










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Latinoamericana de Psicología
versión impresa ISSN 0120-0534
rev.latinoam.psicol. vol.45 no.2 Bogotá mayo/ago. 2013
Bloqueo y ensombrecimiento en un grupo de vertebrados filogenéticamente antiguo: los anfibios
Blocking and overshadowing in a phylogenetically ancient vertebrate group: amphibians
M. Florencia Daneri, Rubén N. Muzio
Instituto de Biología y Medicina Experimental (IBYME-CONICET) y Universidad de Buenos Aires, Argentina
Correspondencia: Dr. Rubén N. Muzio. Grupo de Aprendizaje y Cognición Comparada, Laboratorio de Biología del Comportamiento, Instituto de Biología y Medicina Experimental (IBYME-CONICET) - Cátedra de Biología del Comportamiento e Instituto de Investigaciones, Facultad de Psicología, Universidad de Buenos Aires (UBA), Argentina. Correo electrónico: rnmuzio@gmail.com Teléfono: 54-11-47832869 Ext. 256
Agradecimientos: Este trabajo se realizó gracias a la financiación del subsidio UBACYT-P052 de la Universidad de Buenos Aires y del subsidio PIP 3196 del CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas) Argentina, dirigidos por Rubén N. Muzio. Los autores agradecen a los dos revisores anónimos por los útiles comentarios realizados sobre este manuscrito.
Recibido: Diciembre de 2012 Revisado: Mayo de 2013 Aceptado: Julio de 2013
Resumen
En este artículo se describe el estudio de los fenómenos de bloqueo y ensombrecimiento en una tarea de aprendizaje espacial en un anfibio, el sapo terrestre Rhinella arenarum. Ambos fenómenos de aprendizaje, ampliamente observados en otras clases de vertebrados, se describen por primera vez en un grupo con un cerebro flogenéticamente antiguo, como es el caso de los anfibios. Específicamente, se observó durante el aprendizaje espacial: (1) bloqueo entre claves visuales asociadas a una meta, y (2) ensombrecimiento de una clave visual lejana por la presencia de una clave cercana. Este hecho permite sentar un precedente para estudiar posteriormente los mecanismos biológicos que rigen el aprendizaje espacial, en búsqueda de patrones funcionales comunes con otras clases de vertebrados, potencialmente presentes en un ancestro común.
Palabras clave: Aprendizaje Espacial - Ensombrecimiento Bloqueo - Anfibios - Análisis Comparado.
Abstract
This article is a study of blocking and overshadowing phenomena in a spatial learning task tested in an amphibian, the common toad Rhinella arenarum. Both phenomena, previously observed in other vertebrates, are described for the first time in a group with a phylogenetically ancient brain - the amphibians. Specifically, it was observed during spatial learning: (1) blocking between visual cues associated to a goal, and (2) overshadowing of a distant visual cue by the presence of a nearby cue. This fact is a precedent for the study of the biological mechanisms that rules spatial learning, thereby looking for common functional patterns with other vertebrates, potentially present in a common ancestor.
Key words: Spatial Learning - Overshadowing - Blocking Amphibians - Comparative Analysis.
Sabemos que un animal puede aprender cuándo responder y/o dónde hacerlo para llegar a la solución de un problema. En este sentido, uno de los problemas clave para la supervivencia es la orientación en el espacio. Animales tan diversos como las abejas y los humanos presentan habilidades para el aprendizaje especial, lo que les permite encontrar una meta oculta que se encuentra a una distancia determinada de una o más claves (Pearce, 2009).
En un trabajo pionero, O'Keefe y Nadel (1978) muestran que las ratas pueden aprender la trayectoria correcta para alcanzar la meta de dos maneras: (i) a través de un verdadero aprendizaje espacial (hipótesis del mapa cognitivo) o (ii) a través de un aprendizaje de guía. En el primer caso, la rata formaría un "mapa cognitivo" del ambiente donde el laberinto está ubicado, y de la ubicación específica del brazo reforzado dentro de ese entorno. Esto implicaría la formación y reajuste de una representación completa del entorno en respuesta a las novedades. O'Keefe y Nadel consideraron este tipo de aprendizaje como no asociativo, muy flexible y que la estructura cerebral responsable es el hipocampo. Por otro lado, el "aprendizaje de guía" implica el acercamiento a una clave o grupo de claves específicas (un color, una forma, olor o textura particular en el brazo reforzado o una clave o configuración de claves visuales detrás del brazo reforzado) (Chamizo, 2002). Este aprendizaje de guía está catalogado como una forma de aprendizaje taxonómico; la otra forma sería la orientación -que Hull (1943) denominó aprendizaje de "respuesta"-. El aprendizaje de guía es un aprendizaje asociativo y no depende del hipocampo; por otro lado, también es menos flexible (Chamizo, 2002). Las teorías del aprendizaje asociativo aplicadas al aprendizaje espacial difieren considerablemente en sus detalles, pero comparten el concepto común de que los estímulos compiten por el control que adquieren sobre el comportamiento (Pearce et al., 2006).
O'Keefe y Nadel enfatizan que las dos estrategias planteadas son formas de aprendizaje diferentes e independientes, cada una de ellas controlada por una estructura cerebral distinta y que sólo la estrategia taxonómica, la de guía, está controlada por las leyes asociativas. Una manera para tratar de esclarecer esta cuestión es considerar si es posible hallar paralelos entre el aprendizaje espacial y otras formas de aprendizaje (Chamizo, 2002).
Para analizar estos paralelismos hemos seleccionado dos de los más llamativos efectos básicos del aprendizaje: el bloqueo y el ensombrecimiento entre claves visuales ambientales. Ambos fenómenos de aprendizaje, ampliamente descriptos en otras clases de animales, son en el presente estudio por primera vez observados en un grupo de vertebrados filogenéticamente antiguo, como es el caso de los anfibios.
Bloqueo y Ensombrecimiento
El fenómeno de bloqueo se observa cuando el establecimiento previo de un elemento de una clave compuesta como señal del reforzador, reduce o "bloquea" lo que se puede aprender acerca del segundo (Kamin, 1969a). Así, el entrenamiento previo con una sola clave bloquea la asociación del refuerzo con otra clave cuando, en una segunda fase del experimento, ambas claves son presentadas simultáneamente señalando la posición del refuerzo. Por otra parte, el término ensombrecimiento se refiere al hecho de que una segunda clave, muy conspicua y saliente, causa que los animales aprendan menos acerca de una primera de lo que lo habrían hecho si se los hubiese entrenado sólo con ella (Kamin, 1969a; Pavlov, 1927). En una situación de entrenamiento donde se ofrecen dos claves, la más conspicua "ensombrecerá" a la otra quitándole poder de asociación con el reforzador.
Un experimento clásico de bloqueo en ratas durante un aprendizaje espacial fue realizado por Roberts y Pearce (1999). Los animales del grupo Bloqueo fueron entrenados en una primera etapa del experimento donde debían encontrar una plataforma sumergida que estaba señalada por una clave adyacente (una barra vertical) en una pileta circular de agua opaca rodeada por cortinas. Una vez que las ratas habían aprendido la tarea, nadando directamente hacia la plataforma cuando eran liberadas en la pileta, se retiraban las cortinas para dar comienzo a la segunda etapa del experimento. El entrenamiento continuó de la misma manera para el grupo Bloqueo, excepto que los animales podían ver ahora las claves provistas por el cuarto experimental. Si el aprendizaje espacial estuviera regido por los principios del aprendizaje asociativo, la barra adyacente a la plataforma tendría que ser considerada una clave confiable, bloqueando el aprendizaje acerca de las claves del cuarto durante la segunda etapa y evitando que éstas ganen control sobre el comportamiento de búsqueda de la plataforma. Para corroborar esta hipótesis, durante los ensayos de prueba los animales fueron colocados en la pileta con las cortinas abiertas, pero sin la barra y sin la plataforma. Si la presencia de la barra no bloqueara el aprendizaje de las claves ambientales, se esperaría que las ratas pasen un tiempo considerable buscando la pileta en el cuadrante donde estaba ubicada originalmente. Si, por el contrario, ocurriera un bloqueo las claves ambientales no les servirían de nada a los animales para hallar la plataforma y la buscarían al azar en cualquier cuadrante de la pileta. Los resultados de los ensayos de prueba se ajustaron a la segunda de estas predicciones (adaptado de Pearce, 2009).
