










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.18 no.2 Medellín July/Sep. 2005
Importancia del manejo de hongos micorrizógenos en el establecimiento de árboles en sistemas silvopastoriles
Mauricio Molina L1, Zoot, Esp; Liliana Mahecha L2, Zoot, MSc; Marisol Medina S2, I.A., MSc.12
Aspirante Maestría en Ciencias Animales Universidad de Antioquia, Secretaría de Agricultura y Desarrollo Rural de Antioquia, Docentes
Universidad de Antioquia. Facultad de Ciencias Agrarias.
(Recibido: 2 marzo, 2004; aceptado: 13 junio, 2005)
Los sistemas silvopastoriles han sido reportados como una alternativa viable para la ganadería tropical. Sin embargo, su aceptación e implementación por parte de los productores no ha sido consecuente con el desarrollo investigativo. Una de las principales causas de la lenta implementación, es el largo período en que deben permanecer en descanso los potreros luego de la siembra de los árboles. Esta situación ha generado el planteamiento de estrategias tales como el uso de microorganismos que permitan estimular el crecimiento de los árboles. En este artículo se hace una revisión bibliográfica sobre el funcionamiento, importancia, tipos y efectos del uso de micorrizas en el establecimiento de árboles. La revisión permite demostrar que en Colombia las micorrizas tienen gran potencial para contribuir a una producción sostenible. Sin embargo, se resalta la importancia de profundizar en la investigación sobre el uso de micorrizas nativas o del sitio, lo que implica conocer más sobre la relación hongo-planta, buscando aislar cepas específicas que permitan potencializar el crecimiento y productividad de los árboles en determinados agroecosistemas, y evaluar las interacciones que se generan.
Palabras clave: ganadería, micorrizas, microorganismos, sostenibilidad, suelo.
Introducción
Entre las principales causas atribuibles a la baja Los suelos de estos sistemas se caracterizan por productividad y pérdida de la biodiversidad en los tener un pH bajo, alto contenido de aluminio y sistemas de producción bovina en los trópicos se manganeso, bajos en bases y alta capacidad de encuentra la degradación de las pasturas, relacionada retención de fósforo. Por lo tanto, los rendimientos no con la destrucción de los bosques y de las selvas son buenos, lo que se traduce en baja producción donde tropicales, mediante la tumba y la quema, para dar en la mayoría de los casos se llega a la insostenibilidad paso a las praderas de gramíneas dirigidas a la y se afecta la competitividad (15, 40, 50). producción de bovinos. Además, está asociada en la incorporación de suelos con menor fertilidad, en los La problemática anterior indica que se requiere que se plantan especies no adaptadas, generándose establecer sistemas de producción sostenibles y así mayor proporción de pasturas degradadas y poco adaptables a nuestras regiones, entre los que se productivas; estas condiciones hacen ineficientes los encuentran los sistemas silvopastoriles, los cuales actuales sistemas de producción animal y afectan representan una alternativa viable para la ganadería negativamente el medio ambiente (30). bovina. En estos sistemas se han encontrado múltiples beneficios reportados a través de diversas investigaciones (27, 38, 47). No obstante, su adopción e implementación por parte de los productores aún es muy lenta, relacionándose en gran parte, con los largos períodos de tiempo que el productor debe dejar sin utilizar los potreros luego de la siembra de los árboles y los arbustos. El productor generalmente debe esperar entre ocho meses y un año como mínimo, según la especie y el uso al que se va a destinar (ramoneo, sombra, maderable, otros), si se introducen antes los animales; cuando aún los árboles no están debidamente anclados con sus raíces y no han alcanzado la vigorosidad y altura suficiente que impida el daño por los animales, su sobrevivencia se verá comprometida y con ésto, el éxito del sistema.
La situación es aún más crítica en sistemas ganaderos de trópico alto, en los cuales el tamaño de los predios es pequeño, dificultándose la destinación de áreas para establecimiento de árboles, lo que conlleva a un aislamiento del área por un largo periodo de tiempo, el cual va desde el momento de siembra hasta el inicio de pastoreo por parte del animal, mientras los árboles crecen. Además, en estas zonas ubicadas a más de 1800 m.s.n.m, la luminosidad disminuye (46), la tasa de crecimiento de los árboles es menor comparada la de los árboles que se establecen en zonas de trópico medio y bajo, y por lo tanto, se retarda aún más el uso de los sistemas silvopastoriles. Lo anterior ocasiona que el ganadero no implemente el sistema, debido a que este no puede esperar largos períodos de tiempo, por las exigencias productivas que exigen los predios de las estas zonas trópico alto.
Esta situación ha generado el planteamiento de estrategias que permitan reducir el período de espera, tales como: uso de especies de rápido crecimiento y desarrollo, aislamiento de los árboles con cercas fijas o cerca eléctrica, fumigación con estiércol bovino, y uso de microorganismos para estimular su crecimiento. Esta última opción ha tomado gran auge en los últimos años porque además de ser una opción ecológica, puede ser utilizada de forma integral en la solución del problema. Al respecto, Pate (39) considera, que el uso adecuado de los microorganismos del suelo, permite lograr una agricultura sostenible que resulta práctica y económica para cada unidad agrícola y también favorece el reciclaje de nutrientes para mejorar la fertilidad del suelo; por lo tanto, se convierte en alternativa para contribuir al establecimiento de sistemas de producción sostenibles, competitivos y rentables.
Entre los microorganismos de mayor uso se destacan los hongos micorrizógenos. Estos organismos pueden ser aislados, seleccionados, multiplicados e incorporados al suelo o a las plantas en forma de inóculos. El proceso de inoculación es complejo. Implica diseñar métodos de aislamiento, selección, multiplicación e incorporación adecuados para cada especie o efecto deseado y por otra parte, es necesario determinar las condiciones y técnicas culturales que permitan la manifestación óptima de los efectos. Esta complejidad hace que su efecto no sea predecible bajo todas las condiciones ni para todas las especies. Por lo tanto, es importante incentivar la investigación al respecto, así como profundizar en el conocimiento de sus principios de funcionamiento y de los resultados encontrados con su uso.
Con base en lo anterior, se planteó la necesidad de realizar una revisión bibliográfica sobre la definición, funcionamiento, importancia, tipos y efectos del uso de micorrizas en árboles que pueda ser utilizada como base teórica para el inicio de un trabajo de investigación mediante el cual se haga evaluación del efecto de la inoculación con micorrizas, en el crecimiento y el desarrollo de árboles en sistemas silvopastoriles de trópico alto.
Definición, funcionamiento e importancia de las micorrizas
Varios autores coinciden en definir a las micorrizas (mikes=hongo, rhiza=raíz) como asociaciones mutualistas entre un hongo y las raíces de la planta, en la que ambos miembros de la asociación se benefician y participan activamente en el transporte y absorción de nutrientes, influyendo tanto en la estructura como en la estabilidad de las comunidades vegetales (5, 28, 32). En la asociación mutualista que se establece con la micorriza, el hongo coloniza biotróficamente la corteza de la raíz, sin causar daño a la planta, llegando a ser, fisiológica y morfológicamente, parte integrante de dicho órgano. A su vez, la planta hospedera proporciona al hongo simbionte (heterótrofo), compuestos carbonados procedentes de la fotosíntesis, y un hábitat ecológico protegido.
