










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.19 no.1 Medellín Jan./Mar. 2006
Relación entre el flujo de proteína microbiana hacia el duodeno y algunos parámetros metabólicos y productivos en vacas lactantes de un hato lechero del Oriente Antioqueño.
Sandra L Rueda1, Zoot; Lorena M Taborda1 Zoot; Héctor J Correa2, Zoot, MSc.
1Zootecnista, Universidad Nacional de Colombia, sede Medellín.
2Profesor de Departamento de Producción Animal, Universidad Nacional de Colombia, sede Medellín. AA 1779.
(Recibido: 9 agosto, 2004; aceptado: 10 octubre, 2005)
Resumen
Con la finalidad de estimar el flujo de proteína microbial hacia el duodeno (PMFD) en vacas lactantes a partir de la excreción urinaria de alantoína y establecer su relación con algunos parámetros del comportamiento productivo, se utilizaron ocho vacas (4 Holstein y 4 BON x Holstein) en diferente estado de lactancia (55 a 361 días en lactancia) que permanecieron bajo estabulación durante nueve días consumiendo pasto kikuyo (Pennisetum clandestinum), concentrado comercial y sal mineralizada.
Durante los tres últimos días se registró la producción de leche (PL) y se tomaron muestras para analizar la concentración de proteína cruda (PCL) y nitrógeno ureico (NUL). En muestras del pasto y del concentrado se determinó la materia seca (MS), la proteína cruda (PC), las cenizas (Cen), el extracto etéreo (EE), la fibra en detergente neutro (FDN), el nitrógeno insoluble en detergente neutro (PCIDN) y los carbohidratos no estructurales (CNE) y se estimó la concentración de nutrientes digestibles totales (NDT). Se determinó el consumo de materia seca (CMS) (11.63 ± 1.8 kg/vaca/d), el nitrógeno ingerido total (NIT) (249.9 ± 57.7 gr/vaca/d), la proteína degradable en rumen (PDR) del pasto y del concentrado mediante la técnica in situ, y la relación CNE: PDR (1.1 ± 0.15 gr: gr). Los dos últimos días se recolectó la orina excretada por cada vaca (OR) donde se determinó la concentración de nitrógeno (NO) (23.08 ± 5.64 kg/vaca/d) y de alantoina (16.9 ± 15.1 g/d) a partir de la cual se estimó la PMFD (482.9 ± 509.7 g/vaca/d). Asimismo, con base en los NDT consumidos se estimó la PMFD (PMFDNRC) (769.80 ± 161.20 g/cow/d). Se establecieron relaciones mediante ecuaciones de regresión lineal entre el NO y la OR (p<0.03), la PMFD con el CMS (p<0.04) y con el NIT (p<0.04), y entre la PL (p<0.05), la PCL (p<0.03) y el NUL (p<0.06) con la PMFD. La relación con CNE: PDR no fue estadísticamente significativa (p> 0.7). Los datos sugieren que la excreción de alantoína en orina se puede usar para estimar la PMFD.
Palabras clave: alantoína, estimación, regresión.
Introducción
Los sistemas intensivos de producción de leche en el departamento de Antioquia se basan en el uso de gramíneas, que como el pasto kikuyo (Pennisetum clandestinum), son manejadas con aplicaciones altas de fertilizantes nitrogenados (13). Esto se debe al efecto positivo que estos fertilizantes generan sobre la producción de biomasa forrajera y, en última instancia, sobre la producción de leche en el hato (35). La fertilización nitrogenada, sin embargo, afecta la calidad de nutricional de los pastos conduciendo al incremento del contenido de nitrógeno (N) en el forraje (28, 35, 44) debido al aumento en el N soluble y el N no proteico (NNP), es decir, en la fracción soluble (fracción a), en detrimento de la fracción potencialmente degradable (fracción b) (35). Estos cambios finalmente se traducen en un alto contenido de proteína degradable en rumen (PDR) que por lo general, es superior al 12% de la MS (17, 40).
Ha sido señalado, sin embargo, que el uso eficiente de la PDR para la síntesis de proteína microbial depende de la concentración de carbohidratos no estructurales (CNE) en la dieta postulándose que debe existir una relación entre los CNE y la PDR que oscile entre 3.2 y 4.0 (19, 31, 41). El contenido de CNE del pasto kikuyo (Pennisetum clandestinum), sin embargo, generalmente en inferior al 12% de la MS (17, 29, 40) lo que aunado al alto contenido de PDR conduce a una relación CNE: PDR inferior a 1.0.
La suplementación con alimentos concentrados no logra corregir el desequilibrio del pasto kikuyo (Pennisetum clandestinum) en cuanto a su relación CNE: PDR (6, 12) así como tampoco el uso de alimentos con alto contenido de CNE y bajo contenido de proteína cruda como la papa (29). Bajo estas condiciones es de esperarse que la síntesis de proteína microbiana en el rumen sea más baja que la que se puede estimar (31) y que su flujo hacia el duodeno sea igualmente baja. Esto trae implicaciones en el uso metabólico del nitrógeno (N) tales como en la síntesis de urea, la concentración de este metabolito en la leche; la pérdida urinaria del N, la producción de leche y de proteínas lácteas (11). Por lo tanto, es de esperarse que existan relaciones entre la proteína microbiana hacia el duodeno (PMFD) y estas variables. Es por ello que el objetivo de este trabajo fue estimar el flujo de PMFD en vacas lactantes alimentadas con pasto kikuyo (Pennisetum clandestinum) y establecer su relación con algunos parámetros metabólicos y productivos.
Materiales y métodos
Se llevó a cabo un experimento en el Centro de Producción Paysandú de la Universidad Nacional de Colombia, localizado en el corregimiento de Santa Elena (Medellín, Antioquia), a 2300 msnm, con una temperatura promedio de 16°C, perteneciente a una zona ecológica de bosque húmedo – Montano Bajo (bh – MB). Para ello se utilizaron cuatro vacas lactantes de las raza Holstein y cuatro F1 (Holstein x Bon) que se encontraban en diferentes momentos de lactancia, pero en iguales condiciones de manejo y de alimentación, consistente en el suministro de pasto kikuyo recién cosechado y un alimento concentrado comercial de acuerdo con el programa de suplementación que se tiene establecido en este hato para cada animal (15) y que está en función del grupo genético, el número de partos, los días en lactancia (DEL) y la cantidad de leche producida (PL) y que se incorporan en dos ecuaciones:
1. Si el grupo genético es Holstein, entonces (1):
PL/(3.074066+0.00385772*DEL + 0.1*Número de partos)
2. Si grupo genético es F1, entonces (2):
(PL/(3.074066+0.00385772*DEL + 0.1*Número de partos) – 0.7
Las vacas seleccionadas hacían parte de un experimento en el que el objetivo fue estimar el balance de nitrógeno en dos grupos genéticos (16) empleando datos pareados y del cual se tomó la información y las muestras para ejecutar el presente experimento. Estas vacas se estabularon durante nueve días de los cuales los siete primeros fueron de adaptación a las condiciones de manejo y los tres últimos de recolección de muestras. El piso del establo fue cubierto con una capa de viruta seca, la cual fue reemplazada en la medida que se contaminaba con heces y orina. Durante todo el periodo experimental se cosechó el pasto kikuyo dos veces al día proveniente de un mismo potrero y que tenía aproximadamente 40 días de rebrote.