En términos generales, el bloqueo de información en el aprendizaje espacial ha sido estudiado extensamente. Sin embargo, la mayoría de las investigaciones se han centrado en el bloqueo entre dominios espaciales diferentes (como el bloqueo entre claves intra y extra laberinto nombrado previamente). Pocos estudios han examinado el bloqueo dentro de un mismo dominio espacial (Stahlman & Blaisdell, 2009). Por ejemplo, Cheng y Spetch (2001) obtuvieron un fenómeno de bloqueo en el mismo dominio espacial en abejas utilizando un diseño convencional: observaron que el pre-entrenamiento de una clave como indicador de la localización espacial de una solución azucarada bloqueaba el condicionamiento posterior con otras claves agregadas a la situación experimental. En otro estudio, analizando una tarea de forrajeo en ratas, Stahlman y Blaisdell (2009) también reportaron un fenómeno de bloqueo en el mismo dominio espacial utilizando bloques de madera de distintas formas y colores en una arena experimental cuadrada (open field).
Un aspecto de vital importancia a la hora de verificar la presencia de algún fenómeno del aprendizaje es la utilización de grupos controles adecuados. Una discusión detallada de los grupos controles de bloqueo en general (y en no mamíferos en particular) ha sido desarrollada por Acebes y colaboradores (2009). Estos autores se plantean el uso de distintos grupos controles en un diseño de bloqueo utilizando caracoles terrestres (Helix aspersa) en una tarea de aprendizaje asociativo entre un olor y la presentación de alimento. En uno de los experimentos el grupo control no recibió ningún tratamiento durante la primera fase. Éste es uno de los controles más comúnmente utilizados en los experimentos de bloqueo (ver Kamin, 1969a; Sahley et al., 1981), razón por la cual (siendo el presente un primer estudio de este fenómeno en anfibios) luego fue el control seleccionado en nuestro diseño experimental. Esta condición de control permitiría también evaluar la contribución del ensombrecimiento al bloqueo: si la reducción de la respuesta condicionada ante el elemento agregado en la segunda fase se debía simplemente al ensombrecimiento, no habría diferencia entre el grupo experimental y este grupo control. Otros dos tipos de controles que se plantearon Acebes y colaboradores fueron: (i) un grupo control que recibía presentaciones no correlacionadas de un olor y la comida. Este tratamiento ha sido empleado en otras demostraciones de bloqueo (Rescorla, 1971) e implica la equiparación de la experiencia con el estímulo condicionado y el incondicionado durante la primera fase del entrenamiento para los dos grupos (control y experimental), excepto por la contigüidad entre ellos; y (ii) un grupo control que recibía en la primera etapa un condicionamiento con un olor diferente al utilizado en el estímulo compuesto de la segunda etapa. Este tratamiento tiene como objetivo evitar diferencias en el condicionamiento de contexto entre los dos grupos. El uso de estos dos últimos grupos controles será utilizado en próximos diseños experimentales en anfibios.
Finalmente, otro aspecto importante a considerar en el marco del aprendizaje espacial es la influencia de las claves visuales locales en la expresión de estos fenómenos. Chamizo y colaboradores (1985) verificaron por primera vez la existencia de los fenómenos de bloqueo y ensombrecimiento en el uso de claves visuales locales. Estos trabajos permitieron luego ahondar en el análisis de los mismos, encontrándose que las claves espaciales que rodean a un laberinto de Morris pueden afectar al aprendizaje de esta tarea de modo similar a los estímulos condicionados en el condicionamiento clásico o a los estímulos discriminativos en el condicionamiento instrumental (Rodrigo et al., 1997; Sanchez-Moreno et.al., 1999).
Cheng y Spetch (1998) realizaron una revisión general de los posibles mecanismos de uso de claves visuales en el aprendizaje espacial en aves y mamíferos. Allí, describen tres de estos potenciales mecanismos utilizados para encontrar una meta que está en un lugar fijo con respecto a un grupo de puntos de referencia: (1) el uso de las propiedades métricas de la distancia y la dirección (por ejemplo, mediante la observación de errores sistemáticos en una arena de geometría rectangular -Cheng, 1986- o la preferencia observada por los puntos de referencia más cercanos -Cheng, 1989), (2) los principios del uso de claves múltiples (como los usados para definir la ubicación de una meta a través de dos puntos de referencia -Collet et al., 1986); y (3) las distintas maneras en que son usados en conjunto varios puntos de referencia (por ejemplo, mediante el uso de configuraciones y elementos en un arreglo espacial particular -Spetch et al., 1996).
En el caso específico del aprendizaje espacial en anfibios, tenemos un interés inicial en los efectos de la proximidad absoluta entre un punto de referencia y una meta, un fenómeno que en mamíferos fue observado, por ejemplo, en ratas (Chamizo & Rodrigo, 2004) y en humanos (Artigas et al., 2005). En este último trabajo se realizó un experimento en el cual los sujetos debían realizar una tarea virtual de laberinto de Morris. Así, se entrenaron diferentes grupos de sujetos para encontrar una plataforma oculta ubicada en una posición específica (distinta para cada grupo) en relación con uno o dos objetos. Los resultados confirmaron en todos los casos que los puntos de referencia más cercanos adquirían un mejor control respecto de los más distantes (Artigas et al., 2005).
Competencia entre estímulos en el bloqueo y el ensombrecimiento
El ensombrecimiento es un ejemplo de un fenómeno consistente con el concepto de competencia entre estímulos por el control de la conducta. Si dos estímulos son simultáneamente apareados con un estímulo incondicionado en forma repetida, se observa que la respuesta ante la presentación de uno de ellos en forma aislada será menor que si éste hubiese sido apareado de manera individual con el estímulo incondicionado (Pearce et al., 2006). Los efectos del ensombrecimiento pueden ser aumentados mediante un entrenamiento previo adicional con sólo uno de los estímulos antes del condicionamiento con el estímulo compuesto, generando así el fenómeno de bloqueo (Pearce et al., 2006).
Estos fenómenos del aprendizaje parecen estar regidos por un principio de corrección de errores, lo que resulta en una competencia entre estímulos por la fuerza asociativa que puedan adquirir (Rescorla & Wagner, 1972). Por ejemplo, si dos estímulos de igual saliencia señalan un mismo reforzador, cada uno de ellos ganará la mitad de la fuerza asociativa que podrían adquirir si fuesen apareados individualmente con ese mismo reforzador. En esta instancia, la presencia de un estímulo se dice que "ensombrece" al otro. También es posible imaginar una situación donde la presentación de dos estímulos, A y B, señalando juntos un evento particular, es precedida por un entrenamiento en el cual uno de los estímulos (A) por sí solo se aparea con el mismo evento.
Como resultado de este procedimiento, A ganará toda la fuerza asociativa durante la primera etapa, siendo B entonces incapaz de entrar en una asociación con el evento reforzador cuando es posteriormente presentado. Se dice entonces que la presencia de A "bloquea" el aprendizaje acerca de B. En un nivel más general, la competencia por la fuerza asociativa resulta en estímulos que son mejores predictores de un reforzador, los cuales ganan más fuerza asociativa y, por lo tanto, más control sobre el comportamiento que estímulos que son predictores relativamente pobres del reforzador (Pearce, 2009).