Se sabe que las micorrizas juegan papel muy importante en el desarrollo de las plantas y en el ciclado de nutrientes en el ecosistema. Se encuentran prácticamente en todos los suelos y climas de la tierra y sólo en unas pocas familias botánicas hay especies que no forman micorrizas. Los ejemplos más significativos de familias con especies no micorrizables son las crucíferas, quenopodiáceas y ciperáceas (1, 3, 11). Al respecto, Infante (23), estima que aproximadamente el 85-95% de las especies vegetales conocidas viven en simbiosis con una gran cantidad de hongos del suelo.
Estas formas simbióticas se inician con la activación del micelio del hongo procedente de la germinación de las esporas, dando comienzo a la formación de la micorriza, lo que genera los propágalos para lograr en forma natural perpetuar y propagar la especie. Cuando las esporas germinan, desarrollan unos filamentos conocidos como hifas, que por proliferación dan lugar al micelio del hongo que se extiende en el suelo según su potencial saprofítico; el micelio es el encargado de llevar a cabo la infección de la raíz, que se produce por una identificación mutua, en primera instancia, entre la planta y el hongo en la rizósfera en regiones próximas a las raíces; este reconocimiento es mediado por sustancias exudadas por la raíz, que provocan el crecimiento del micelio y un biotropismo positivo del mismo hacia la raíz. Luego se produce el contacto intercelular al formarse una estructura llamada apresorio; posteriormente se producen cambios morfológicos y estructurales, tanto en los tejidos colonizados por el hongo, como en la organización de la pared celular del simbionte fúngico. El proceso continúa con la integración fisiológica de ambos simbiontes, y por último, se presenta una alteración de las actividades enzimáticas, para integrar los procesos metabólicos (3, 32).
Se reporta que el micelio activado coloniza los tejidos de la raíz y las células corticales de la misma, donde en algunas micorrizas el hongo forma ramificaciones intracelulares llamadas arbúsculos, que sirven para llevar a cabo el intercambio de metabolitos entre el hongo y la planta (28). Las hifas del hongo crecen extensivamente desde la raíz hacia el suelo, donde desarrollan una red tridimensional de micelio especializado en colonizar y explorar muy eficazmente los microhábitats del mismo, para captar elementos minerales y agua. Las hifas externas del hongo actúan realmente como «puentes», que superan la zona de «agotamiento» en nutrientes que rodea la raíz y llega a distancias de varios centímetros de la superficie de la misma. Se considera que la función del micelio externo es particularmente crítica para la captación de nutrientes poco móviles, como son fósforo, nitrógeno y algunos micronutrientes (28).
El término micorriza fue propuesto por primera vez en 1885, por Albert Bernard Frank, quien descubrió la asociación regular de tejidos fungosos (manto) con el tejido radical de los árboles (47). Sin embargo, los descubrimientos fósiles, en los que se han encontrado micorrizas llamadas fósiles endomicorrizales, hacen pensar que este tipo de asociaciones han podido ser cruciales para permitir la colonización terrestre de las plantas y que por lo tanto su existencia data de mucho tiempo atrás (32, 48). No obstante, sólo a partir de 1950, que se inicia a nivel mundial la investigación básica sobre su uso, profundizándose en el tema desde el año 1970. Investigaciones realizadas (3,4,43), han logrado demostrar que las micorrizas son capaces de beneficiar al 95% de todas las especies existentes y que su aplicación produce mejoras significativas para el crecimiento de árboles nativos, frutales, café, banano, cultivos agrícolas, hortalizas, ornamentales, caña, palma africana, pastos para pastoreo y de corte. A pesar de esto, Raddattz (47) es de la opinión que en Latinoamérica, y en Colombia, han sido subutilizadas, debido a que se ha limitado su aplicación a sólo cuatro cultivos: pastos, yuca, fríjol y arroz, en los cuales las micorrizas no destacan su total potencialidad.
Principios fisiológicos de los beneficios de las micorrizas a las plantas asociadas
Entre los beneficios que producen las micorrizas en las plantas asociadas, se pueden destacar los siguientes: facilitan su nutrición, crecimiento y desarrollo; mejoran su tolerancia frente al estrés hídrico y a los agentes patógenos; facilitan su adaptación a suelos salinos y contribuyen con la disminución de la erosión en las áreas aledañas (28, 47, 49). Estos beneficios son la base fundamental que le permite a la planta mayor adaptación al medio, mayor competitividad con las plantas acompañantes no micorrizadas y mayor productividad.
Para mayor entendimiento de los beneficios de las micorrizas a continuación se explican cada uno de ellos:
Facilitan la nutrición, crecimiento y desarrollo
Bolan (5) y Fitter (18) consideran que el beneficio de las micorrizas se traduce en mayor crecimiento y desarrollo de las plantas en beneficio de la adaptación y eficiencia de éstas al facilitar una mayor absorción de nutrientes minerales del suelo (nutrición). Este beneficio se basa en que las plantas requieren de diferentes iones de minerales para un óptimo crecimiento y desarrollo. Las micorrizas pueden ayudar a las plantas a incrementar esta captación, tanto de forma directa como indirecta. De forma directa, las micorrizas aumentan en las raíces la toma de nutrientes minerales y agua del suelo debido a que poseen micelio externo que explora gran volumen de suelo que no está micorrizado (28, 47, 49). En este proceso se acepta que el papel clave de las micorrizas radica en que las hifas del hongo extienden el campo de absorción de la raíz más allá de la zona normal de agotamiento radicular (en 1-5 mm), y permiten a la raíz incrementar su superficie de absorción y explorar un volumen de suelo mayor del que lo hacen las raíces no micorrizadas, concretamente hasta 7 cm de la superficie radicular. Además, se ha logrado poner de manifiesto que las raíces micorrizadas absorben más eficazmente los fosfatos que las no micorrizadas y han calculado que en un centímetro de raíz micorrizada posee unos 80 cm de hifas externas. Sin embargo, Miyasaka (32) en estudios realizados concluyó, que la absorción más eficiente por las raíces micorrizadas se debe fundamentalmente a una aceleración de la disociación del fosfato insoluble. De igual forma, Maldonado (28), considera que las micorrizas permiten lograr una mayor absorción de nutrientes en la solución del suelo, en especial de elementos pocos móviles, fundamentalmente el fósforo y otros como zinc, azufre, calcio, molibdeno, boro. Miyasaka (32) y Velasco et al (50), igualmente destacan que el papel de las micorrizas es ayudar a las plantas a incrementar la eficiencia de la utilización del fósforo. Así mismo, Raddattz (41), Londoño (26) y Pate (39) reportan que en la asociación, el hongo recibe de la planta productos fotosintéticos (fotosintatos), los cuales son adquiridos por el hongo en un porcentaje de 1 a 12% de todos los fotosintatos asimilados por la planta, los que a su vez son intercambiados en las células del parénquima de la raíz por moléculas que contienen fósforo, nitrógeno, potasio, magnesio, zinc, azufre, calcio, entre otros. Miyasaka (32) presenta parámetros como la longitud, el diámetro, el área de superficie y la densidad de vellosidades de la raíz, importantes para aumentar los nutrientes tomados por la planta. Consecuentemente, cambios que traen las micorrizas en la morfología de la raíz como la ramificación y elongación, podrían constituir un mecanismo adicional para la absorción de fósforo. Sin embargo, el mecanismo por el cual las micorrizas modifican la morfología de la raíz aún no es claro (39).