Inmediatamente después de la cosecha, el pasto fue pesado y empacado en bultos de 10 kilogramos que eran suministrados a los animales en estado fresco registrando el número de bultos por día y la cantidad de material rechazado cada 24 horas. El suplemento concentrado asignado para cada animal fue suministrado dos veces al día al momento del ordeño.
Con el alimento concentrado, se mezclaron 60 gr de sal mineralizada (con aproximadamente 14% de Ca y 8% de P) y se garantizó el suministro permanente de agua potable. Diariamente se recolectaron al azar muestras tanto del forraje como del alimento concentrado, las cuales se conservaron refrigeradas y al final de los diez días se mezclaron y se tomó una muestra final, la cual se secó a 60°C durante 48 horas y se conservó hasta los análisis correspondientes.
Tanto en las muestras de pasto como del suplemento alimenticio se determinó el contenido de materia seca (MS), las cenizas (Cen), el extracto etéreo (EE) y laproteína cruda (PC) de acuerdo a los métodos descritos por la AOAC (4) en tanto que la determinación del contenido de fibra en detergente neutro (FDN) y de fibra en detergente ácido (FDA) se llevó a cabo por los métodos descritos por Van Soest y Robertson (43). El contenido de CNE se calculó por diferencia (41). En el residuo de la FDA se determinó el contenido de PC para determinar la PC insoluble en el detergenteácido (PCIDA).
Asimismo, durante todo el periodo experimental las vacas conservaron arneses para la sujeción de las bolsas utilizadas en la recolección de orina. Esta recolección se realizó durante los últimos dos días del periodo experimental tomando al azar muestras de orina luego de la medición del volumen excretado, las cuales se llevaron inmediatamente a congelación para evitar su descomposición. Al final del periodo experimental se mezclaron las muestras recolectadas para cada animal tomándose tres submuestras que se congelaron hasta la realización de los análisis correspondientes.
Durante los nueve días que permanecieron los animales en estabulación se registró la producción de leche de los dos ordeños y se tomaron muestras de cada ordeño de los últimos tres días al final de los cuales se mezclaron y se tomaron dos submuestras que se congelaron hasta las determinaciones correspondientes.
En un trabajo paralelo (17) se utilizaron dos vacas canuladas al rumen para la determinación de la degradabilidad de la MS y la PC tanto del forraje como del suplemento concentrado. Estas vacas consumieron el mismo forraje que se suministró a los animales experimentales. Esta prueba de degradabilidad se llevó a cabo de acuerdo a la metodología descrita por Ørskov y McDonald (32) y que consistió en la incubación de tres gramos de muestra (molidas a 1.0 mm) en bolsas de nylon durante siete tiempos de incubación (20 minutos y 2, 4, 8, 24, 48 y 72 horas) con dos bolsas por tiempo y por animal para un total de cuatro bolsas por tiempo y por alimento. Luego de la incubación las bolsas fueron lavadas con agua corriente hasta que esta salió limpia, se secaron a 60oC durante 48 horas, se pesaron y se retiraron los residuos en los que se determinó el contenido de MS y PC (4).
Los parámetros de cinética ruminal de la proteína se estimaron con base en el modelo atribuido a Ørskov y McDonald (32): Y = a + b*(1-exp(-kd*t)) donde Y es la degradación ruminal en el tiempo t, a es la fracción soluble, b es la fracción potencialmente degradable y Kd es la constante de la cinética de degradación ruminal.
El contenido de PDR tanto del pasto como del suplemento alimenticio se estimó mediante la ecuación propuesta por Waldo et al (45): PDR = a + b*Kd/(kd + Kp) donde Kp es la constante de la cinética de pasaje ruminal y las demás variables fueron definidas previamente. Para el cálculo de la PDR se asumió un valor de 0.05/h para la Kp.
En las muestras de leche y de orina se determinó la concentración de PC por el método Kjeldahl (4) y el contenido de nitrógeno ureico mediante el método de la ureasa de Berthelot (Bayer Corp., Diagnostic Division, Elkhart, IN).
En las muestras de orina también se determinó la concentración de alantoína de acuerdo al método descrito por Chen y Gomes (9). Para ello previamente se preparó una curva de calibración con una solución de alantoína estándar (Sigma Chemical, St. Louis MO) a 10, 20, 30, 40, 50 y 60 mg/L de concentración.
Posteriormente las muestras de orina fueron diluidas con agua destilada en dos tiempos: inicialmente se tomó 1.0 ml de orina y se diluyó en 4.0 ml de agua destilada. En un segundo tiempo, se tomó 1.0 ml de la solución anterior y se diluyó en 8.0 ml de agua destilada. Finalmente, 1.0 ml de esta segunda dilución fue sometido al procedimiento descrito por Chen y Gomes (9) y a partir de la curva de calibración, se estimó la concentración de alantoína. Este valor fue multiplicado por 45 debido a la doble dilución que se realizó a la orina (5 y 9 veces, en la primera y segunda dilución, respectivamente). El valor obtenido fue multiplicado, a su vez, por la cantidad total de orina eliminada por los animales por día para estimar la cantidad total de alantoína en orina/día.
Dado que la estimación de la PMFD se basa en la concentración de purinas totales dada en mmol y no en mg (9), los datos obtenido en el paso anterior se dividieron por el factor de conversión correspondiente para la alantoína que es 158 (peso molecular de la alantoína). El dato obtenido se dividió en 0.85 asumiendo que la alantoína corresponde al 85% de las purinas totales (9).
Debido a que parte de las purinas que aparecen en la orina tienen un origen endógeno, es necesario restar de las purinas totales las correspondientes a las endógenas. Para ello se empleó la ecuación propuesta por Osuji et al (33) ya que como lo señalan Chen y Gomes (9), aparentemente los animales en zonas tropicales presentan una excreción endógena de purinas menor que animales en zona templada: y = 0.185 X0.75, donde y son los mmol de purinas endógenas y X es el peso vivo del animal.
De acuerdo al principio en el que se basa la técnica de la alantoína, las purinas absorbidas intestinalmente tienen su origen únicamente en los ácidos nucleicos de los microorganismos absorbidos y además, estas purinas no sufren ningún tipo de metabolismo en el animal y como tales son excretadas en la orina (9).
Por tal razón la estimación de las purinas absorbidas en el intestino se calcularon como la diferencia entre las purinas totales y las endógenas.
Para el cálculo de la proteína microbiana absorbida en duodeno se siguió el procedimiento descrito por Chen y Gomes (9) en el que se asume que la digestibilidad de las purinas microbianas es del 83 %, que la concentración de nitrógeno en las purinas microbianas es de 70 mg/mmol, y que la relación entre el nitrógeno de las purinas y el nitrógeno total de los microorganismos ruminales es de 11.6 : 100 (9).