Los fenómenos de ensombrecimiento y bloqueo se han observado en peces (Tennant & Bitterman, 1975), aves (Mackintosh & Honig, 1970) y mamíferos (e.g., Kamin, 1969a, 1969b, Chamizo, 2002). Estos fenómenos han sido revelados con una gran diversidad de estímulos en una extensa variedad de procedimientos. Parecería entonces, por razones empíricas y teóricas, que la competencia entre claves por el control del comportamiento es una característica fundamental y persistente del aprendizaje en animales (Pearce et al., 2006). Por lo tanto, la selección de estímulos relevantes (junto a la economía conductual y los procesos atencionales básicos, entre otros) serían características presentes en el bloqueo y el ensombrecimiento que contribuirían a otorgarles relevancia biológica y una posible razón de esta amplia selección evolutiva. A pesar de esta cuestión crucial, explorada en una amplia variedad de clases animales, hasta el presente trabajo no existían datos que puedan confirmar la presencia de estos fenómenos en anfibios.
Estudios en anfibios
Los anfibios son un excelente modelo para el estudio de distintos fenómenos de aprendizaje (Daneri, et al., 2011; Muzio, 1999, 2012). Para trabajar con anfibios es necesario generar un paradigma experimental adecuado que se ajuste a la biología particular de la especie seleccionada y, en este aspecto, su dependencia del agua es un factor clave. Así, hemos desarrollado un modelo experimental exitoso en el sapo Rhinella arenarum utilizando agua como reforzador después de una deshidratación moderada como estímulo para motivar la búsqueda de agua en estos animales (Muzio, 2012).
Los anfibios poseen una gran importancia a nivel evolutivo ya que representan el estado de transición entre el modo de vida acuático y terrestre entre los vertebrados. Algunas especies conservan el hábitat acuático ancestral durante todos los estadios de sus vidas, pero la mayoría evolucionó hacia un ciclo de vida complejo con huevos y larvas acuáticas y juveniles y adultos terrestres. Este cambio de hábitat obliga a las especies a movimientos a través de largas distancias tales como la dispersión post-metamórfica desde los sitos de nacimiento y las migraciones entre distintas áreas en busca de charcas para reproducirse, lo que demanda habilidades para el desplazamiento orientado en el espacio. Por otra parte, comparando el telencéfalo de los anfibios con los de otros tetrápodos presenta anatómicamente un nivel de organización primitivo, en el sentido de no derivado (Muzio, 1999, 2012).
De esta manera, el análisis de la presencia de los fenómenos de bloqueo y ensombrecimiento en anfibios nos permitirá estudiar luego los mecanismos biológicos que rigen estos aprendizajes, en búsqueda de patrones funcionales comunes con otras clases de vertebrados, potencialmente presentes en un ancestro común.
Uso de claves visuales en anfibios
La habilidad de orientarse en un campo abierto (open field) usando claves visuales locales ya ha sido probada con anterioridad en el sapo Rhinella arenarum. Daneri, Casanave y Muzio (2013) entrenaron sapos para que encuentren un contenedor con agua en presencia de claves visuales intralaberinto. Animales parcialmente deshidratados debían recorrer un campo abierto y seleccionar cuál de cuatro contenedores posibles poseía agua accesible para rehidratarse. En el primer experimento, se observó que los animales eran capaces de orientarse en el espacio utilizando dos claves visuales cercanas al contenedor reforzado (localizadas en la pared interna del campo abierto). En el segundo experimento, se utilizó sólo una clave localizada a distintas distancias del contenedor reforzado y se observó que el aprendizaje estuvo afectado por la proximidad a la meta (cuanto más cercana la clave al reforzador, más rápido aprendían los animales). El último experimento reveló que la posición relativa en que está ubicada una clave, le otorga a la misma un valor predictivo particular. En este caso, los animales prefirieron utilizar como referencia las claves más cercanas al reforzador para orientarse y llegar a la meta. Estos resultados previos demostraron que los sapos pueden aprender a orientarse en el espacio usando claves visuales. Además, los datos mostraron que el efecto de proximidad espacial entre una clave y el reforzador, un fenómeno del aprendizaje observado previamente en otros grupos de animales como mamíferos (Chamizo & Rodrigo, 2004, Chamizo et al., 2006; Milgram et al., 1999) e invertebrados (Cheng et al., 1987), también está presente en anfibios.
Los siguientes experimentos de bloqueo y ensombrecimiento constituyen los primeros resultados acerca de la existencia de estos dos fenómenos en el aprendizaje espacial en anfibios usando claves visuales.
Experimento 1: bloqueo entre claves visuales
Método
Sujetos. Se utilizaron 10 sapos machos adultos de la especie Rhinella arenarum provenientes de la provincia de Buenos Aires. Los valores de peso estándar (peso del animal totalmente hidratado con sus vejigas vacías; Ruibal, 1962) variaron entre 65 y 115 g. Los animales se distribuyeron azarosamente en dos grupos, no registrándose diferencias significativas entre los pesos de los mismos (F (1,8)= 2.22, p = 0.17).
Dispositivo Experimental. Se utilizó un campo abierto, arena u open field circular de 86 cm de diámetro y paredes blancas de 100 cm de altura. Se ubicó en el centro del cuarto de experimentación, iluminado uniformemente desde arriba. El dispositivo se encontraba rodeado con cortinas blancas para evitar la visualización de las claves externas y ubicado sobre una mesa giratoria, lo que permitía rotarlo entre ensayos para evitar que alguna marca potencial en la pared interna del laberinto pudiera ser usada como guía. La observación de los animales se realizó desde detrás de la cortina a través de un pequeño visor. Durante los ensayos se ubicaron cuatro piletas idénticas de acrílico color verde (13 x 10 x 3 cm, largo x ancho x altura) distribuidas en cruz contra las paredes laterales, donde sólo una permitía acceso al refuerzo (agua deionizada). Las otras tres piletas también poseían agua, pero la misma estaba fuera del alcance del animal (una rejilla evitaba el contacto del sujeto con el líquido dentro del contenedor). Este procedimiento posee la finalidad de evitar que los animales se guíen hacia la pileta correcta utilizando un gradiente de humedad. La ubicación de la pileta recompensada en las cuatro posiciones se determinó de manera contrabalanceada y pseudoazarosa.
Procedimiento previo general. Durante las dos semanas anteriores al inicio del experimento, los animales fueron habituados a las condiciones de laboratorio y se los alimentó con alimento balanceado para ranas. Durante la primera semana, además, se le agregó a la comida un desparasitante y un antibiótico de acuerdo con un protocolo adecuado para la especie.
El día previo a comenzar con el experimento se determinó el peso estándar de los animales. Los sapos comenzaron los ensayos de entrenamiento y pre-entrenamiento al 80 % de su peso estándar. Para ello, los animales fueron sometidos diariamente a un proceso de deshidratación en jaulas, sin acceso a agua, a fin de aumentar la motivación de los individuos para su búsqueda. Este protocolo de deshidratación es muy utilizado en anfibios y ha demostrado ser no demasiado estresante, e incluso motivante (Brattstrom, 1990; Muzio et al., 1992). Antes y después de cada sesión se registró el peso del animal, calculándose así la variación de peso. Los animales fueron mantenidos en contenedores plásticos individuales sin acceso a agua durante todo el período experimental.
Procedimiento Experimental. Los 10 animales se distribuyeron azarosamente en dos grupos:
Bloqueo (n=5): Para los animales de este grupo, en una primera etapa de entrenamiento la pileta reforzada se encontraba señalada por una clave visual (10 x 10 cm, realizada en goma eva) pegada en la pared blanca de la arena a 10 cm de la pileta (para 3 animales con un rombo rojo a la izquierda, y para 2 con un cuadrado rayado horizontal multicolor a la derecha). Luego, en una segunda etapa, se agregó otra clave visual (la clave a bloquear: rombo o cuadrado, según la clave que recibieron en la primera etapa).
No Bloqueo (n=5): En este caso la pileta reforzada siempre estaba señalada por dos claves visuales ubicadas a 10 cm de la pileta (rombo rojo y cuadrado rayado). Los animales de este grupo sólo tuvieron una etapa de entrenamiento.
