students, ages 20 and 23; and the other dyad included one 22-year-old female student and one 45- year- old male student. The experiment took place in a research room equipped with a table, three chairs and a computer. Stimuli, responses and delivery system were the same as for Experiment 1.
Experimental Design
The level of environmental scarcity was the antecedent condition manipulated. Poor and Rich conditions were quantified as proportion of hunts that each player obtained 0 rabbits (p = .46 in rich conditions and p = .16 in poor conditions). Each dyad worked under two different conditions of environmental scarcity, shown in Table 1. A reversal A-B-A-B-A-B design was used, beginning with a poor condition. Dyadic fitness, measured as in Experiment 1, was compared in Rich and Poor conditions.
Procedure
A sequence of 100 pairs of numbers (rabbits obtained from the 100 hunts) was programmed in Python for Experiment 2. In each season, there were 14 trials in which each participant received only one rabbit. The number of rabbits received on the remaining 86 trials differed for seasons in the "Poor" and "Rich" conditions. During "Poor" seasons there were 36 trials in which participants received 0 rabbits, 25 trials in which they received 2 rabbits and 25 in which they received 3 rabbits. While in "Rich" seasons participants received 0 rabbits in 16 trials, 2 rabbits in 35 trials, and 3 in the remaining 35 trials.
The dyads took part in one l session comprising six "hunting seasons" of 100 trials each. Other elements of the procedure were the same as in Experiment 1.
Results
Figure 6 shows overall dyadic fitness for Dyads 4 and 5, comparing performance for all Rich conditions to performance for all Poor conditions. Dyads 4 and 5 hunted 300 days, when the environment was characterized by scarcity (Poor), and 300 days when the environment was characterized by more abundance (Rich). Dyad 4 was fit for 199 of 300 (66.3%) days in Poor seasons and 262 of the 180 (87.33%) hunting days in Rich seasons. Dyad 5 was fit for 146 of 300 (48.7%) days in Poor seasons and 252 of 300 (84%) of Rich seasons.
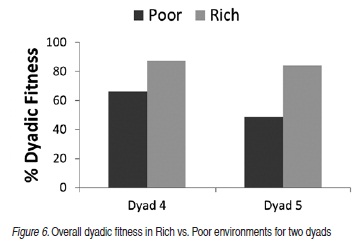
Figure 7 shows season by season dyadic fitness as the independent variable was manipulated across the six seasons. Dyad 4 was fit for 40 days in Season I (Poor), 100 days in Season II (Rich), 65 days in Season III (Poor), 71 days in Season IV (Rich), 94 days in Season V (Poor) and 91 days in Season VI (Rich). Dyad 5 was fit for 20 days in Season I (Poor), 89 days in Season II (Rich), 49 days in Season III (Poor), 100 days in Season IV (Rich), 89 days in Season V (Poor), and 77 days in Season VI (Rich).
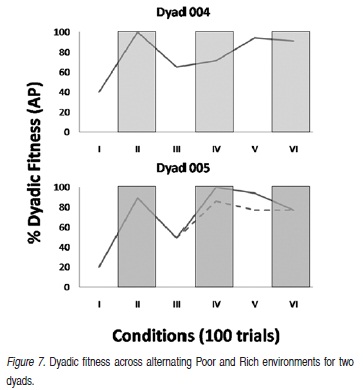
In Poor conditions, dyadic fitness was low in the first season and increased in each of the succeeding Poor seasons (III and V) for Dyads 4 and 5. Dyadic fitness was fairly consistent across Rich conditions. It dropped slightly in the fourth season for Dyad 4, increased again in the next Rich season (VI). For Dyad 5, dyadic fitness increased slightly from the first to the second Rich season (II and IV) and dropped slightly in the last Rich season (VI). In seasons IV and V, rapidly performing participants sometimes pressed the Done key "erroneously" (i.e., when they had rabbits to eat.) The data points connected by the dashed data path in Figure 7 depict results corrected for that error which was likely an artifact of the preparation.
Discussion
The two dyads in the second experiment show an overall difference in dyadic fitness as a function of scarcity in the environment. A comparison of Figures 3 and 6 reveals that an overall quantitative difference in dyadic fitness as a function of scarcity, which was not clear in Experiment 1, is evident in Experiment 2. Further, it can be seen in Figure 7 that, given repeated opportunities for the dyads to function as a unit, IBCs consistent with requirements of the environment gradually form as Poor seasons are experienced. By the end of the experiment, dyadic fitness is as high in Poor as in Rich conditions for both dyads.