Finalmente, este valor se multiplicó por 6.25 para obtener la PMFD.
Por otro lado, con base en la ecuación propuesta por el NRC (31) se estimó la proteína microbial sintetizada en rumen (PMicr): PMicr (kg/día) = 0.13*NDT, donde NDT son los nutrientes digestibles totales consumidos/día por cada animal (kg/día). Para ello se calculó el aporte de los NDT a partir de la composición química del forraje y del concentrado que consumieron los animales experimentales basado en el modelo del NRC (31) de acuerdo a los cálculos realizados por Gaitán y Pabón (17). Dado que estos cálculos requieren información sobre el contenido de la PC insoluble en el detergente neutro (PCIDN) y de lignina (Lig) de los alimentos, además de los componentes químicos analizados, la PCIDN se estimó por la ecuación propuesta por Weiss et al (46) (PCIDN = (-87.7 +0.33*(%PC/10) + 0.143*(%FDN/10))/10) en tanto que se asumió un valor de 4.0% de Lig para el pasto kikuyo y un valor de 2% para el alimento concentrado.
La PMicr estimada por este procedimiento se multiplicó por 0.8 para obtener el valor estimado de PMFD (PMFDNRC) asumiendo que el 80% de la PMicr fue digestible y que esta es la que fluye al duodeno para hacer parte de la proteína metabolizable (31).
La PMFD estimada mediante la técnica de la alantoína en la orina, se relacionó con el consumo de materia seca total (CMS), el nitrógeno consumido total (NIT), la relación CNE: PDR, la producción de leche (PL), la PC de la leche (PCL), y el nitrógeno ureico en leche (NUL), la relación entre el nitrógeno excretado en la orina (NO) y la producción de orina (OR), así como la PMFDNRC y la PMFD mediante ecuaciones de regresión lineal para lo cual se empleó el PROC REG del programa estadístico SAS (37).
Resultados
La composición química del pasto kikuyo y del alimento comercial utilizado en el primer experimento se presenta en la tabla 1 donde se puede apreciar que el pasto kikuyo presentó un contenido relativamente alto de PC pero muy bajo de CNE. El alimento concentrado comercial, por su parte, presentó una concentración mucho más alta tanto de PC como de CNE que el pasto kikuyo.
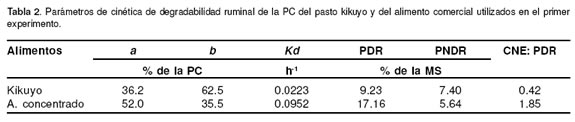
En la tabla 2 se muestran los resultados de la prueba de degradabilidad ruminal del pasto kikuyo y del concentrado comercial utilizados en el primer experimento así como el cálculo de la relación CNE: PDR. Se puede apreciar que aunque la PDR del pasto kikuyo es alta esta fue mucho mayor en el alimento concentrado. La relación CNE: PDR fue muy baja en el pasto kikuyo y aunque mayor en el alimento concentrado comercial, esta estuvo por debajo del valor mínimo recomendado (31).
Los animales experimentales estaban en promedio con 148 DEL variando entre 55 y 361 días, lo quesignifico un CV del 69.4 % (véase Tabla 3). La PL fue menos variable (CV = 34.3%) con un promedio de 13.3 kg/d mientras que la PCL fue muy baja presentando un promedio de 2.18% con una variación media (CV = 16.5%). El peso vivo fue el menos variable (CV = 10.6%) promediando 518 kg.
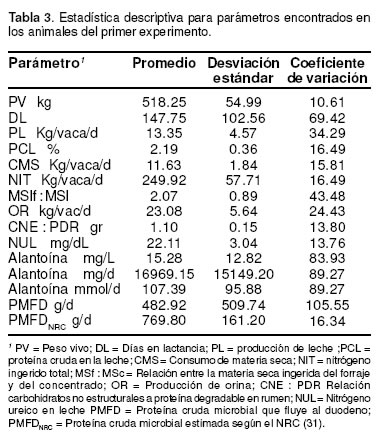
El CMS presentó un promedio de 11.6 kg /vaca/ d con una variación del 15.8% la cual fue muy similar a la del NIT (CV = 16.5%) (véase Tabla 1).
La excreción de orina promedió 23 kg/vaca/d con un CV del 24% mientras que la concentración promedio de NUL fue ligeramente alta (22.11 mg/ dL) con una variación relativamente baja (CV = 13.76%).
En la tabla 3 se aprecia que la PMFD fue la variable que presentó el CV más alto (105.5%) seguida por la variación encontrada en la concentración de alantoína en la orina (CV = 83.9%). El cálculo del FPMD a partir del consumo de NDT empleando la ecuación del NRC (31) (FPMDNRC), presentó un promedio mucho más alto que el valor estimado mediante la técnica de la alantoína pero con una variación mucho más baja (véase Tabla 3).
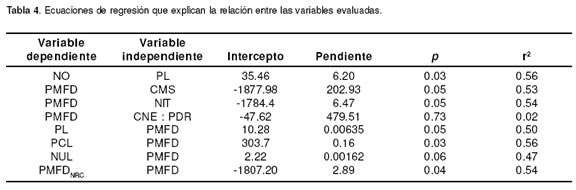
En la tabla 4 se presentan las ecuaciones de regresión que fueron calculadas para explicar la relación entre las variables evaluadas en este trabajo.
Como se puede observar, exceptuando la ecuación de regresión que estima la PMFD a partir de la relación CNE: PDR, todas las demás ecuaciones fueron estadísticamente significativas con un que osciló entre 0.47 y 0.56 y un nivel de probabilidad entre 0.03 y 0.06.
Discusión
La baja concentración de CNE en el pasto kikuyo ha sido referenciada en varios reportes (17, 29, 40) lo que se convierte en el principal limitante para la síntesis de proteína microbial en el rumen y, por lo mismo, en la eficiencia en el uso de la PDR. Ha sido recomendado que la concentración de CNE en la ración de vacas lactantes no sea inferior al 35% de MS (19, 31). En el pasto kikuyo, sin embargo, la concentración de CNE apenas logra superar el 12% de la MS (40) lo que implica la necesidad de suplementar a los animales con alimentos que contengan altas concentraciones de CNE. Aunque el contenido de CNE del alimento concentrado comercial utilizado fue más alto que el del pasto kikuyo, esta concentración, sin embargo, fue inferior al mínimo requerido por vacas en producción que según el NRC (31) debe ser de 36% de la MS.
Debido a esto y al hecho que este alimento representó solamente una parte de la ración, la concentración final de CNE de la ración fue mucho menor correspondiendo al 13.65% de la MS (datos no mostrados). Lo anterior indica lo inadecuado que resulta ser este tipo de alimentos comerciales lo cual ha sido señalado por otros autores (3, 29).
Debido a la baja concentración de CNE y a la alta concentración de la PDR del pasto, la relación CNE: PDR fue muy baja. En el alimento comercial, a pesar de presentar una concentración más alta de CNE que el pasto, debido a su alto contenido de PDR, la relación CNE: PDR fue menor a la recomendada para vacas lactantes (19, 31, 41), lo mismo que la de la ración diaria.