Para establecer los tiempos de entrenamiento se estableció como criterio de aprendizaje un mínimo del 50% de respuestas correctas en 3 sesiones consecutivas (5 de 9 ensayos correctos para cada animal).
Los animales de los dos grupos tuvieron 2 sesiones de pre-entrenamiento (1 ensayo cada sesión). A continuación, los animales del grupo Bloqueo tuvieron sesiones de entrenamiento divididas en tres etapas: 1 clave, 2 claves y pruebas (en total, experimentaron 40 sesiones de entrenamiento). En la primera etapa se entrenó a los animales con una sola clave visual hasta criterio de aprendizaje (que todos los animales llegaran a criterio llevó 18 sesiones). Luego, en la segunda etapa de entrenamiento se agregó también la otra clave. Esta etapa se extendió durante la cantidad de sesiones necesarias hasta que el grupo No Bloqueo alcanzara el criterio de aprendizaje (esto llevó 16 sesiones más). Finalmente, se procedió con la etapa de pruebas. Los animales del grupo No Bloqueo se entrenaron a partir de la segunda etapa con las dos claves visuales durante las 16 sesiones que fueron necesarias para alcanzar criterio de aprendizaje; luego también comenzaron las pruebas. La etapa de pruebas consistió de 6 sesiones. Durante las mismas los animales recibieron 4 ensayos por sesión: 3 ensayos de entrenamiento y 1 ensayo de prueba (en segundo o tercer lugar). Los ensayos de prueba fueron incorporados para determinar el valor de predicción de las distintas claves visuales para los animales de ambos grupos. Estos ensayos se dispusieron pseudoazarosamente entre los ensayos de entrenamiento y se retiró el reforzador. Se plantearon 3 tipos de ensayos de prueba: (i) con las dos claves, (ii) sólo con el rombo, y (iii) sólo con el panel rayado (para el grupo Bloqueo, con la primera clave presentada: "clave bloqueadora", o con la segunda clave presentada: "clave bloqueada"). Se dieron dos presentaciones de cada tipo de ensayo (una vez en la segunda posición y la otra en la tercera).
Pre-Entrenamiento. Todos los sujetos fueron sometidos a dos días de pre-entrenamiento con el fin de habituarlos a las condiciones experimentales. Durante las sesiones de pre-entrenamiento los animales podían desplazarse libremente dentro del dispositivo experimental, en el cual se hallaban las cuatro piletas reforzadas con agua. La duración de cada sesión de pre-entrenamiento fue de 5 minutos.
Entrenamiento. Al día siguiente se inició la fase de entrenamiento, que constó de sesiones diarias de 3 ensayos cada una. Cada ensayo se iniciaba extrayendo al animal de su contenedor individual, apoyándolo en un lienzo para eliminar cualquier posible exceso de humedad, y colocándolo suavemente en el centro de la arena debajo de un contenedor plástico cuadrado translúcido invertido (medidas 12cm lado, 8cm alto), siempre en la misma posición (con el rostro hacia el norte). Los animales no podían ver a través de las paredes del contenedor. Se los mantuvo allí por 30 segundos para que se recuperaran de los efectos de la manipulación. Luego se retiraba el contenedor, dejando al animal libre para comenzar a moverse por el campo abierto. Cabe aclarar que dentro del contenedor tenían libertad de movimiento, por lo que el inicio de cada ensayo de entrenamiento se daba con el animal orientado de acuerdo a sus desplazamientos previos dentro del recipiente (siendo de manera azarosa y no observándose ningún sesgo particular). Al comenzar el ensayo se observaba el comportamiento del sujeto en el dispositivo y se registraba el Tipo de Respuesta (Correcta, si accedía a la pileta reforzada; Incorrecta, en caso contrario). Una vez que alcanzaba alguna de las piletas, se le proporcionaba al animal un tiempo de reforzamiento de 2 minutos, dándose luego por concluido el ensayo. El criterio utilizado para determinar la selección de una de las piletas fue que el animal introdujera alguno de sus miembros o la cabeza en ella. Una vez que el sapo accedía a la pileta reforzada era común observar movimientos de frotación de su parche pélvico contra el agua (zona ventral altamente vascularizada y permeable al agua), conducta que está asociada a la rehidratación del animal (Loza Coll & Muzio, 2013).
El movimiento de los animales en el campo abierto durante el ensayo fue grabado en video; esto permitió observar a posteriori con mayor detalle la trayectoria hasta la elección de una de las piletas. El intervalo entre ensayos fue de 2 minutos (donde el animal permanecía en su contenedor individual). Una vez finalizada la sesión (3 ensayos) se retiraba al animal del laberinto, se lo pesaba y se lo devolvía a su recipiente hasta el día siguiente. En caso de ensayo erróneo, se dejaba al animal un minuto en contacto con la pileta no reforzada (o hasta que bajara); luego de ese período se procedía a realizar un ensayo guiado. En este ensayo guiado se estimulaba al animal a caminar hasta acceder a la pileta reforzada (el experimentador lo guiaba suavemente con la mano), donde permanecía 2 minutos. Generalmente, los animales comenzaban a explorar las otras piletas sin necesidad de guiarlos.
Resultados
La Figura 1 muestra el porcentaje de respuestas correctas por sesión de entrenamiento para cada grupo. Un ANOVA de un factor (Sesión) para el grupo Bloqueo durante la primera etapa del entrenamiento muestra diferencias significativas (F (17,74) = 2.05, p = 0.018), revelando la adquisición de la respuesta. Para la segunda etapa del entrenamiento, un ANOVA mixto de dos factores (Grupo x Sesión) con medidas repetidas indica una interacción significativa entre los factores (F (15,120) = 1.91, p = 0.027), siendo también significativa las diferencias para Grupo (F (1,8) = 5.35, p = 0.049), pero no para Sesión (F (15,120) = 0.85, p = 0.611). Al analizar el desempeño individual de cada grupo se observan diferencias significativas a lo largo de las sesiones para el grupo No Bloqueo (F (15,64) = 1.92, p = 0.037), en pleno proceso de adquisición, que no para el grupo Bloqueo (F (15,64) = 0.41, p = 0.969), con un desempeño ya estabilizado en el tiempo.
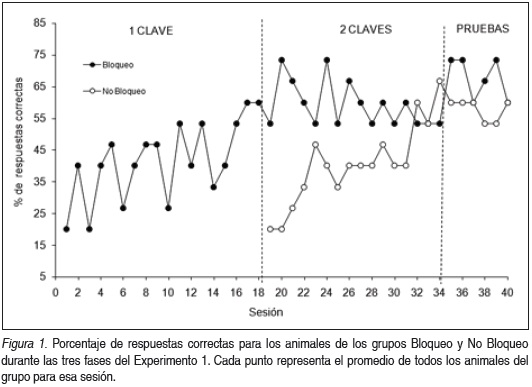
Si analizamos las últimas sesiones de la segunda etapa del entrenamiento, un ANOVA de medidas repetidas señala que no existen diferencias significativas entre los grupos ni en el tiempo durante las 3 últimas sesiones antes de comenzar con los ensayos de prueba (para los factores Grupo, Sesión e Interacción todos los ps > 0.1). Tampoco se vieron diferencias en el desempeño de los grupos durante estos ensayos (para los factores Grupo, Sesión e Interacción todos los ps > 0.1).
El análisis mediante un ANOVA de los ensayos de prueba reveló diferencias significativas entre las distintas condiciones. En la Figura 2 podemos observar que los animales del grupo Bloqueo presentaron un desempeño significativamente menor ante la presentación de la segunda clave (clave bloqueada) (F (2,12) = 4.30, p = 0.038; Test LSD a posteriori, 2 Claves vs 1ra Clave: p = 0.355, 2 Claves vs 2da Clave: p = 0.013, 1ra Clave vs 2da Clave: p = 0.078). No se observaron diferencias significativas en las distintas condiciones de prueba para los animales del Grupo No Bloqueo (ps > 0.868).