Estos reportes muestran cómo el fósforo se convierte en un elemento fundamental en los beneficios de las micorrizas. Esto es debido a que el micelio de los hongos endomicorrízicos puede crecer más allá de la zona de crecimiento radical de la planta que permite incrementar el volumen de suelo de donde la planta absorbe el fósforo. La zona de crecimiento radical es una zona en donde los iones se agotan rápidamente y en donde debido a la tasa de difusión tan baja de fósforo, en condiciones normales, no alcanza a ser aprovisionada en forma adecuada (50). La transferencia de fósforo inorgánico entre el hongo micorrícico y la raíz huésped se produce en tres etapas: transporte activo dentro del hongo y el exterior de la raíz, transporte pasivo del hongo en la interfase raízhongo y transporte activo dentro de la raíz (5,19).
Los beneficios de los hongos micorrícicos en la captación de fósforo del suelo tiene especial importancia si se tiene en cuenta que el fósforo es esencial para el crecimiento de las plantas y que es absorbido casi enteramente en forma inorgánica. No existe otro nutriente que pueda sustituirlo y aunque de los elementos primarios es el requerido en menor cantidad, su disponibilidad en la mayor parte de los suelos agrícolas del trópico es limitada (14). El fósforo es constituyente de ácidos nucleicos, fosfolípidos, vitaminas, es indispensable en procesos donde hay transporte, almacenamiento y transformación de energía; actúa también en la fotosíntesis, respiración, división y elongación celular. Otras de sus funciones son estimular la formación temprana y el crecimiento de las raíces, intervenir en la formación de los órganos de reproducción de las plantas, la formación de semillas y acelera la maduración de los frutos en los cuales generalmente se almacena en altas concentraciones (14,47). La falta de fósforo en una planta retarda el crecimiento de la parte aérea y reduce el de las raíces y trae como consecuencia baja producción del cultivo (14, 32, 47). En los trópicos, la producción es seriamente limitada por la deficiencia en los suelos, fósforo como también de nitrogeno y de agua (4).
Además de las explicaciones dadas sobre por qué los hongos micorrícicos facilitan de forma directa la mayor captación de nutrientes por parte de la planta y repercuten en su crecimiento y desarrollo, Ingham (22) considera que se debe a cierto tipo de hongos micorrícicos que colonizan células por fuera de las raíces y extienden largos hilos o hifas, formando un eslabón entre el suelo y la raíz.
En cuanto al efecto indirecto de las micorrizas, ha sido relacionado con la facilitación del proceso de absorción normal por parte de la planta, al mejorar el reciclaje de nutrientes (22). Maldonado (32) plantea otro efecto indirecto como es el aumento en la eficiencia de otros microorganismos que tienden a asociarse con las micorrizas, tales como Rizhobium, Azospirillum, Azotobacter, que a su vez incrementan la captación de nutrientes para las plantas (32). Fitter (18), encontró que las micorrizas interactúan con otros organismos del suelo y estas interacciones pueden inhibir (crean competencia entre ellos) o estimular (forman asociaciones mutualistas). Sin embargo, en muchos casos no ha sido posible definir completamente dichas interacciones. Entre los organismos mencionados pueden estar hongos patógenos (a nivel del micelio interno), protozoarios, nemátodos y artrópodos.
Además de las explicaciones mencionadas, Alarcón y Ferrera (2), reportan informes de otros autores que indican que el beneficio no sólo se debe al establecimiento de los hongos en el sistema radical, sino que también intervienen diversos factores edáficos y ambientales e incluso de manejo de los agroecosistemas, que interactúan potencializando la capacidad del hongo para compensar o superar las funciones de la raíz en la absorción de nutrimentos y agua.
Igualmente, Raddattz (41), reporta la producción de hormonas estimulantes para el crecimiento y desarrollo de las plantas, aunque Miyasaka (32) manifiesta que resulta extremadamente difícil diferenciar los efectos producidos por las hormonas del hongo, de los producidos por hormonas vegetales y de los producidos indirectamente por el estado nutricional de las plantas como consecuencia de la micorrización.
Mejoran la tolerancia al estrés hídrico
Una de las mayores bondades que tienen las micorrizas es su capacidad de absorción de agua y por lo tanto, permiten mayor resistencia de la planta a la sequía (14,41). Esta resistencia es debida al incremento de la conductividad hídrica de la planta o a la disminución de la resistencia al flujo de agua a través de ella. También ha sido relacionado con la mayor absorción a través de la extensa red de hifas externas del hongo, extendidas más allá de la zona a la cual tiene acceso directo el sistema radical (3). De igual forma, se ha encontrado que en suelos arenosos, con poca capacidad de retención de agua, la presencia de micorrizas ha ocasionado retención de agua cinco veces mayor que en ausencia de micorrizas en los mismos (26,41).
Mejoran la tolerancia frente a patógenos
Varios autores concuerdan en que las micorrizas pueden contribuir a la salud de la planta y a su productividad al aumentar el desarrollo de tolerancia a enfermedades y parásitos (28,29,32). Las ectomorrizas protegen la raíz ya que reciclan los carbohidratos, aminoácidos y otros compuestos producidos por las raíces, capaces de atraer agentes patógenos. Además proveen una barrera física a patógenos debido a la formación del manto, en el cual las hifas individuales o cordones de hifas crecen hacia afuera, introduciéndose en el suelo, y hacia adentro, intercalándose entre las células del córtex de la raíz a través de la lámina media, formando un entramado denominado red de Harting. En esta red el micelio deja de estar tabicado, es xenobiótico, lo que se interpreta como ventaja para acelerar los procesos de intercambio. La red de Harting puede sintetizar compuestos como el diatretinenitrilo, con efecto de tipo antibiótico. Así mismo, Raddattz (41) reporta tolerancia contra ataques de nemátodos.
Khanizadeh (25) y Fitter (18) encontraron mejor control de enfermedades y la disminución en gasto de insecticidas y funguicidas, con el uso de plantas micorrizadas.
En investigaciones realizadas, Ingham (22) encontró que la mezcla de bacterias del genero Rhizobium con micorrizas, dan mayor vigor a la planta , lo cual la hace más resistente al ataque de patógenos, porque las micorrizas forman una barrera física impenetrable en la superficie de las raíces, variando en grosor y densidad. Además se encontró que especies de micorrizas parasitan a otros hongos como es el caso del Trichoderma y lo destruyen. Sin embargo existen interacciones donde las micorrizas pueden sobrecolonizar la planta y convertirse en parásitos (18).