The experiments reported here as well as those reported by Ward et al. (2009) did not support the hypothesis that rich environments (or production efficiencies) are less likely to produce IBCs of sharing resources than are poor environments (inefficiencies). All the nomoclones in all the experiments showed as much or more sharing in richer environments. However, in Ward et al. environments became steadily richer over time, so the appearance of sharing at the end of their experiments may have resulted from continuing experience. In the current experiments, discrepancies between sharing in rich and poor conditions could not be attributed to continuing experience.
General Discussion
The experiments reported here derived from the authors' interest in similarities (and differences) in the formulations of Harris, Skinner, and Glenn with respect to cultural analysis.
All three of those authors posit reinforcement contingencies as the phenomena from which cultural processes emerge. However, the concept of metacontingency adds a dimension that neither Harris nor Skinner addressed, which is a cultural level unit (i.e., IBCs) that can undergo selection. It is thought Harris gave us a clue in saying "The cumulative shaping of individual behavior is precisely what cultural evolution is all about. Of course, these behaviors are interrelated and, in conjunction with various environmental and social feedback processes, possess systemic properties that are the logico-empirical basis for the concepts of society, culture, and sociocultural systems" (Harris, 1984, p. 491, emphases added).
In adopting the concept of nomoclone, groups as recurring interlocking behavioral contingencies in which two people produced outcomes (dyadic fitness) that contributed to the origin and maintenance of the IBCs were operationalized. The object of interest in the experiment was on the interlocking behavioral contingencies, not the organisms. This is similar to the focus in operant experiments on behavior rather than on the organism itself. In addition, the focus is on the relation of the IBCs to their antecedent and consequent environments rather than on the topography or controlling variables accounting for individual behavior of participants.
Perhaps it is important to highlight that performance of individual participants at some length and was unable to detect orderly changes in the operant contingencies for individual participants were examined. The research failure to detect regularities may have been due to the complexity of the experimental arrangement. The several ways in which a subject could respond on any given trial and the constantly varying stimulus conditions for each subject across trials may have presented a computational challenge we were not equipped to meet. It is also possible that operant contingencies embedded in IBCs can be highly variable and still meet the requirements of metacontingencies.
Finally, it is remarkable that close examination of Harris's taxonomy of cultural things will yield further opportunities for the extension of a behavior analytic world view to the evolution of cultures. And the study hopes the present experiments offer a preparation that can be used to investigate both antecedents and consequences in metacontingency arrangements. They represent one effort to construct an apparatus that allows measurement and experimental analysis of cultural level units.
References
Glenn, S. S. (1988). Contingencies and metacontingencies: Toward a synthesis of behavior analysis and cultural materialism. The Behavior Analyst, 11, 161-179. [ Links ]
Glenn, S. S. (2003). Operant contingencies and the origin of cultures. In Lattal, K.A. & Chase, P.N. (Eds). Behavior Theory and Philosophy. New York: Kluwer Academic/Plenum. [ Links ]
Glenn, S. S. (2004). Individual behavior, culture, and social change. The Behavior Analyst, 27, 133-151. [ Links ]
Glenn, S. S. & Malott, M. E. (2004). Complexity and selection: Implications for organizational change. Behavior and Social Issues, 13, 89-106. [ Links ]
Harris, M. (1964). The nature of cultural things. New York: Random House. [ Links ]
Harris, M. (1984). Group and individual effects in selection. In Open Peer Commentary. Behavioral and Brain Sciences, 7 (4), 490-491. [ Links ]
Skinner, B. F. (1981). Selection by consequences. Science, 213, 501-504. [ Links ]
Vichi, C., Andery, M.A.P.A. and Glenn, S. S. (2009). A metacontingency experiment: The effects of contingent consequences on patterns of interlocking contingencies of reinforcement. Behavior and Social Issues, 18, 41-57. [ Links ]
Ward, T. A., Eastman, R. L. & Ninness, H.A.C. (2009). An experimental analysis of cultural materialism: The effects of various modes of production on resource sharing. Behavior and Social Issues, 18, 58-80. [ Links ]