La variación observada en las variables evaluadas en los animales experimentales fue debida a que, como se señaló previamente, este experimento se derivó de otro en el que el objetivo era estimar el balance de nitrógeno en dos grupos genéticos (15) empleando datos pareados en el que hubo una alta variación en los DEL y, por lo mismo, en la PDL, y dado que ambos parámetros afectan el comportamiento ingestivo de los animales (31), las demás variables relacionadas también se vieron afectadas por esta variabilidad.
La excreción de orina observada en este trabajo fue aparentemente más alta de lo esperado. Así, Holter y Urban (23) hallaron que la excreción de agua en la orina en vacas holstein lactantes fue de 14.1 kg/d pero que fue bastante variable oscilando entre 4.7 y 36.4 kg/d para vacas con una producción de leche de 34.6 kg/d, y que correspondió a una excreción del 15 al 20% del agua consumida. Al aplicar la ecuación propuesta por Adams y Sharpe (2) para estimar el consumo total de agua (CTA) en vacas lactantes (CTA kg/day) = (4 x CMS) + kg de LCG + 25.6, donde LCG es la leche corregida al 4% de grasa), utilizando los datos de las vacas experimentales (la leche presentó un contenido de grasa del 3.4%) se obtuvo un consumo de 84 kg/día, que al multiplicar por el valor más alto de excreción de agua por orina señalado por Holter y Urban (23) (20% del agua consumida), se obtiene una excreción máxima estimada de 16.8 kg/ vac/d, valor mucho menor que el hallado en este trabajo (23.08 kg/vaca/d). Esta mayor excreción de orina podría estar explicada en las características de la dieta, particularmente en lo que respecta al contenido de proteína ya que Holter y Urban (23) afirman que dietas con altos contenidos de proteína podrían incrementar la excreción de orina. En el trabajo de estos autores la concentración de PC de las dietas fue de 16.0% en promedio en tanto que la del presente trabajo estuvo entre 21.3 y 22.8 %. Murphy (30), haciendo un estudio sobre el metabolismo del agua en vacas lactantes encontró una relación positiva entre la excreción urinaria del agua y la disponibilidad de agua para el consumo, la cantidad de agua absorbida intestinalmente y la concentración de nitrógeno en la orina.
Efectivamente, en el presente trabajo se encontró una relación positiva y significativa (p<0.03) entre la excreción total de orina en las vacas y la concentración de nitrógeno en la misma (véase Tabla 4).
Adicionalmente, otro factor que habría incidido en la alta excreción de orina es la alta humedad del forraje ofrecido ya que como lo indican Paquay et al (34), las dietas con alto contenido de humedad tienden a incrementar la excreción de agua por orina.
La concentración de NUL estuvo ligeramente más alta que el valor reportado por Montoya et al (29) para vacas holstein lactantes en el municipio de San Pedro de los Milagros, pero muy similar a los valores reportados por Abreu y Petri (1) para hatos lecheros en la Sabana de Bogotá (21.75 mg/dL). Estos últimos autores reportaron un valor mucho más bajo para hatos en el departamento de Boyacá (13.8 mg/dL) argumentando que estos son un reflejo del menor contenido de proteína cruda de la dieta. Este contenido de proteína en los pastos en Boyacá, sin embargo, fue muy similar al encontrado en el presente trabajo (20.6 %) lo que estaría indicando que existen otros factores adicionales que estarían explicando estas diferencias. Uno de estos sería el nivel de producción de leche ya que en el trabajo realizadoen la Sabana de Bogotá el promedio de producción de leche fue 24.6 l/vaca/d en tanto que en Boyacá fue de 15.2 l/vaca/d. En general, sin embargo, se puede afirmar, como lo señala Carulla (8), que los niveles de urea en leche de los sistemas de producción de lechería especializada en nuestro medio son más altos que los recomendados (15 mg/dL). Hall (21) afirma que altos niveles de urea (> 16 mg/dl) indican una sobrealimentación de proteína o una baja relación entre la energía de los carbohidratos y la proteína.
En la tabla 3 se aprecia que la PMFD fue la variable que presentó el CV más alto (105.5%). Esta variación es de alguna manera reflejo de la variación encontrada en la concentración de alantoína en la orina como se aprecia en la misma tabla 4 (CV = 83.9%). Esta estaría respondiendo a la variación en la concentración de proteína microbiana que fluyó al duodeno en estos animales. Aunque el cálculo de la síntesis de proteína microbiana en rumen a partir del consumo de NDT empleando la ecuación del NRC (31) presentó una variación mucho más baja (véase Tabla 3) y que la ecuación de regresión que estima la PMFD a partir de la PMFDNRC fue estadísticamente significativa (p<0.04) (véase Tabla 4), cabe señalar que el modelo del NRC (31) asume que el suministro de los alimentos que conforman la ración diaria se hace simultáneamente y no de manera separada como sucede bajo el esquema de alimentación que predomina en nuestro medio y que fue el que se aplicó a las vacas experimentales, esto es, suministrar el concentrado aparte del forraje. Si la ecuación del NRC (31) aplicara, se esperaría un mayor flujo de proteína microbiana hacia el duodeno (769.80 gr/d) que la estimada en este trabajo a través de la excreción de alantoína en orina (482.9 gr/d). Huntington (24) ha señalado la necesidad de que exista una sincronía en el momento en que los sustratos que participan en la síntesis de proteína microbial en el rumen, ingresen a este simultáneamente. De no ser así, esta síntesis se verá reducida. Esto fue probablemente lo que se presentó en este trabajo, es decir, una baja eficiencia en el uso de los sustratos para la síntesis de la proteína microbiana. Esto concuerda con los resultados de Montoya et al (29) quienes observaron que la producción de leche y de proteínas lácteas fue más baja en vacas alimentadas bajo el esquema de alimentación tradicional que cuando estas se suplementaron con papa durante el pastoreo. Por el contrario, esta suplementación disminuyó la concentración sanguínea de urea indicando un mejor uso de los sustratos a nivel ruminal.Otra posible causa en la variación encontrada en la concentración de alantoína en la orina pudo ser el haber utilizado dos días de recolección de orina cuando Chen y Gomes (9) recomiendan que este se prolongue al menos por cinco días para reducir la variación. Sin embargo, otros autores han señalado que periodos de tiempo menores pueden ser suficientes hasta el punto de sugerirse que la recolección por un periodo de 24 horas podría ser suficiente (42). Valadares et al (42) desarrollaron una ecuación para estimar la producción total de orina en vacas lactantes a partir de la concentración de creatinina en la orina encontrando que esta estima de manera satisfactoria la excreción de orina y que en vista de que la concentración de creatinina es muy constante a lo largo del día, la recolección de muestras de orina por un periodo de doce horas podría suficiente para estimar la producción total de orina pero que para estimar la producción total de derivados de purinas es necesario recolectar muestras de orina a lo largo de 24 horas.