Facilitan la adaptación a suelos salinos
La salinidad es un factor limitante de la producción agrícola, los estudios sobre micorrizas y tolerancia de las plantas a este estrés es relativamente reciente. Resultados reportados por Barea (3), demuestran el efecto inducido por la micorrización en la disminución de la deficiencia nutritiva provocada por antagonismos iónicos, efecto secundario del estrés salino, lo que permite crecimiento mayor de las plantas micorrizadas. Concretamente, las micorrizas mejoran diversos procesos fisiológicos (incremento del ritmo de intercambio de CO2, transpiración, cambios en la conductancia estomática, eficacia en el uso de agua), aparte del derivado de la captación de nutrientes. Adicionalmente, Corredor (3) y Barea (14), sugieren otros mecanismos para justificar el papel de las micorrizas en relación con la tolerancia a salinidad, tales como la inducción de cambios hormonales o la mejora en la capacidad de agua.
Contribuyen con la disminución de la erosión en las áreas aledañas
Se sabe que tanto el medio agrícola como los ecosistemas naturales pueden ser afectados por procesos de degradación de diversa índole, en cuanto a su origen y naturaleza, que inciden en la productividad y calidad de las cosechas y/o en la estabilidad, diversidad y productividad de los ecosistemas. La utilización de micorrizas no sólo ha facilitado mejor revegetalización en condiciones particulares, como pueden ser la recuperación de suelos degradados por la minería y la introducción de especies exóticas en distintas partes del mundo, sino también ha mejorado la repoblación de especies vegetales en suelos forestales (3). Lo anterior es confirmado por Londoño (26) y Rodríguez (44), quienes manifiestan que cuando se aplican hongos edáficos, la pérdida por lixiviación, fijación y erosión se disminuye, dado que la red de hifas captura y trasloca elementos nutritivos hacia la planta desde sitios no explorados por la raíz; así en los sistemas selváticos de los trópicos húmedos, el reciclaje de nutrientes de la materia orgánica descompuesta hacia la planta, la hacen los hongos, principalmente por los sistemas micorrícicos.
Tipos de micorrizas
Las micorrizas han sido agrupadas, con base en la anatomía de las plantas colonizadoras en: ectomicorrizas, endomicorrizas y un grupo intermedio denominado ectoendomicorrizas (2, 46, 47).
Ectomicorrizas o micorrizas ectotróficas
En estas micorrizas las hifas del hongo envuelven los segmentos de raíces colonizadas y se entretejen alrededor de ellas formando una estructura anatómica denominada manto, característica de esta simbiosis; a partir del manto, se desprenden hifas que colonizan el medio y cordones hifales denominados rizomorfos, los cuales pueden dar origen a cuerpos fructíferos epígeos o hipógeos. En general, en la literatura revisada los hongos asociados pertenecen a las subdivisiones Basidiomycotina y Ascomycotina (9, 47, 48).
Las ectomicorrizas son relativamente poco frecuentes: entre un 3 y un 5 % de las plantas terrestres establecen este tipo de simbiosis. Su mutualismo se ha reportado especialmente con plantas de importancia forestal, como los pinos, robles, abedules, encinas, sauces, tilos o nogales (22,46).
Endomicorrizas o micorrizas endotróficas
Se caracterizan por la penetración del hongo inter e intracelularmente, la ausencia de manto y las acentuadas modificaciones anatómicas en las raíces no visibles a simple vista (48).
Dentro de las endomicorrizas se distinguen varios tipos:
1. Micorrizas Arbusculares (MA)
Deben su nombre a las estructuras que producen; son organismos que se asocian con las especies vegetales en el área de vegetación nativa y cultivos comerciales (7).
En las arbusculares se encuentran asociados hongos de la clase Zygomycetes, orden Glomales y Endogonales (46, 48, 49). De acuerdo a Sánchez et al (47), en los trópicos las endomicorrizas arbusculares son diez veces más abundantes que las ectomicorrizas y en la mayoría de las especies vegetales donde se reportan, se establece que más del 90% de las especies existentes en el planeta están micorrizadas cuando crecen en condiciones naturales. El 97% de las fanerógamas, incluidas casi todas las especies de interés agronómico, pastoril y selvático, presentan este tipo de asociaciones (12,46,49).
Varios autores reportan que las endomicorrizas predominan en las plantas herbáceas, muchas de ellas de interés agrícola, como trigo, maíz, legumbres, verduras, entre otras, en algunas leñosas, como naranjos, manzanos, cerezos, ciruelos, plataneras y árboles más comunes (12,48,52).
Sólo se ha reportado ausencia en familias como Aizoceae, Amaranthaceae, Fumariaceae, Pinaceae, Caryophyllaceae, Caparceae, Cruciferae, Chenopodiaceae, Commelinaceae, Cyperaceae, Juncaceae, Nyctaginaceae, Phytolaccaceae, Portulaceae, Proteaceae, Resitonaceae, Urticaceae, Zypophyllaceae y Betulaceae (32). Sin embargo, varios autores reportan especies de micorrizas en el árbol aliso, el cual pertenece a la familia de las Betulaceae (46, 47, 49).
2.Micorrizas Ericáceas
Son del grupo de hongos ascomycetos, afectan a las plantas del orden Ericales, que son plantas leñosas arbustivas que viven en suelos ácidos y pobres en nutrientes; también afectan a plantas del orden Monotropaceae (49).
3. Micorrizas Orchidaceas
Están formadas principalmente por el hongo Rhizoctonia solani y afectan a las plantas de la familia Orchidiaceae (22,48,49).
Ecto-endomicorrizas
Generalmente presentan las características de ectomicorrizas, con la diferencia que hay penetración intracelular. Algunos autores las localizan como endomicorrizas, mientras que otros, basándose en la cercanía filogenética de los hongos asociados con los Asco y Basidiomycotina, las ubican como ectomicorrizas. Estas se encuentran en algunos subgrupos de Pinaceae y de Ericales, como los géneros Arbutus (46,48,49).
En muchas regiones geográficas del mundo y del país, todavía los hongos micorrizógenos no han sido caracterizados, por lo cual es posible que se puedan descubrir nuevas especies en los hábitat. Además, se debe establecer la evolución de las micorrizas con respecto a la correlación entre la planta huésped y el hongo simbionte.
Se debe tener en cuenta hoy en día las ventajas que nos ofrece la biotecnología, la diversidad de hongos MA no ha sido investigada intensivamente a pesar de que se ha encontrado que la presencia de micorrizas presenta una relación directa con la diversidad florística y los ciclos de carbono y fósforo en comunidades naturales. Esta situación se explica por la dificultad que implica la identificación de hongos MA. Aunque la morfología de las esporas ha sido usada con propósitos de identificación, esta debe ser realizada a partir de cultivos puros (a partir de una sola espora), proceso que es lento e impredecible debido a que es posible que las esporas extraídas no sean viables (14).
Por estas razones, los métodos moleculares pueden ser útiles en la identificación de especies. La reacción en cadena de la polimerasa (PCR) permite amplificar DNA de una sola espora, y las técnicas basadas en PCR han sido utilizadas para amplificar DNA a partir de hongos MA e identificar materiales colectados incluso a nivel de especie. Los requerimientos de cualquier método molecular aplicado a la investigación de la diversidad y ecología de hongos MA, tienen que ver con la reproducibilidad usando DNA de una sola espora de hongo MA, que sean rápidos, fáciles y económicos para permitir el trabajo de muchas muestras (15).