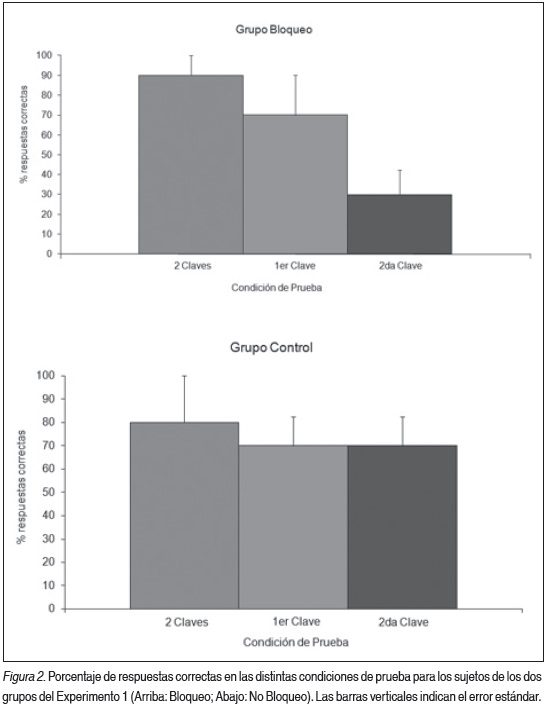
Si comparamos el uso de la segunda clave entre los dos grupos podemos observar diferencias significativas (F (1,8) = 5.33, p = 0.049), lo que señalaría el distinto valor predictivo que adquirió esta clave para los animales de cada grupo.
Discusión
En los animales del grupo Bloqueo se observó que el entrenamiento previo con una sola de las claves bloquea la asociación del refuerzo con la otra clave cuando, en una segunda fase del experimento, ambas claves son presentadas simultáneamente señalando la posición del refuerzo. Este fenómeno no se observó en los animales del grupo No Bloqueo (sin exposición previa), donde las dos claves de presentación simultánea fueron asociadas exitosamente con el reforzador. Este resultado demuestra que el bloqueo entre claves visuales durante el aprendizaje espacial es un fenómeno generalizado, presente en una amplia variedad de vertebrados, que incluiría también a los anfibios.
Una descripción heurística del bloqueo sería que los sujetos no desperdician recursos procesando estímulos redundantes, sólo aprenden nuevas asociaciones cuando es necesario. Este fenómeno, ampliamente estudiado, es relevante para la comprensión de las bases psicológicas del aprendizaje espacial, porque debería producirse el bloqueo si esta forma de aprendizaje cumple con los principios bien establecidos del condicionamiento asociativo (Biegler & Morris, 1999).
Experimento 2: Ensombrecimiento entre claves visuales
Método
Sujetos. Se utilizaron 8 sapos machos adultos de la especie Rhinella arenarum provenientes de la Provincia de Buenos Aires. Los valores de peso estándar variaron entre 103 y 120 g. Los animales se distribuyeron azarosamente en dos grupos, no registrándose diferencias significativas entre los pesos de los mismos (F (1,6) = 1.38, p = 0.283).
Procedimiento Experimental. Los animales se distribuyeron azarosamente en dos grupos:
Ensombrecimiento (n=4): Para los animales de este grupo la pileta reforzada se encontraba señalada por dos claves visuales (un rombo rojo y un panel rayado horizontal multicolor) pegadas en la pared blanca de la arena a 10 cm de altura: la primera, ubicada a 10 cm a la derecha de la pileta reforzada (clave cercana), y la segunda, colocada en la pared que estaba entre la pileta opuesta y la contigua izquierda (clave lejana). Se entrenaron 2 sujetos con el rombo cerca y el panel rayado lejos, y 2 sujetos a la inversa.
Control (n=4): En este caso la pileta reforzada estaba señalada por sólo una clave visual (para la mitad de los animales el panel rayado y para la otra mitad el rombo) ubicada en la pared que estaba entre la pileta opuesta a la pileta reforzada y la contigua izquierda (clave lejana).
Los animales de los dos grupos tuvieron 2 sesiones de pre-entrenamiento (1 ensayo cada sesión), 24 sesiones de entrenamiento y 8 sesiones de prueba. Las sesiones de entrenamiento se dieron hasta que ambos grupos habían alcanzado el criterio de aprendizaje. Los ensayos de prueba fueron 8 para el grupo Ensombrecimiento y 4 para el grupo Control. Durante las sesiones de prueba los animales recibieron 4 ensayos por sesión: 3 ensayos de entrenamiento y 1 ensayo de prueba (en segundo o tercer lugar). Los ensayos de prueba fueron incorporados para determinar el valor de predicción de las distintas claves visuales para los animales de ambos grupos. Estos ensayos se dispusieron pseudoazarosamente entre los ensayos de entrenamiento y se retiró el reforzador. Se plantearon 4 tipos de ensayos de prueba: (i) con las dos claves, (ii) sólo con la clave lejana, (iii) sólo con la clave cercana, y (iv) sin claves. Se dieron dos presentaciones de cada tipo de ensayo (una vez en la segunda posición y la otra en la tercera), sumando 8 ensayos de prueba totales. Los animales del grupo Control sólo tuvieron dos condiciones de prueba (con la clave lejana y sin clave), con un total de 4 ensayos de prueba.
Dispositivo Experimental, Pre-Entrenamiento y Entrenamiento. Se utilizó el mismo dispositivo experimental y el mismo procedimiento que en el Experimento 1.
Resultados
La Figura 3 presenta el porcentaje de respuestas correctas por sesión de entrenamiento para cada grupo. Un ANOVA de medidas repetidas durante la fase de adquisición revela que ambos grupos mejoran significativamente su desempeño a lo largo de las Sesiones (F (23,138) = 2.59, p = 0.0003), no mostrando diferencias significativas en el factor Grupo (F (1,6) = 2.76, p= 0.14), ni en la interacción (F (23,138) = 0.73, p = 0.807). Al analizar las últimas 3 sesiones antes de comenzar con los ensayos de prueba, no existen diferencias significativas entre los grupos ni en el tiempo (para los factores Grupo, Sesión e Interacción, todos los ps > 0.05), lo que señala una adquisición estabilizada para ambos grupos. Tampoco se observaron diferencias significativas durante los ensayos de prueba (para los factores Grupo, Sesión e Interacción, todos los ps > 0.05).
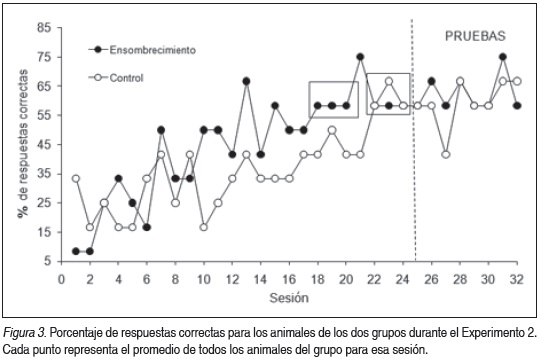
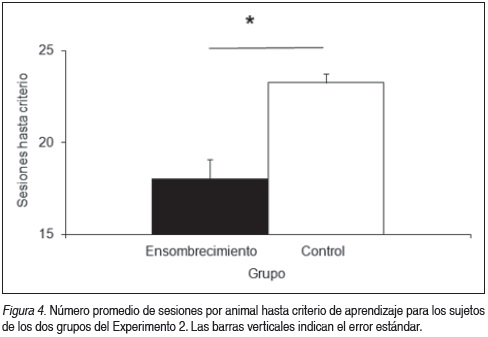
Para ilustrar las diferencias en el tiempo de adquisición de los dos grupos se tomó como variable el tiempo hasta llegar a criterio de aprendizaje, graficado en la Figura 4. Un ANOVA mostró que existen diferencias significativas entre grupos (F (1,6) = 19.74, p < 0.005). El grupo Ensombrecimiento necesitó menos sesiones hasta adquirir el aprendizaje. Esto probablemente se deba a la presencia de una clave cercana al reforzador.