Adicional a lo anteriormente expuesto, cabe resaltar que la variación en el CMS y en el NIT pudieron haber afectado la variación en la síntesis de proteína microbiana y, por ende, en su flujo hacia el duodeno.
Los datos de Gonda y Lindberg (18) muestran una muy baja variación en la concentración de alantoína en la orina con un coeficiente de variación que osciló entre 4.8% y 9.8% para los tres experimentos reportados, pero, así mismo, la variación en la materia orgánica (MO) y en la proteína consumida fue baja oscilando entre 2.3 % y 3.3% para la MO y entre 3.2 % y 8.4% para la proteína. La variación encontrada en el presente trabajo para la materia seca consumida fue de 15.8% y de 16.5% para el NIT.
Por otro lado, hay que señalar que la eficiencia en la síntesis de proteína microbial depende de la relación entre los sustratos que son aportados tanto por el forraje como por el concentrado. Esta relación se estableció entre el CMS a partir del forraje sobre el CMS a partir del suplemento, indicando que hubo una variación relativamente alta (43.5%) la que posiblemente habría contribuido a una menor síntesis de proteína microbiana en rumen y a su alta variación.
Finalmente, no se pueden descartar errores metodológicos que pudieron haber incidido en esta variación entre los que se destacan, además del tiempo de recolección de orina anteriormente discutido, el hecho de haber trabajo con una técnica en la que la lectura final de la alantoína se hace indirectamente a través de un cromóforo inestable en donde el tiempo de lectura es crítico (9). La baja variabilidad en los datos junto con el coeficiente de determinación alto (0.98) de la ecuación que se obtuvo entre las concentraciones de la alantoína estándar y la densidadóptica, hacen presumir que la manera como se realizaron los análisis en el laboratorio fue correcta y habría incidido muy poco sobre la variación en los datos de alantoína de las muestras de orina evaluadas. De todas maneras es necesario resaltar nuevamente que los animales experimentales no estuvieron sometidos a ningún tipo de control en cuanto a los alimentos ofrecidos para su consumo y que fueron muy heterogéneos en el nivel de producción de leche (CV = 34.3%) y en los días en lactancia (CV = 69.4%).
En la tabla 4 se presentan las ecuaciones de regresión entre la PMFD y las variables con que se relacionó. El coeficiente de determinación (r2) para las ecuaciones que fueron estadísticamente significativas, fue oscilando entre 0.47 y 0.56. Este coeficiente estaría reflejando la variación propia de la PMFD (CV = 105 %) y de las variables con las que se relacionó. Estos r2 no resultan ser tan bajos toda vez que se están relacionando variables que se hallan sujetas a un gran número de factores, de modo que valores de r2 que explican cerca del 50 por ciento de la variación, indican una relación muy importante entre estas. También es necesario tener en cuenta que el número de individuos utilizados para la toma de información fue bajo (n = 8) y que presentaban una alta variabilidad en su comportamiento productivo como se discutió previamente.
La relación entre el CMS y la PMFD fue relación estadísticamente significativa (p<0.04). Esto posiblemente fue debido a que el incremento en el CMS significa mayor disponibilidad de proteína y carbohidratos de la dieta hacia el rumen lo que habría permitido un incremento en el crecimiento microbial.
De igual manera, al aumentar el consumo habría un aumento de la tasa de pasaje de la digesta ruminal, la cual contiene la proteína microbiana, que posteriormente llegará al duodeno. Esto es confirmado por Bahadur (5) quien señaló que la eficiencia en la síntesis de proteína microbial en el rumen se ve incrementada con el aumento en el CMS. Este autor mostró que el pasaje de la proteína microbiana hacia el duodeno se incrementó proporcionalmente con el CMS.
Johnson et al (25), reportaron que existe una relación lineal entre la cantidad de alantoina excretada en orina de novillas y el CMS digestible como producto de una mayor síntesis de ácidos nucleicos de origen microbiano en el rumen. Estudios realizados por Chen et al (10) indican que la eficiencia en el suministro de nitrógeno microbiano fue mayor con un alto CMS.
Estos mismos autores reportaron un incremento lineal en el suministro de proteína microbial con el incremento de CMS en ovejas. Así mismo, Csori (14) reporta que existe una alta correlación entre la cantidad de alimento consumido y los derivados de purinas o nitrógeno microbial. En vacas lecheras el consumo de materia orgánica aparentemente digestible se ha correlacionado positivamente con la excreción renal de derivados de purinas lo que indica un incremento en la síntesis de proteína microbiana a causa de un aumento en el consumo de materia orgánica digestible (18).
La relación entre el NIT y la PMFD también fue estadísticamente significativa (p<0.05). La ecuación indica que al aumentar el NIT, se aumenta la PMFD lo cual pudo ser debido a un incremento de algunos de los sustratos necesarios para el crecimiento microbial: nitrógeno en forma de urea, aminoácidos y amonio, entre otros. Al incrementarse la disponibilidad de dichos compuestos se espera que aumente la síntesis de proteína microbial, debido a que la proteína del forraje es transformada en amonio para luego ser convertida en proteína microbial. La razón de estaúltima transformación esta determinada por la fermentación de la energía. Cabe señalar que el NRC (31) estima el crecimiento microbial en el rumen bajo dos escenarios diferentes: primero, cuando el contenido de energía en la dieta es limitante se estima que el crecimiento microbial corresponde a 130 gr de proteína bruta microbial por cada kilogramo de nutrientes digestibles totales consumidos; segundo, cuando el contenido de proteína es limitante se estima que la síntesis de proteína bruta microbial se produce con una eficiencia del 85%.
En un estudio realizado por Johnson et al (25) sobre el flujo de proteína microbial al duodeno utilizando diferentes dietas, se encontró que las dietas con altas concentraciones de proteína degradable en rumen (PDR) y carbohidratos no estructurales (CNE) tuvieron un mayor flujo de proteína microbial (p<0.03), y las vacas alimentadas con estas dietas tuvieron la mayor excreción de alantoina (p<0.06) en orina.
La ecuación de regresión entre la PMFD y la relación CNE: PDR no fue significativa (p> 0.7) lo que pudo ser debido a que el rango de observaciones de la variable independiente (CNE: PDR) fue demasiado estrecho (0.91 a 1.3) y a que la variabilidad observada en la variable dependiente (PMFD) fue muy alta (CV = 105%). Se ha señalado que la concentración de CNE en las raciones para vacas lactantes debe estar entre 35 y 40% de la MS (19, 41) y que la PDR deberá estar entre 10 y 12.3% de la MS (27). Esto significa que la relación entre CNE y PDR deberá oscilar entre 3.2 y 3.5: 1.0 para optimizar la síntesis de proteína microbiana. Sin embargo bajo las condiciones de alimentación que prevalece en los sistemas intensivos de producción de leche en Antioquia, esta relación se presenta muy estrecha y normalmente es menor a la unidad debido a los bajos contenidos de CNE de los forrajes. Gaitán y Pabón (17) reportaron que en el pasto kikuyo la concentración de CNE no superó el 9.0% como porcentaje de la MS en tanto que la PDR representó el 77% de la PC (21.32%), lo que corresponde a una relación CNE: PDR de 0.54, lo que es muy inferior al ideal planteado para esta relación. Esto significaría que bajo condiciones de alimentación basadas en este tipo de forrajes existiría una gran limitación para la síntesis de proteína microbiana, lo que se agudizaría si se considera que, como se señaló previamente, los suplementos comerciales que se utilizan no corrigen los desbalances presentados por estos forrajes y además se suministran en momentos inadecuados (29).