Resultados encontrados con el uso de micorrizas en árboles
Estudios realizados por Vilar et al (51), referentes a valores de colonización o infestación de micorrizas, indican que son componentes importantes para desarrollar modelos de agricultura sostenible y en especial en sistemas donde existe elevada fertilización de fósforo proveniente de fertilizantes solubles en suelos de bajo pH. Estos resultados concuerdan con los reportados por Colozzi (13), quien evaluó el efecto de intercalar leguminosas en verano para abono verde, en la aparición de micorrizas en café y encontró incremento en la diversidad de especies de micorrizas, lo que a su vez estuvo relacionado con el incremento en la producción y la sostenibilidad del sistema de producción de café.
Ingham (22), reporta efectos benéficos en el establecimiento de plantaciones de Pino en Puerto Rico. En este país, el establecimiento de pinos por muchos años, fue una actividad difícil, debido a que los pinos no son nativos y la micorriza para esta especie en condiciones naturales se encontraba ausente en el suelo. Sin embargo, una vez inoculadas las plantas, estas sobrevivieron después de un año de establecidas mientras que las no inoculadas murieron.
Así mismo, Barrera et al ( 4 ), evaluaron el efecto de la aplicación de micorrizas y de inóculos en el desarrollo de plántulas de Chachafruto, Eryhrina edulis, en etapa de vivero en cuanto a: tipo de micorriza y sustrato utilizado, la altura de la planta, el porcentaje de infección de raíces por la micorriza, el peso seco de la parte aérea de la planta y el peso de la raíz de la plántula.
En todos los tratamientos, aún en el testigo (suelo cafetero) y en los que no se aplicó directamente inóculo, al momento de la evaluación estaban presentes géneros de micorrizas incluidos en la tabla 1, indicando ésto, su alta presencia en los suelos donde se realizó la investigación, mostrando su importante influencia en el desarrollo de las plantas. Además, se observó en relación con la altura, que los valores para todos los tratamientos no tuvieron variación significativa con respecto al testigo (suelo cafetero) (veáse Figura 1)

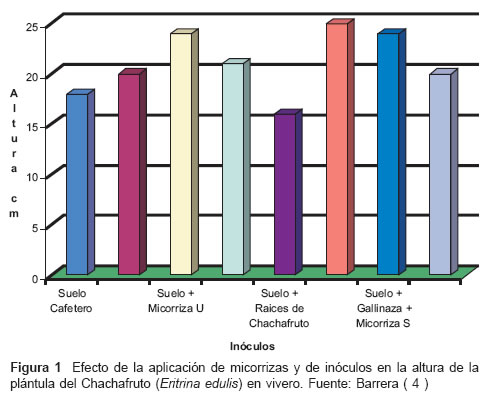
Por otro lado, Infante (23), cuantificó los niveles de infección micorrícica e identificó las especies de hongos micorrícicos más frecuentes asociadas con árboles de pijuayo (Bactris gasipaes H.B.K., especie potencial para utilizar en sistemas silvopastoriles) en sistemas de producción de cultivos arbóreos en multiestrato y en monocultivo, en suelos ultisoles en Yurimaguas, Perú.
El nivel de infección micorrícica fue de 46% en suelos de textura areno franca y 80% en textura franco-arenosa. Esta diferencia por la textura de estos suelos pueden estar correlacionadas con la disponibilidad de fósforo en las parcelas (10.8 ppm en la areno-franca contra 4.8 ppm en la franco-arenosa), en el que es mayor en el suelo franco-arenoso por contener mayor cantidad de arcilla; las mismas tendencias con respecto a textura del suelo se observaron en parcelas de pijuayo en monocultivo, sin embargo, los niveles de infección micorrícica fueron más bajos.
La diversidad de especies de hongos micorrícicos fue diferente en los dos sistemas, la que puede estar influenciada por su composición florística, las especies del género Glomus predominan en pijuayo. En este trabajo se demostró el efecto benéfico de la inoculación con micorrizas en el crecimiento y en el contenido de nutrimentos en plántulas de pijuayo (veánse Tablas 2 y 3).
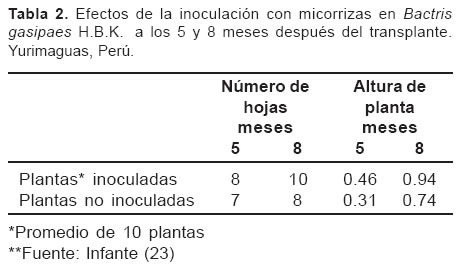
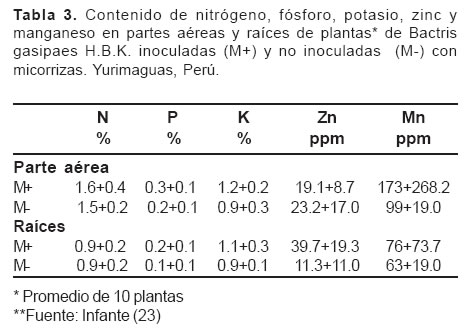
En otras investigaciones, Ramírez et al (42) evaluaron los efectos de Glomus fistulosum, Pseudomonas fluorescentes y fosfato, sobre el crecimiento y la absorción de fósforo por Leucaena leucocephala sembrada en un suelo andisol. La inoculación con G. fistulosum incrementó el crecimiento de las plantas y la absorción de fósforo. Los efectos fueron mayores cuando se aplicaron conjuntamente Pseudomonas y G. fistulosum. La absorción de fósforo estuvo significativamente correlacionada con la longitud de raíces, que a su vez fue significativamente afectada por la inoculación con G. fistulosum. En ausencia de G. fistulosum la aplicación de fósforo no incrementó significativamente su absorción por las plantas. En contraste, cuando este hongo micorrícico fue inoculado, hubo aumento significativo en la cantidad de fósforo absorbido, siendo este incremento mayor con contenido de 30 mg de fósforo por kg que con 15 mg de fósforo por kg.
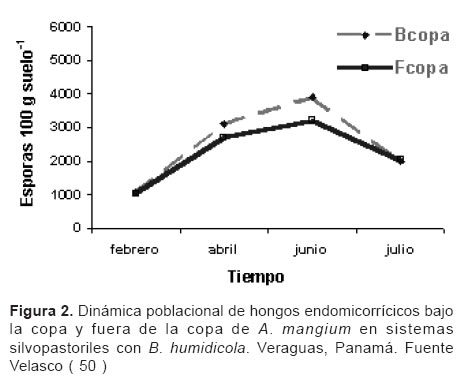
De igual manera, Velasco et al ( 50 ), investigaron el mejoramiento del suelo por Acacia mangium en un sistema silvopastoril con Brachiaria humidicola, donde encontraron una relación directa entre el contenido de fósforo edáfico y la población de hongos endomicorrícicos; el número de esporas bajo la copa fue mayor que fuera de ella, aunque no existen diferencias estadísticas. Cuando la humedad en el suelo bajo el efecto de los árboles fue mayor, se favoreció la población de hongos micorrizógenos. Durante los meses húmedos (junio y julio) la población de hongos endomicorrícicos en el sistema de alta densidad superó al de baja densidad en 79 %. (véase Figura 2).