La Figura 5 muestra los resultados de los ensayos de prueba. Un ANOVA comparando el desempeño de ambos grupos en cada situación de prueba revela que los animales del grupo Ensombrecimiento se desempeñaron significativamente peor en la condición de prueba en la cual se presentaba únicamente la clave visual lejana, comportándose de manera similar a la condición sin clave (F (3,12) = 5.66, p < 0.05; Test LSD a posteriori, Clave lejana vs Clave cercana: p < 0.01, Clave lejana vs dos claves: p < 0.05, Clave lejana vs sin clave: p = 1). Esto sugiere que la clave visual lejana en este grupo fue ensombrecida por la presencia de una clave visual cercana al contenedor y se transformó en una clave sin valor predictivo. Esto no se observó en el grupo Control, donde la clave lejana fue asociada exitosamente con la ubicación del refuerzo (F (1,6) = 8, p < 0.05). Si comparamos el uso de la clave lejana entre los dos grupos podemos observar diferencias significativas (F (1,6) = 8.00, p = 0.030), lo que indicaría la diferencia en el valor predictivo que adquirió esta clave para los animales de cada grupo. Por otra parte, tal como era esperado, no se observaron diferencias en el desempeño de los grupos en la condición sin clave (F (1,6) = 0, p = 1).
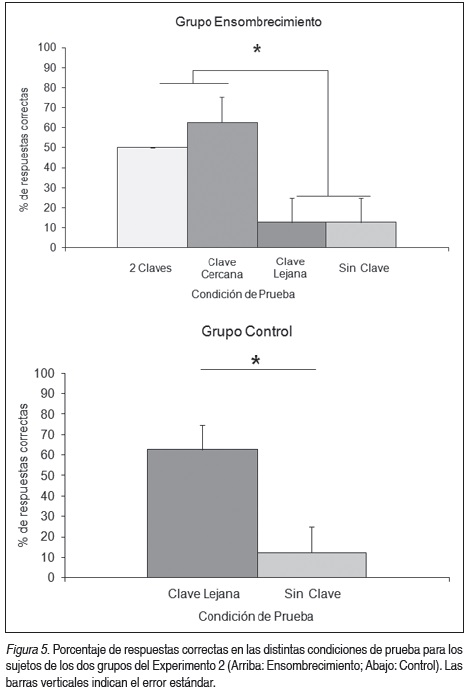
Discusión
Los resultados obtenidos en este experimento sugieren que la clave visual localizada lejos de la pileta con agua fue ensombrecida por la presencia de la clave cercana en los animales del grupo Ensombrecimiento (dicha clave no adquirió ningún valor predictivo). Este fenómeno no se observó en los animales del grupo Control, donde la clave fue estrechamente asociada con la presencia del reforzador y pudo ser utilizada para predecir su posición. Estos resultados sugieren que el fenómeno de ensombrecimiento estaría presente en anfibios.
Discusión General
En general, la evidencia indica que la mayoría de los fenómenos de aprendizaje espacial se pueden explicar mediante los principios del aprendizaje asociativo. Así, los fenómenos de bloqueo y ensombrecimiento observables en el aprendizaje espacial podrían ser explicados por los efectos de la competencia entre claves. En particular, estos resultados sugieren que el aprendizaje espacial estaría regido por una regla de corrección de errores, lo que genera que la clave que es la predictora más confiable de la localización de una meta gane más control sobre el comportamiento del animal (Pearce, 2009). Por ejemplo, la mayoría de las teorías asociativas del aprendizaje (Mackintosh, 1975; Pearce & Hall, 1980; Rescorla & Wagner, 1972) predicen el fenómeno de ensombrecimiento, ya que asumen que los componentes de una clave compuesta compiten entre sí por una cantidad determinada de fuerza asociativa. Así, como hemos visto, un buen predictor ganará esta competencia en detrimento de uno más pobre. Pero al mismo tiempo, el ensombrecimiento podría ser susceptible a otras interpretaciones alternativas. Por ejemplo, este fenómeno podría ser explicado mediante un mecanismo de decremento de la generalización: un animal entrenado con un compuesto AB y testeado sólo con B experimenta un gran cambio desde la etapa de entrenamiento a la etapa de prueba, comparado con otro animal entrenado y testeado sólo con B (Chamizo et al., 2012). Para corroborar si éste es el caso para los resultados obtenidos en anfibios, debemos complementar esta información con futuros experimentos en los cuales se incluyan (entre otros elementos) varios lugares de partida dentro del campo abierto. En particular, en el diseño implementado en nuestro estudio, una clave visual (clave cercana) provee claramente una mejor predicción de la pileta reforzada que otra (clave lejana), lo cual predice una competencia asociativa en la que el mejor predictor ensombrecerá al peor. Sin embargo, si distintas claves visuales son distribuidas equidistantes alrededor del perímetro del campo abierto y los animales son libres de aproximarse a la meta desde cualquier dirección (en lugar de partir siempre desde el centro de la arena), sería igualmente razonable sugerir que después de una buena cantidad de ensayos todas las claves sean más o menos equivalentes como predictores de la ubicación de la meta, y que no haya necesariamente competencia entre ellas (Chamizo et al., 2012). Esta posibilidad alternativa queda pendiente de ser estudiada más adelante en anfibios.
En el caso del bloqueo, otro aspecto teórico a considerar ha surgido a partir advertirse la presencia potencial de una variedad de problemas de control. Existe la posibilidad, recientemente demostrada en abejas (Blaser et al., 2006), que diferentes tratamientos con estímulos no objetivo en los experimentos convencionales de bloqueo (con diseños entre grupos) puedan dar la apariencia de bloqueo, independientemente de la experiencia con el estímulo objetivo. Considerando el diseño prototipo de Kamin (1969a), un grupo bloqueo tiene reforzados los ensayos con el estímulo A (ensayos A+) que son seguidos por ensayos reforzados con un compuesto de estímulos A y B (ensayos AB+), mientras que un grupo control tiene reforzados sólo los ensayos con el compuesto AB. En una prueba posterior, el grupo bloqueo responde menos a B de lo que lo hace el grupo control, aunque la contigüidad de B con el reforzamiento ha sido la misma para ambos. En este caso, Blaser y colaboradores (2006) repitieron el mismo procedimiento con sólo una diferencia: B estaba ausente en el entrenamiento y era encontrado por primera vez en la etapa de prueba. En cada caso, aún así, había menos respuesta a B en el grupo cuyo tratamiento correspondía al mismo que el del grupo bloqueo, que en el grupo cuyo tratamiento correspondía al mismo que el del grupo control (un efecto que se ha denominado "pseudobloqueo"). Estos resultados demuestran que diferentes tratamientos de estímulos no objetivo pueden de hecho influenciar la respuesta a B en los ensayos de prueba en forma independiente de la experiencia con B en el entrenamiento. Por lo tanto, estos autores sugieren que una demostración de bloqueo en cualquier experimento convencional (con un diseño entre grupos) puede ser poco convincente si no va acompañado por resultados nulos para grupos controles paralelos sin el estímulo B. Además, esta dificultad podría evitarse mediante la comparación de tratamientos intra-sujeto (Blaser et al., 2008). El análisis de los presentes resultados en anfibios con esta visión, todavía requiere de más información al respecto.
Aún con estas limitaciones en la interpretación de los resultados, es importante destacar que el presente estudio aporta datos valiosos registrados por primera vez en la clase Amphibia.
En el marco de un enfoque comparativo, Spetch (1995) realizó experimentos en palomas y humanos utilizando una pantalla táctil y claves generadas por computadora. Halló que en ambas especies el control adquirido sobre la respuesta (picoteo en las palomas y presionar en los humanos) por una clave a una distancia dada del objetivo, se reducía en presencia de otra clave más cercana. Estos resultados son una clara demostración de ensombrecimiento debido a la proximidad espacial relativa. Este fenómeno de ensombrecimiento también ha sido hallado en humanos utilizando un procedimiento de navegación virtual (Chamizo, 2002).