En vista de esta situación y presumiendo por lo tanto, que el flujo de proteína microbiana al duodeno es mucho menor que el que podría encontrarse en zonas templadas se ha postulado que el aporte de bases púricas por la proteína microbiana sería menor que en zonas templadas e incapaces de cubrir las pérdidas endógenas (36). Osuji et al (33) sugirieron, además, que las pérdidas endógenas con ganados tropicales son menores. Esta fue la razón por la que se seleccionó la corrección planteada por Osuji et al (33) que indica que las pérdidas endógenas de bases púricas corresponden a 0.185 mmol/kg PV0.75 en lugar del valor planteado por Chen y Gomes (9) de 0.385 mmol/kg PV0.75.
La ecuación de regresión que explica la relación entre la PMFD y la PDL fue positiva y significativa (p<0.05) como se esperaba si se considera que la proteína microbiana puede representar entre el 59 y 81% del total de la proteína verdadera que alcanza el duodeno (20). Además, ha sido señalado que en la medida en que el nivel de producción de leche se incrementa la cantidad de proteína metabolizable requerida para cubrir las demandas de aminoácidos se incrementa a una tasa más alta que la tasa de aporte de aminoácidos que haría la proteína microbiana. Esto hace que como proporción de la proteína requerida, la proteína microbiana represente un porcentaje más bajo de la proteína metabolizable, en tanto que la proteína no degradada en rumen (PNDR) representaría un porcentaje más alto (38). Así podría esperarse que en vacas de alta producción la proteína microbiana correspondiera al valor más bajo planteado por Grieve et al (20), lo contrario sucedería en vacas de baja producción. En el presente trabajo los datos de proteína microbiana se expresaron como gramos que fluyen al duodeno y no como proporción de la proteína metabolizable. De allí que la relación hallada sea positiva. De los datos presentados por Brun-Bellut et al (7) se aprecia una aparente relación positiva entre la proteína microbiana y la producción de leche en cabras aunque ellos no analizaron estadísticamente esta relación.
La relación observada entre la PMFD y la PCL fue estadísticamente significativa (p<0.03) y es explicada por la necesidad de aminoácidos absorbidos en el intestino para la síntesis de proteínas lácteas. Sin embargo, es bien conocido que los aminoácidos que alcanzan la glándula mamaria tienen su origen tanto en los que se absorben intestinalmente como en aquellos que se movilizan desde proteínas lábiles (26) y que estos últimos incrementan su participación al inicio de la lactancia cuando se incrementa la movilización tisular. Con la información recolectada en el presente trabajo, no es posible señalar hasta que punto la respuesta en producción de proteínas lácteas fue debida a los aportes que habrían realizado las proteínas movilizadas debido a que no se hizo un seguimiento al peso de los animales durante el periodo experimental o de algún indicador específico de movilización de proteínas tisulares. De todas maneras, el análisis estadístico de la ecuación con la que se estableció la relación entre la PMFD y las proteínas lácteas indica que el 56% de la variación observada en esta última fue debido a la primera. La síntesis de proteína láctea responde en última instancia a la absorción de aminoácidos intestinales cuyos principales orígenes están en la proteína microbiana y la proteína no degradable en rumen. Algunos trabajos muestran una respuesta al incremento de aminoácidos disponibles a nivel intestinal. Es así como Komaragiri y Erdman (26) encontraron una respuesta positiva a la suplementación con PNDR proveniente de harina de pescado y de carne y huesos argumentando que el incremento en la cantidad de aminoácidos que alcanzaron el duodeno a partir de estas fuentes, habría incrementado la disponibilidad de aminoácidos para la síntesis de proteínas lácteas. De manera similar, Wright et al (47) encontraron una respuesta positiva tanto en producción de leche como de proteínas lácteas a la suplementación con PNDR y explican sus resultados en términos del aporte de aminoácidos esenciales que haría esta PNDR al intestino.
Aunque los resultados no son muy consistentes se han adelantado algunos ensayos para incrementar la síntesis de proteína microbiana mediante el uso de algunas estrategias de alimentación. Es así como Montoya et al (29) reportaron un incremento en producción de leche y proteínas lácteas en vacas suplementadas con papa como fuente de CNE basados en el supuesto de que al sincronizar la relación CNE: PDR y el momento del suministro, habría un incremento en síntesis de proteína microbiana y un mayor aprovechamiento de la PDR. Esto aparentemente fue logrado dado que hubo una disminución del nitrógeno ureico tanto en sangre como en leche con esta suplementación lo que indicaría que hubo una mejor utilización del amonio ruminal que se habría traducido en síntesis de proteína microbiana, la que a su vez explicaría parcialmente los incrementos en producción y proteína láctea. Otros autores (22, 39), sin embargo no encontraron respuesta a esta sincronización debido posiblemente a que la diferencia entre los tratamientos no fue tan dramática como la de Montoya et al (29).La relación entre la PMFD y el nitrógeno ureico en leche (NUL) fue positiva y significativa (p<0.06). Aunque aparentemente esto signifique una contradicción con lo que se argumentó para explicar la respuesta en la producción de proteína láctea, es necesario considerar que este fue un trabajo de caracterización del estado nutricional y metabólico de un grupo de animales que no se sometieron a ningún tratamiento nutricional o alimenticio y que en esencia, busca explicar la relación causa - efecto entre eventos que encuentran relacionados. Así, ya se había explicado anteriormente la respuesta positiva entre el CMS y el nitrógeno ingerido con la PMFD, y la relación entre esta última con la producción de leche y de proteínas lácteas. Todas estas relaciones fueron directas y explicadas en términos de sustratos disponibles. En ese mismo sentido al aceptar que a mayor cantidad de nitrógeno ingerido hay una mayor disponibilidad de PDR para la síntesis microbiana, se debe reconocer así mismo que esta PDR no es totalmente utilizada para la síntesis de proteína microbiana y por lo tanto aquella que no se utilice para este fin generará amonio que necesariamente tendrá que ser transformada en urea a nivel hepático. Esta última se ve reflejada como un incremento en la concentración de urea en los fluidos del animal (sangre, orina, leche, saliva) tal y como se encontró en este trabajo.