Con el fin de investigar la actividad de las micorrizas en condiciones de vivero, Reyes (43 ), evaluó el efecto de Glomus spp., bacterias, vermicomposta y un testigo en el desarrollo de plántulas de aguacate. Los sustratos fueron una mezcla de suelo agrícola y de arena de río (1:1 v/v) y uno referencia que era un suelo forestal; la evaluación del efecto se hizo a los 200 días después del transplante. La altura y diámetro del tallo se favorecieron con los tratamientos de vermicomposta y la micorriza Glomus. La vermicomposta y la multicepa Glomus spp., promovieron mayor número de hojas, mayor superficie de área foliar y peso seco de la parte aérea con respecto al testigo. El resto de los tratamientos superaron o al menos igualaron la respuesta de las plantas que se desarrollaron en el tratamiento testigo, sin inoculación microbiana ni aplicación de materia orgánica (véase Tabla 4). En el caso de la fotosíntesis, se observó mayor actividad en los tratamientos que tuvieron la presencia de la micorriza y la bacteria, y la más baja en los tratamientos testigo y de referencia. La colonización de la raíz por estos microorganismos probablemente aumentó la actividad fotosintética de las plantas, debido a que pudo existir demanda extra de fotosintatos por parte de ellos.
La altura de la planta fue favorecida con la vermicomposta, así como la inoculación de hongos micorrícicos en sus diferentes combinaciones, observándose diferencias con respecto al testigo y tratamiento-sustrato de referencia. Se observa que la bacteria sola o en combinación con la micorriza o vermicomposta no favorecen el desarrollo de la altura. La vermicomposta y la micorriza actuando en forma aislada tuvieron un efecto mayor en la promoción de altura. En varias investigaciones se ha demostrado la eficiencia de los hongos micorrícicos del género Glomus en promover el desarrollo de plántulas.
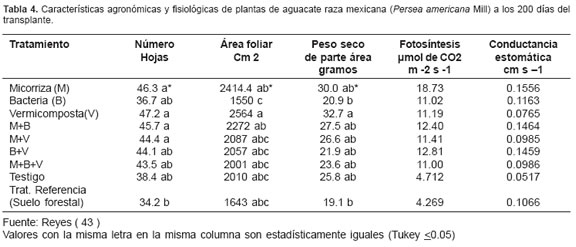
Con respecto a la fertilización con fósforo, Rodríguez et al (44) y Ramírez et al (42), encontraron efecto positivo en la actividad de la micorriza y en el crecimiento de las plantas. Estos autores encontraron respuesta de la Leucaena a la inoculación micorrizal, aún con niveles altos de fósforo en la solución del suelo.
A pesar de los reportes del efecto positivo del fósforo en la actividad de las micorrizas, algunos autores indican un efecto contradictorio. Rodríguez (42) reporta que la micorrización es generalmente inhibida en suelos con un alto contenido en fósforo.
Manejo de la micorrización controlada y factores que la afectan
La primera etapa en un programa de inoculación de MA o ectomicorrizas para usar en agricultura o silvicultura depende de la inoculación más apropiada. En este sentido, la principal opción es el manejo de las poblaciones nativas (17).
Para la obtención de los MA a utilizar, se han desarrollado diversas metodologías tendientes al entendimiento de la ecología y funcionamiento de esta asociación simbiótica, incluye el aislamiento de esporas a partir de suelo, la tinción diferencial de raíces para evidenciar la colonización y metodologías moleculares para la identificación y taxonomía (1, 14).
La micorrización controlada en viveros es una operación hoy en día necesaria para obtener buenos resultados, y para que un programa sea eficaz se necesita utilizar hongos que sean competitivos tanto en vivero, como en el campo. Se ha demostrado la efectividad de la inoculación con micorrizas en numerosos cultivos y regiones (3, 48).
El factor a tener en cuenta es definir las cepas micorrícicas nativas para las inoculaciones de cada especie o cepa, donde se tiene limitaciones ecológicas en las que el comportamiento es el más efectivo, en términos de crecimiento de su hospedante. Si esto no se tiene en cuenta es posible que el hongo inoculado en el vivero sea desplazado en el campo por otros hongos más adaptados al medio (11).
Para una buena evaluación, es básico realizar un muestreo que sea representativo. El mismo debe hacerse a una profundidad entre 0 y 20 cm en forma de «X» o «W». Las muestras tomadas en los diferentes puntos de muestreo seleccionados se deben mezclar y homogenizar completamente y pasadas por un tamiz de 2mm (15). CORPOICA, en la metodología empleada para el estudio de MA, recomienda que la muestra debe secarse a temperatura ambiente hasta obtener el mínimo de humedad, donde posteriormente se toman entre 10 y 100 gr de suelo, según las necesidades de lo que se quiere conocer y se realiza el aislamiento de las esporas presentes siguiendo la metodología de tamizaje y centrifugación en gradiente de sacarosa (15).
De acuerdo a la literatura revisada, lo más importante en el método seleccionado, es estandarizar el procedimiento a las condiciones ambientales donde se va a realizar el trabajo de investigación.
Además como en nuestro medio limitante las estructuras que son transitorias y algunos tejidos presentan dificultades (clareo y tinción), así la posibilidad para identificar taxonómicamente es limitada (15).
El potencial de uso de las micorrizas ha sido relacionado principalmente con cultivos que llevan fase de trasplante como práctica habitual, como acontece en horticultura, fruticultura, programas de reforestación, pero no se recomienda para cultivos arables debido a que los hongos micorrizógenos, no pueden completar su ciclo de vida salvo asociados con la planta con la que forman una mezcla de estructuras del hongo con el substrato de crecimiento y restos de raíces, esto hace que los inóculos estén muy asociados al suelo y tengan una participación importante en el proceso (32,48).
La efectividad de la inoculación dependerá del balance de factores ecofisiológicos en el sistema planta-suelo, lo cual significa que un mismo inóculo puede desencadenar efectos muy variados sobre un cultivo, dependiendo del tipo de manejo agronómico que se realice. Por lo tanto, no se puede generalizar el efecto de un tipo de micorrizas sobre todos los árboles ni sobre todos los ecosistemas. Sin embargo es importante determinar la aplicación del inóculo el cual puede realizarse directamente sobre la planta de una forma homogénea aprovechando el agua de riego. La época de tratamiento suele ser al momento de la siembra o la germinación. La dosis de aplicación puede variar según la especie de hospedante, siendo un valor medio 107 esporas por planta; de acuerdo a consulta a expertos en nuestro medio se recomienda 100 a 150 gr por árbol con una densidad de 50 esporas/gr; se espera que los efectos de la micorrización se empiece a ver a los cinco o seis meses de la germinación.
Según Hadte (21), para determinar la efectividad de la micorriza aplicada se debe determinar y cuantificar la colonización del hongo micorrícico en las raíces y determinar las características del mismo, lo cual se puede visualizar bajo un estereoscopio (40x) por el método de intercepto de líneas cuadráticas, después de que las raíces han sido lavadas (al eliminar los materiales nucleares y citoplasmático), acidificadas y tinturadas en vías específicas.