El efecto de bloqueo también ha sido probado en humanos utilizando una versión computarizada del laberinto de Morris (Hamilton & Sutherland, 1999). Los sujetos inicialmente entrenados para localizar una meta invisible con un determinado conjunto de claves, se desempeñaron pobremente al intentar localizar la meta cuando fueron probados con un grupo de claves agregadas con posterioridad (un claro efecto de bloqueo).
Por otra parte, el bloqueo y el ensombrecimiento entre claves ha sido además encontrado y estudiado extensamente en abejas (Bitterman, 1996; Hammer & Menzel, 1995), lo cual sugiere que son fenómenos presentes en un amplio grupo de seres vivos, incluyendo invertebrados.
El hallazgo de estos dos fenómenos de aprendizaje en anfibios realizado en el presente trabajo se suma al conjunto de resultados anteriores y apoyan la idea de que tanto el bloqueo como el ensombrecimiento tendrían un origen evolutivo temprano.
Al mismo tiempo, estos resultados permiten estudiar a futuro los mecanismos neurales que subyacen a estos fenómenos de aprendizaje espacial en anfibios. Dada la organización más simple del cerebro de este grupo, se pondrá especial énfasis en la búsqueda de patrones funcionales comunes con otras clases de vertebrados, potencialmente presentes en un ancestro común. Al explorar las bases neurales, se ha observado que en los distintos grupos de vertebrados las habilidades de aprendizaje espacial parecen estar asentadas en mecanismos neurales homólogos: como el hipocampo en aves y mamíferos, el córtex medial en los reptiles es crítico para las habilidades de mapeo cognitivo y de manera similar, el pallium telencefálico lateral de peces (el presunto homólogo del hipocampo de amniotas), está selectivamente involucrado en la cognición espacial (Salas et al., 2003). Así, en peces y reptiles, el sustrato neural involucrado en el aprendizaje espacial podría corresponder a estructuras telencefálicas homólogas al hipocampo: el pallium lateral y medial, respectivamente (López et al., 1999; Papini et al., 1999). Entre los tetrápodos, el telencéfalo de los anfibios presenta un nivel de organización primitivo (en el sentido de no derivado). Éste puede dividirse en una región palial o pallium (con una porción medial y una lateral) y otra subpalial (Muzio, 1999, 2012; Rodríguez et al., 1998). Existe una hipótesis muy aceptada con respecto al pallium medial de anuros respecto de que es un área homóloga al complejo hipocampal de otros tetrápodos, basado en la posición topográfica (cuadrante dorsomedial del telencéfalo), sus interconexiones neurales y correspondencia neurohistoquímica (Northcutt & Ronan, 1992). Esto convierte al pallium medial de los anfibios en la estructura clave a investigar a futuro, con el fin de contribuir a una mejor comprensión tanto de su homología como de sus equivalencias funcionales con el hipocampo de otros grupos de vertebrados.
Si bien los datos aportados por el presente trabajo sugieren fuertemente la existencia en anfibios de dos fenómenos generales del aprendizaje como son el bloqueo y el ensombrecimiento, será necesario reunir aún más información sobre los mismos, sumando una mayor cantidad de estudios experimentales para fortalecer las conclusiones halladas. Además, es importante incorporar el estudio de otros fenómenos generales del aprendizaje (como por ejemplo, la inhibición latente) con el fin de evaluar la extensión de estos hallazgos.
Finalmente, la existencia del bloqueo y el ensombrecimiento en una clase de vertebrado flogenéticamente antiguo como los anfibios podría reflejar tanto la existencia de un ancestro común que compartan distintos grupos de vertebrados surgidos por evolución divergente, como su aparición en grupos diferentes por convergencia evolutiva. Para discernir entre estas dos posibilidades, será necesario incorporar a los datos reunidos en el presente trabajo información sobre los mecanismos que subyacen a los comportamientos implicados. Sólo a través de las posibles equivalencias funcionales de las estructuras homólogas implicadas se podrá aclarar esta cuestión.
Referencias
Acebes, F., Solar, P., Carnero, S. & Loy, I. (2009). Blocking of conditioning of tentacle lowering in the snail (Helix aspersa). The Quarterly Journal of Experimental Psychology, 62(7), 1315-1327. doi: 10.1080/17470210802483545 [ Links ]
Artigas, A. A., Aznar-Casanova, J. A. & Chamizo, V. D. (2005). Effects of absolute proximity between landmark and platform in a virtual Morris pool task with humans. International Journal of Comparative Psychology, 18, 224-238. Disponible en: http://www.escholarship.org/uc/item/8m99x7mn [ Links ]
Biegler, R. & Morris, R. G. M. (1999). Blocking in the spatial domain with arrays of discrete landmarks. Journal of Experimental Psychology: Animal Behavior Processes, 25 (3), 334-351. [ Links ]
Bitterman, M. E. (1996). Comparative analysis of learning in honeybee. Animal Learning and Behavior, 24, 123-141. [ Links ]
Blaser, R. E., Couvillon, P. A. & Bitterman, M. E. (2006). Blocking and pseudoblocking: New control experiments with honeybees. Quarterly Journal of Experimental Psychology; 59, 68-76. doi:10.1080/17470210500242938 [ Links ]
Blaser, R E., Couvillon, P. A. & Bitterman, M. E. (2008). Within subjects experiments on blocking and facilitation in honeybees (Apis mellifera). Journal of Comparative Psychology; 122, 373-378. doi:10.1037/a0012623 [ Links ]
Brattstrom, B. H. (1990). Maze learning in the frebellied toad, Bombina orientalis. Journal of Herpetology, 24(1), 44-47. http://www.jstor.org/stable/1564288 [ Links ]
Chamizo, V. D. (2002). Spatial Learning: Conditions and Basic Effects. Psicológica, 23, 33-57. [ Links ]
Chamizo, V D., Manteiga, R D., Rodrigo, T. & Mackintosh, N. J. (2006). Competition between landmarks in spatial learning: The role of proximity to the goal. Behavioural Processes, 71, 59-65. doi:10.1016/j.beproc.2005.11.003. [ Links ]
Chamizo, V. D. & Rodrigo, T. (2004). Effect of absolute spatial proximity between a landmark and a goal. Learning and Motivation, 35, 102-114. doi: 10.1016/S0023-9690(03)00059-6. [ Links ]
Chamizo, V D., Rodríguez, C. A, Espinet, A. & Mackintosh, N. J. (2012). Generalization decrement and not overshadowing among pairs of landmarks in a navigation task. Journal of Experimental Psychology: Animal Behavior Processes, 38, 255-265. doi: 10.1037/a0029015. [ Links ]
Chamizo, V. D., Sterio, D. & Mackintosh, N. J. (1985). Blocking and overshadowing between intramaze and extra-maze cues: a test of the independence of locale and guidance learning. Quarterly Journal of Experimental Psychology, 37B, 235-253. doi: 10.1080/14640748508402098. [ Links ]
Cheng, K. (1986). A purely geometric module in the rat's spatial representation. Cognition, 23, 149-178. [ Links ]
Cheng, K. (1989). The vector sum model of pigeon landmark use. Journal of Experimental Psychology: Animal Behavior Processes, 15, 366-375. doi:10.1037/0097-7403.15.4.366. [ Links ]
Cheng, K., Collett, T. S., Pickhard, A. & Wehner, R. (1987). The use of visual landmark by honeybees: Bees weight landmarks according to their distance from the goal. Journal of Comparative Physiology A, 161, 469-475. doi:10.1007/BF00603972. [ Links ]