Los resultados obtenidos en este trabajo mostraron que la proteína que fluyó al duodeno estimada a partir de la técnica de la alantoína en orina presentó una alta variación que pudo ser consecuencia de la asincronía en el momento en que se suministraron los sustratos al rumen, y en la asincronía en la cantidad de sustratos provenientes tanto del forraje como del concentrado. De igual manera, la variación en los días en lactancia y en la producción de leche de los animales xperimentales también pudo contribuir a esta variación. No obstante esto, los resultados también sugieren que la concentración de alantoina en orina puede ser un buen indicador para estimar el flujo de proteína microbial que alcanza el duodeno permitiendo establecer relaciones con diversas variables productivas y metabólicas. Es así como se encontró una relación positiva entre el flujo de proteína microbial que alcanza el duodeno con el consumo de materia seca, el nitrógeno ingerido total, la producción de leche, las proteínas lácteas y el nitrógeno ureico en leche.
El trabajo permitió corroborar la información de la literatura que señala que el flujo de proteína microbiana depende del CMS y del nitrógeno ingerido. Así mismo, que la cantidad de proteína microbiana que fluye al duodeno explica en una alta proporción la producción de leche, de proteínas lácteas y de nitrógeno ureico en la leche.
No se pudo establecer el efecto de la relación CNE: PDR sobre el flujo de proteína microbial al duodeno debido probablemente a que el rango en el que varió la relación CNE: PDR fue demasiado estrecho y la variabilidad en los valores del flujo de proteína microbial al duodeno fue muy alta.
Agradecimientos
Este trabajo fue realizado gracias al apoyo financiero dado por la Dirección Nacional de Investigaciones de la Universidad Nacional, (código DINAIN: DI00C307) y a la ayuda de los trabajadores del Centro de Producción Paysandú.
Summary
Relationship between the microbial protein flow to the duodenum and some metabolic and productive parameters of lactating cows in a dairy herd of the Antioquia east With the purpose to estimate the microbial protein flow to the duodenum (MPFD) in lactating cows from the urinary excretion of allatoin and establish its relationship with some productive parameters, were used eight cows (4 Holstein and 4 BON x Holstein) in different lactation stage (55 to 361 days in lactation) they were kept under confinement conditions for 9 days eating kikuyu grass (Pennisetum clandestinum), commercial mixed food and mineralized salt. During the 3 last days was registered the milk production (MPD) and were collected samples to analyze the crude protein (CPM) and urea nitrogen (MUN) concentration. In samples of the grass and of the commercial mixed food were analyzed the dry matter (DM), crude protein (CP), ash, ether extract (EE), neutral detergent fiber (NDF), insoluble nitrogen in detergent fiber (INDF) and were estimated the non structural carbohydrates (NSC) and the total digestible nutrients (TDN). The dry matter intake (DMI) (11.63 ± 1.8 kg/cow/d) and total nitrogen intake (TNI) (249.9 ± 57.7 gr/cow/d) were determined. Besides, the rumen degradable protein (RDP), both, the grass and the commercial mixed food using the in situ technique, and the NSC: RDP relationship (1.1 ± 0.15 gr: gr) were determined. Urine excretion (UE) of each cow was collected during the last 2 days in which the nitrogen (UN) (23.08 ± 5.64 kg/cow/d) and allantoin concentration (16.9 ± 15.1 g/d) were determined and the MPFD (482.9 ± 509.7 g/cow/d) was estimate from this last. The other hand, using the TDN intake was estimate the MPFD (PMFDNRC) (769.80 ± 161.20 g/cow/d). Linear regression equations between UN and UE (p<0.03), MPFD and DMI (p<0.04), MPFD and TNI (p<0.04), and between the MPD and MPFD (p<0.05), the CPM and MPFD (p<0.03), and the MUN and MPFD (p<0.06), were estimated. The linear regression equation between NSC: RDP relationship and MPFD was not statistically significant (p> 0.7). The results suggested that the urine allantoin excretion can used to estimate the MPFD.
Key words: allantoin, estimation, regression.
Referencias
1. Abreu A, Petri HA. Uso del MUN (Nitrógeno ureico en leche) para diagnosticar balance proteína – energía en la dieta de vacas lecheras Holstein en pastoreo en el altiplano cundiboyacense. Trabajo de grado, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional de Colombia, Santafe de Bogotá, 1998. [ Links ]
2. Adams RS, Sharpe WE. Water Intake and Quality for Dairy Cattle Department of Dairy and Animal Science. Penn State Cooperative Extensión. DAS 95 – 8 1995; URL: http:// www.das.psu.edu/dcn/catnut/PDF /Water.PDF [ Links ]
3. Agudelo MA, Puerta HM. Efecto del esquema de suministro de un suplemento alimenticio comercial sobre algunos parámetros metabólicos y productivos en vacas lactantes. Trabajo de grado, Universidad Nacional de Colombia, Sede Medellín, 2004. 31 p. [ Links ]
4. Association of Official Analytical Chemists. Official Methods of Analysis (13th Ed.). Washington, DC. 1980. [ Links ]
5. Bahadur D. Purine nitrogen index, a possible parameter for rapid feed evaluation in ruminants. MSc Thesis, University of Aberdeen, 1997: 89 p. [ Links ]
6. Bernal LC, Montoya S. Balance energético y proteico en vacas al inicio de la lactancia y su relación con el estado metabólico. Trabajo de grado, Universidad Nacional de Colombia, Sede Medellín, 2004. 75 p. [ Links ]
7. Brun-Bellut J, Lindberg JE, Hadjipanayiotou M.. Protein nutrition and requirements of adult dairy goats. In: Morand-Fehr P, editors. Goat Nutrition. Pudoc, Wageningen; 1991. p 82-93. [ Links ]
8. Carulla J. De la proteína del forraje a la proteína en la leche. Metabolismo del nitrógeno del forraje en la vaca lechera. En: Simposio Internacional sobre la Proteína en la Leche. Medellín, noviembre de 1999. [ Links ]
9. Chen XB, Gómez. Estimation of microbial protein supply to sheep and cattle based on urinary excretion of purine derivatives– an overview of the technical details. Rowell Research Institute,Buckburn, Aberdeen AB2 9SB, UK. 1995. 21p. [ Links ]
10. Chen XB, Ærskov ER, Hovell FD. Excretion of purine derivatives by rumiants: endogenous excretion, differences between cattle and sheep. Br J Nut 1990; 63: 121 – 129. [ Links ]
11. Correa HJ, Cuéllar AE. Aspectos clave del ciclo de la urea con relación al metabolismo energético y proteico en vacas lactantes. Rev Col Cienc Pec 2004; 17(1): 29 – 38. [ Links ]