Consideraciones finales
De acuerdo a la información recopilada, las MA constituyen una alternativa valiosa para disminuir el tiempo de entrada de animales a pastoreo al reducir el tiempo de establecimiento de árboles para los sistemas silvopastoriles. Así mismo, su uso implica una menor aplicación de fertilizantes, riego y pesticidas, lo que permite un sistema de producción más rápido, limpio y eficiente, que aumenta la sostenibilidad de los cultivos, convirtiéndose en una estrategia válida para entregar a los agricultores. En Colombia las micorrizas tienen un gran potencial para contribuir a la solución de múltiples problemas de la agricultura, por las amplias ventajas que ofrecen para condiciones tropicales. Sin embargo, se debe tener en cuenta que existe una multitud de hongos y una dinámica natural del desarrollo de estas micorrizas. Sus efectos no son iguales en todos los árboles ni en todas las condiciones agroclimáticas, debido a las interacciones que se presentan y al tipo de micorriza que actúa. Es importante resaltar la necesidad de profundizar en la investigación sobre el uso de micorrizas nativas o del sitio, lo que implica conocer más sobre la especificidad hongo-planta, buscando aislar cepas específicas que permitan potencializar el crecimiento y productividad de los árboles en determinados agroecosistemas y evaluar las interacciones que se generan.
Finalmente, se debe considerar que las constituyéndose así en una alternativa valiosa para micorrizas pueden ser utilizadas en la agricultura solucionar problemas de micropropagación, en forma de bioprotectores y biorreguladores, en aclimatación y nutrición de diferentes especies de vivero o durante el enraizamiento de vitroplantas, importancia en la agricultura.
Summary
Importance of mycorrhizae fungi handling in the trees establishment in silvopastoral systems
Silvopastoral systems have been reported as an effective alternative to tropical livestock. However, it’s implementation by farmers has not been according to the investigation. One cause of the slow implementation is the large period that one plot can not be used after plantation. That situation has suggested the use of some alternatives as microorganisms that permit to stimulate the trees growing. This document it is a review of the function, importance, kinds and effects of mycorrhizal fungi in forage trees. It permits to consider that in Colombia the mycorrhizal fungi have a big potential to lead solutions of several agricultural problems. Besides, it is important to increase the investigation about the native mycorrhizae use and to get specific strains in order to increase the growing and productivity of agrosystems.
Key words: livestock, microorganisms, mycorrhizal fungi, soil, sustainability.
Referencias
1. Abbott LK, Gazey C. An ecological view of the formation of VA mycorrhizas. Plant and soil 1994; 159: 69 - 78. [ Links ]
2. Alarcón A, Ferrera-Cerrato R. Dinámica de colonización y efecto de hongos endomicorrícicos sobre el crecimiento de Casuarina equisetifolia L. In: J. Pérez-Moreno y R. Ferrera- Cerrato (eds.). Nuevos horizontes en agricultura: Agroecología y desarrollo sustentable. Colegio de Postgraduados en Ciencias Agrícolas, Montecillo, México. 1996; pp. 298-302 [ Links ]
3. Barea JM. Las micorrizas arbusculares componente clave en la productividad y estabilidad de agroecosistemas. Departamento de Microbiología del Suelo y Sistemas Simbióticos, Estación Experimental del Zaidín, Granada, España, 2003, 50 p. [ Links ]
4. Barrera N, Gómez J, Daniel E, Mejía L. Nuevas investigaciones para un adecuado manejo del chachafruto Erythrina edulis especie para la alimentación del hombre y de los animales domésticos. VI Seminario Internacional sobre Sistemas Agropecuarios Sostenibles. Cipav, Buga, 1.999. [ Links ]
5. Bolan, NS. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Journal Plant and soil. New Zealand 1991; Vol 134: 189 - 207. [ Links ]
6. Bolleta A, Rodríguez C. Efecto de la inoculación conjunta bacteria-micorriza sobre los componentes del rendimiento de trigo bajo siembra directa. Copyright INTA, Rivadavia, Intranet Instituto Nacional de Tecnología Agropecuaria 2002. [ Links ]
7. Bonilla R, Roncallo B, García B, Jimeno J. Utilización de hongos micorrizógenos en la producción agrícola. Boletín de Investigación. Corpoica, Valledupar, 2002. [ Links ]
8. Bonilla R, García B, Roncallo A, Jimeno J. Producción y descomposición de hojarasca en un bosque nativo, sistema silvopastoril y monocultivo de gramíneas en Codazzi, Cesar. Curso Nacional de Pastos y Forrajes. Medellín, Octubre 2003. [ Links ]
9. Bowen GD. The ecology of ectomycorrhiza formation and functioning. Plant and soil 1994; 159: 61 - 67. [ Links ]
10. Cárdenas G. Recovering Paradise: Making asturelands Productive for People and Biodiversity. Naranjo L. G. (Editors). First international workshop on Bird Conservation in Livestock Production Systems. American Bird Conservancy, CIPAV. Virginia, USA, 2000, 23 p [ Links ]
11. Carrillo R, Godoy R, Peredo H. Simbiosis micorrícica en comunidades boscosas del Valle Central en el sur de Chile. Revista Bosque. Valdivia, Chile 1992; Vol. 13 (2) 57 - 67. [ Links ]
12. Castellano M., Bougher N. Consideration of the taxonomy and biodiversity of Australian ectomicorrhizal fungi. Plant and soil 1994; 159: 37 - 46. [ Links ]
13. Colozzi A, Jurandy E, Cardoso N, Gomes S. Micorrizas arbusculares no agrossistema cafeeiro e adubacao verde com leguminosas. 14vo Congreso Latinoamericano de la Ciencia del Suelo. 1997. [ Links ]
14. Corredor, G. Micorrizas arbusculares: Aplicación para el manejo sostenible de los agroecosistemas. Programa Nacional De Recursos Biofísicos, Corpoica, Bogotá, 2003, pp. 12 - 17. [ Links ]
15. Corredor G, Otálvaro D, Álvarez C. Metodologías empleadas para el estudio de micorrizas arbusculares. Programa Nacional de Recursos Biofísicos , Corpoica, Bogotá, 2003, 45 p. [ Links ]
16. Crespo G, S Fraga. Nota técnica acerca del aporte de hojarasca y nutrientes del suelo por las especies Cajanus cajan (L.) Millsp y Albizia lebbeck (L.) Benth en sistemas silvopastoriles. 12 p. [ Links ]
17. Dodd J.C, Thomson BD. The screening and selection of inoculant arbuscular-mycorrhizal and ectomycorrhizal fungi. Plant and soil 1994; 159: 149 - 158. [ Links ]