Cheng, K. & Spetch, M. L. (1998). Mechanisms of landmark use in mammals and birds. En S. Healy (Ed.), Spatial representation in animals (pp. 1-17). Oxford: Oxford University Press. [ Links ]
Cheng, K. & Spetch, M. L. (2001). Blocking in landmark-based search in honeybees. Animal Behavior, 29, 1-9. [ Links ]
Collett, T. S., Cartwrigth, B. A. & Smith, B. A. (1986). Landmark learning and visuospatial memories in gerbils. Journal of Comparative Physiology A, 158, 835-851. [ Links ]
Daneri, M. F., Casanave, E. B. & Muzio, R. N. (2011). Control of spatial orientation in terrestrial toads (Rhinella arenarum). Journal of Comparative Psychology, 125(3), 296-307. doi: 10.1037/a0024242. [ Links ]
Daneri, M. F., Casanave, E. B. & Muzio, R. N. (2013). Use of local visual landmarks for spatial orientation in terrestrial toads (Rhinella arenarum): The role of distance to a goal. Enviado para su publicación al Journal of Comparative Psychology. [ Links ]
Hamilton, D. A. & Sutherland, R. J. (1999). Blocking in human place learning: Evidence from virtual navigation. Psychobiology, 27, 453-461. [ Links ]
Hammer, M. & Menzel, R. (1995). Learning and memory in the honeybee. Journal of Neuroscience, 15, 1617-1630. [ Links ]
Hull, C. L. (1943). Principles of behavior. N.Y.: Appleton-Century-Crofits. [ Links ]
Kamin, L. J. (1969a). Predictability, surprise, attention and conditioning. En B.A. Campbell and R.M. Church (Eds), Punishment and aversive behavior. N.Y.: Appleton-Century-Crofits. [ Links ]
Kamin, L. J. (1969b). Selective association and conditioning. En N. J. Mackintosh and W. K. Honig (Eds.), Fundamental issues in associative learning (pp. 42-64). Halifax, Nova Scotia, Canada: Dalhousie University Press. [ Links ]
López, J. C., Broglio, C, Rodríguez, F, Tinus-Blanc, C. & Salas, C. (1999). Multiple spatial learning strategies in goldfish (Carassius auratus). Animal Cognition, 2, 109-120. [ Links ]
Loza Coll, M. A. & Muzio, R. N. (2013). Hyperosmotic NaCl solutions as aversive stimuli in the terrestrial toad Rhinella arenarum. Enviado para su publicación al Journal of Experimental Biology. [ Links ]
Mackintosh, N. J. (1975). A theory of attention: Variations in the associability of stimuli with reinforcement. Psychological Review, 82, 276-298. doi:10.1037/h0076778. [ Links ]
Mackintosh, N. J. & Honig, W. K. (1970). Blocking and enhancement of stimulus control in pigeons. Journal of Comparative and Physiological Psychology, 73, 78-85. doi: 10.1037/h0030021. [ Links ]
Milgram, N., Adams, B., Callahan, H., Head, E., Mackay, B., Tirlwell, C. & Cotman, C. (1999). Landmark Discrimination Learning in the Dog. Learning & Memory, 6, 54-61. doi: 10.1101/lm.6.1.54. [ Links ]
Muzio, R. N. (1999). Aprendizaje instrumental en anfibios. Revista Latinoamericana de Psicología, 31, 35-47. [ Links ]
Muzio, R. N. (2012). Aprendizaje en anfibios, el eslabón perdido: Un modelo simple cerebral en el estudio de conductas complejas. Cuadernos de Herpetología, 27(2), Online. [ Links ]
Muzio, R. N., Segura, E. T. & Papini, M. R. (1992). Effect of schedule and magnitude of reinforcement on instrumental acquisition and extinction in the toad, Bufo arenarum. Learning and Motivation, 23, 406-429. doi:10.1016/0023-9690(92)90004-6. [ Links ]
Northcutt, R. G. & Ronan, M. (1992). Afferent and efferent connections of the bullfrog medial pallium. Brain, Behavior and Evolution, 40, 1-16. doi: 10.1159/000113898. [ Links ]
O'Keefe, J. & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford: Clarendon Press. [ Links ]
Papini, M. R., Salas, C. & Muzio, R. N. (1999). Análisis comparativo del aprendizaje en vertebrados. Revista Latinoamericana de Psicología, 31, 15-34. [ Links ]
Pavlov, I. P. (1927). Conditioned reflexes. London: Oxford University Press. [ Links ]
Pearce, J. M. (2009). The 36th Sir Frederick Bartlett Lecture: An associative analysis of spatial learning, The Quarterly Journal of Experimental Psychology, 62(9), 1665-1684. doi: 10.1080/17470210902805589. [ Links ]
Pearce, J. M., Graham, M., Good, M. A., Jones, P. M. & McGregor, A. (2006). Potentiation, Overshadowing, and Blocking of Spatial Learning Based on the Shape of the Environment. Journal of Experimental Psychology: Animal Behavior Processes, 32(3), 201-214. doi: 10.1037/0097-7403.32.3.201. [ Links ]
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian learning: Variations in the effectiveness of conditioned but not of unconditioned stimuli. Psychological Review, 87, 532-552. doi:10.1037/0033-295X.87.6.532. [ Links ]
Rescorla, R. A. (1971). Variations in the effectiveness of reinforcement and nonreinforcement following prior inhibitory conditioning. Learning and Motivation, 2, 113-123. doi:10.1016/0023-9690(71)90002-6. [ Links ]
Rescorla, R. A. & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. En A. H. Black and W. F. Prokasy (Eds.), Classical conditioning II: Current research and theory (pp. 64-99). New York: Appleton-Century-Crofits. [ Links ]
Roberts, A. D. L. & Pearce, J. M. (1999). Blocking in the Morris swimming pool. Journal of Experimental Psychology: Animal Behavior Processes, 25, 225-235. doi: 10.1037/0097-7403.25.2.225. [ Links ]
Rodrigo, T., Chamizo, V. D., McLaren, I. P. L. & Mackintosh, N. J. (1997). Blocking in the spatial domain. Journal of Experimental Psychology: Animal Behavior Processes, 23, 110-118. [ Links ]
Rodríguez, F., López, J. C., Vargas, J. P. & Salas, C. (1998). Fundamentos de Psicobiología. Manual de Laboratorio. Sevilla: Kronos. [ Links ]
Ruibal, R. (1962). The adaptive value of bladder water in the toad, Bufo cognatus. Physiological Zoology, 35, 218-223. Disponible en: http://www.jstor.org/stable/30152806. [ Links ]
Sahley, C. L., Gelperin, A. & Rudy J. W (1981). An analysis of associative learning in a terrestrial mollusc. I. Higher-order conditioning, blocking and a transient US pre-exposure effect. Journal of Comparative Physiology A, 144, 1-8. [ Links ]
Salas, C, Broglio, C. & Rodríguez, F. (2003). Evolution of forebrain and spatial cognition in vertebrates: conservation across diversity. Brain, Behavior and Evolution, 62, 72-82. doi: 10.1159/000072438. [ Links ]
Sanchez-Moreno, J., Rodrigo, T, Chamizo, V. D. & Mackintosh, N. J. (1999). Overshadowing in the spatial domain. Animal Learning and Behavior, 27, 391-398. doi: 10.3758/BF03209976. [ Links ]
Spetch, M. L. (1995). Overshadowing in landmark learning: Touch-screen studies with pigeon and humans. Journal of Experimental Psychology: Animal Behavior Processes, 21, 166-181. doi: 10.1037/0097-7403.21.2.166 [ Links ]
Spetch, M. L., Cheng, K. & MacDonald, S. E. (1996). Learning the configuration of a landmark array: I. Touch screen studies with pigeons and humans. Journal of Comparative Psychology, 110, 55-68. [ Links ]
Stahlman, W. D. & Blaisdell, A. P. (2009). Blocking of spatial control by landmarks in rats. Behavioural Processes, 81, 114-118. doi: 10.1016/j.beproc.2009.02.007. [ Links ]
Tennant, W. A. & Bitterman, M. E. (1975). Blocking and overshadowing in two species of fish. Journal of Experimental Psychology: Animal Behavior Processes, 1, 22-29. doi: 10.1037/0097-7403.1.1.22. [ Links ]