12. Correa LF, Marín MR. Balance energético y proteico en vacas periparturientas y la relación con su estado metabólico. Trabajo de grado, Universidad Nacional de Colombia, Sede Medellín, 2002. 50 p. [ Links ]
13. Consejo Regional Lácteo. Acuerdo de Competitividad de la Cadena Láctea en Antioquia, 2002. 75 p. [ Links ]
14. Csori MA. Estimación del aporte de nitrógeno microbiano en vacas de lechería mediante la excreción urinaria de los derivados de purinas. Tesis de grado, Universidad Concepción, Chillan, Chile. 1995. [ Links ]
15. Cuellar A. Plan de alimentación centro Paysandú. Universidad Nacional de Colombia, Sede Medellín, 1998. 5 p. [ Links ]
16. Delgado GF. Estudio comparativo del balance de nitrógeno en vacas lactantes de dos grupo genéticos”. Trabajo de grado, Universidad Nacional de Colombia, Sede Medellín. 2003. [ Links ]
17. Gaitán S, Pabón JD. Evaluación energética y proteica de los forrajes utilizados en un hato lechero del oriente antioqueño según el NRC 2001. Trabajo de grado, Universidad Nacional de Colombia, Sede Medellín. Carrera de Zootecnia. 2003. 55 p. [ Links ]
18. Gonda HL, Lindberg JE. Effect of diet on milk allantoin and its relation sheep with urinary allantoin in dairy cows. J Dairy Sci 1997; 80: 364 –373. [ Links ]
19. Grant R. Protein and Carbohydrate Nutrition of High Producing Dairy Cows. University of Nebraska Cooperative Extension 1996; URL: http: //ianrpubs.unl. edu/dairy/ g1027.htm [ Links ]
20. Grieve DG, Wheeler EE, Yu Y, McCleod GK. Effects of dry or ensiled feeds and protein percent on milk production and nitrogen utilization by lactating cows. J Dairy Sci 1980; 63: 1282 - 1290. [ Links ]
21. Hall MB. Strinking a balance: protein feeding and performance. 35th Annual Florida Dairy Production Conference, Gainesville, FL, 1998. [ Links ]
22. Henning PH, Steyn DG, Meissner HH. Effect of synchronization of energy and nitrogen supply on ruminal characteristics and microbial growth. J Anim Sci 1993; 71:2516–2528. [ Links ]
23. Holter JB, Urban WE. Water partitioning and intake prediction in dry and lactating Holstein cows. J Dairy Sci 1992; 75: 1472 – 1479. [ Links ]
24. Huntington G. Starch utilization by ruminants: from basic to the bunk. J Anim Sci 1997; 75: 852 – 567. [ Links ]
25. Johnson LM, Harrison JH, Riley RE. Estimation of the flow of microbial nitrogen to the duodenum using urinary uric acid or allantoin. J Dairy Sci 1998; 81: 2408 – 2420. [ Links ]
26. Komaragiri VS, Erdman AR. Factor affecting body tissu mobilization in early lactation dairy cows. 1. Effect of dietary protein on metabolization of body fat and protein. J Dairy Sci 1997; 80: 929-937. [ Links ]
27. Mahanna B. Dairy Cow Nutritional Guidelines - Part I. Pioneer Hi-Bred International, Inc 1997; URL: http://www.pioneer.com/usa/nutrition/ vettext1.htm [ Links ]
28. Messman MA, Weiss WP, Erickson DO. Effects of nitrogen fertilization and maturity of bromegrass on nitrogen and amino acids utilization by cows. J Anim Sci 1992; 70: 566 - 575. [ Links ]
29. Montoya NF, Pino ID, Correa HJ. Evaluación de la suplementación con papa (Solanum tuberosum) a vacas Holstein lactantes. Rev Col Cienc Pec 2004; 17(3): 241-249 [ Links ]
30. Murphy MR.. Water metabolism of dairy cattle. In: Symposium: Nutritional factors affecting animal water and waste quality. J Dairy Sci 1992; 75: 326 – 333. [ Links ]
31. National Research Council. The nutrient requirement of dairy cattle. Seventh edition. National Academy Press, Washington, D C 2001. 381 p. [ Links ]
32. Ærskov ER, McDonald I. The estimation of protein degradation in the rumen from incubation measurements weighted according to rate of passage. J Agric Sci 1979; 92: 449-503. [ Links ]
33. Osuji PO, Khalili H, Umunna NN. The effects of cottonseed cake with or without molasses on feed utilization, purine excretion and milk production of Boran (Bos indicus) cows fed a mixture of wheat and oat straw. Trop Agric Trinidad 1995; 72: 63-69. [ Links ]
34. Paquay R, De Baere R, Lousse A. Statistical research on the fate of water in the adult cow. 2. The lactating cow. J Agric Sci (Cambridge) 1970 ; 75: 251. [ Links ]
35. Rodríguez D. Caracterización de la respuesta a la fertilización en producción y calidad forrajera en los valles de Chiquinquirá y Simijaca (Estudio de caso). Trabajo de grado, Universidad Nacional de Colombia, sede Bogotá, 1999. [ Links ]
36. Sandoval C, Herrera GF. Estimación de la síntesis de proteína microbial en rumiantes a través de la medición de los derivados de purina en orina. Rev Biomed 1999; 104: 241-251. [ Links ]
37. SAS. SAS User‘s Guide: Statistics (Version 8.0 Ed.) SAS Inst. Inc., Cary, NC. 1999. [ Links ]
38. Santos, AP, Santos JEP, Theurer CB, Huber JT. Effects of rumen-undegradable protein on dairy cow performance: A 12- year literature review. J Dairy Sci 1998; 81: 3183-3213. [ Links ]
39. Sinclair LA, Garnsworthy PC, Newbold JR, Buttery PJ. Effect of synchronizing the rate of dietary energy and nitrogen release on rumen fermentation and microbial protein syntesis in sheep. J Agric Sci (Cambridge) 1993; 120: 251-263. [ Links ]
40. Soto C, Valencia A, Galvis R, Correa C. Efecto de la edad de corte y del nivel de fertilización nitrogenada sobre el valor energético y proteico del pasto kikuyo (Pennisetum clandestinum). Rev Col de Cienc Pec (En publicación). [ Links ]
41. Stokes S. Balancing carbohydrates for optimal rumen function and animal health. Western Canadian Dairy Seminar 1997; URL: http://www.wcds.afns. ualberta.ca/Proceedings/1997/ ch06-97.htm [ Links ]
42. Valadares RFD, Broderick GA, Valadares Filho SC, Clayton MK. Effect of replacing alfalfa silage with high moisture corn on ruminal protein synthesis estimated from excretion of total purine derivatives. J Dairy Sci 1999; 82: 2686 – 2696. [ Links ]
43. Van Soest PJ, Robertson JB. Analysis of forage and fibrous foods. A laboratory manual for animal science. Cornell University, NY. 1985. 503 p. [ Links ]
44. Van Vuuren A, Tamminga S, Ketelaar R. In sacco degradation of organic matter and crude protein of fresh grass (Lolium perenne) in the rumen of grazing dairy cows. J Agric Sci (Cambridge) 1991; 116: 429 - 436. [ Links ]
45. Waldo DR, Smith LW, Cox EL. Model of cellulose disappearance from the rumen. J Dairy Sci 1972; 55: 125-129. [ Links ]
46. Weiss WP, Conrad HR, St Pierre NR. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim Feed Sci And Technol 1992; 39: 95-110. [ Links ]
47. Wrigt TC, Moscardini S, Luimes PH, Susmel P, McBride BW. Effects of rumen-undegradable protein and feed intake on nitrogen balance and milk protein production in dairy cows. J Dairy Sci 1998; 81: 784-793. [ Links ]