18. Fitter AH, Garbaye J. Interactions between mycorrhizal fungi and other soil organisms. Plant and soil 1994; 159: 123 - 133. [ Links ]
19. Francis R, Read DJ. The contributions of mycorrhizal fungi to the determination of plant community structure. Plant and soil 1994; 159, pp. 11 - 25. [ Links ]
20. Gómez L. Los Biofertilizantes. Revista Despertar Lechero, COLANTA, Medellín, 2002, Vol. 21pp. 35 - 45 [ Links ]
21. Habte M, Osorio NW. Arbuscular micorrizas: producing and applyng arbuscular mycorrhizal inoculum. Collage of tropical agriculture and human resources. University of Hawaii at Manoa, 2001, pp. 15 - 23. [ Links ]
22. The soil food wed: tuning in to the world beneath our feet. Revist Acres, A voice for eco-agriculture. USA 2003, Vol: 30, Nro. 4. [ Links ]
23. Infante JM. Las micorrizas 2003 http://www.tri-ton.cl/index.php.pagina=faq [ Links ]
24. Kaur B, Gupta SR, Singh G. Bioamelioration of a sodic soil by silvopastoral systems in northwestern India. Department of Botany, Kurukshetra University, Agroforestry systems 1999; 46: 150 - 175. [ Links ]
25. Khanizadeh S, Hamel C. Effect of three vesicular-arbuscular mycorrhizae species and phosphorus on reproductive and vegetative growth of three strawberry cultivars. Journal of plant nutrition, Canada, 1995; 18 (6): 1073 - 1.079. [ Links ]
26. Londoño C, Estrada J. Evaluación productiva de la micorriza Glomus manihotis en el pasto Axonopus Scoparius. Universidad Nacional de Colombia. Tesis, Facultad de Ciencias Agropecuarias. Medellín, 1991, pp. 10 - 24. [ Links ]
27. Mahecha L. El silvopastoreo: una alternativa de producción que disminuye el impacto ambiental de la ganadería bovina. Revista Colombiana de Ciencias Pecuarias 2002; Vol. 16. [ Links ]
28. Maldonado JM, Ramírez GA. 1997. Efecto de la inoculación con hongos micorrizógenos en almácigos de café (coffea arábiga) Variedad Colombia. Universidad Nacional de Colombia. Tesis, Facultad de Ciencias Agropecuarias, Medellín, 1997, pp. 3 - 83. [ Links ]
29. Marschner H., Dell B. Nutrient uptake in mycorrhizal symbiosis. Plant and soil 1994; 159: 89 - 102. [ Links ]
30. Mateus H. Rehabilitación y manejo de praderas degradadas. Curso Nacional de Pastos y Forrajes. Medellín, 2003; pp. 20 - 35. [ Links ]
31. Miller M, Mcgonigle T, Addy H. An economic approach to evaluate the role of mycrrhizas in managed ecosystems. Plant and soil 1994; 159: 27-35. [ Links ]
32. Miyasaka S, Habte M. Plant mecanisms and mycorrhizal symbiosis to increase phosphorus uptake efficiency. Journal Series Nro. 4468, College of tropical Agriculture and Human resources, Hawaii, Honolulu, 2003; pp. 1101 - 1133. [ Links ]
33. Morton J. Evolutionary relationships among arbuscular mycorrhizal fungi in the endogonaceae. Mycologia, 1990; 82 (2), pp. 192 - 207. [ Links ]
34. Morton J., Bentivenga S.Levels of diversity in endomycorrhizal fungi (Glomales, Zygomycetes) and their role in defining taxonomic and non-taxonomic groups. Plant and soil 1994; 159: 47 - 59. [ Links ]
35. Muhammad Ibrahim. Sistemas Silvopastoriles en América Central: Experiencias de CATIE. Costa Rica, 1999, 12 p. www.cipav.org.co/redagrofor/memorias99/IbrahimM.htm [ Links ]
36. Murgueitio E, Calle Z. Diversidad biológica en sistemas de ganadería bovina en Colombia. Fundación Cipav, AA 20591, Cali, Colombia, 2002, 35 p. [ Links ]
37. Murgueitio RE. Sistemas Agroforestales para la Producción Ganadera en Colombia. Fundación CIPAV, Cali, Colombia, 1999, 22 p. www.cipav.org.co/redagrofor/memorias99/Murgueit.htm [ Links ]
38. Murgueitio RE, Molina C. El silvopastoreo una alternativa para la ganadería sostenible y competitiva. Experiencias El Hatico. Conferencia lanzamiento red Antioqueña Agroforestería. Medellín, 2001. [ Links ]
39. Pate JS. The mycorrhizal association: just one of many nutrient acquiring specializations in natural ecosystems. Plant and soil 1994; 159, pp. 1 - 10. [ Links ]
40. Powlson D, Hirsch P. Managing soil biological processes for sustainable agriculture and environmental protection. 14vo Congreso Latinoamericano de la Ciencia del Suelo 1997, pp. 118. [ Links ]
41. Raddattz E. Micorriza: el abono vivo Campo & Agro. Zamorano, España, 2002, 15 p. [ Links ]
42. Ramírez A, Otálvaro D, Álvarez C, et al. Efectos de microorganismos rizosféricos sobre la absorción de fosfato y el crecimiento de Leucaena en un andisol. Revista Suelos Ecuatoriales 2001; Vol. 31- 2. [ Links ]
43. Reyes JC, Ferrera-Cerrato R, Alarcón A. Endomicorriza vascular, bacterias y vermicomposta en plántulas de aguacate en vivero. Memoria Fundación Salvador Sánchez Colin CICTAMEX S.C. Coatepec Harinas, México, 1998, pp. 12 - 22. [ Links ]
44. Rodríguez I, Crespo G, Rodríguez C, et al. Comportamiento de la macrofauna del suelo en pastizales con gramíneas naturales puras o intercaladas con Leucaena leucocephala para la ceba de toros. Revista Cubana de ciencia Agrícola 2002; Tomo 36. Nro. 2, pp. 181 -185. [ Links ]
45. Rodríguez I, Crespo G, Rodríguez C. Biomasa y diversidad de la macrofauna del suelo en diferentes pastizales. Revista cubana de ciencia Agrícola 2002; Tomo 36, Nro. 4, p 403 - 408. [ Links ]
46. Sadanandan N, Brown A. Management of soil, Nutrients and water in tropical plantation forests. CSIRO Canberra Australia. ACIAR, 1997, 571 p. [ Links ]
47. Sánchez de PM. Endomicorrizas en agroecosistemas colombianos. Universidad Nacional de Colombia. Sede Palmira, 1999, 227 p. [ Links ]
48. Sempere F, Santamarina P. La Aplicación de las Micorrizas. Revista "Agrícola Vergel" 2001; Nº 232 de Abril, pp. 198-201. [ Links ]
49. Sylvia D. Fuhrmann J, Hartel P, Zuberer D. Principles and applications of soil microbiology. Ed. Prentice Hall Inc. New Jersey, USA, 1999, 550 p. [ Links ]
50. Velasco A, Zambrano J. Mejoramiento del Suelo por Acacia Mangium en un sistema silvopastoril con Brachiaria Humidicola. CATIE, Costa Rica, 2000, 15 p. [ Links ]
51. Vilar A, Siquiera J, Layola J, Rocha A. Micorrizas arbusculares. Boletín Pesquisa, EMBRAPA, Brasil, 2000, Nro. 17, 17 p. [ Links ]
52. Walker C. Systematics and taxonomy of the arbuscular endomycorrhizal fungi (Glomales) - a possible way forward. Agronomie 1992; 12, pp. 887 - 897 [ Links ]
53. Young A. Agroforestry for soil management. Second edition. CAB International, Wallingford, UK, 1997, 350 p. [ Links ]

