










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.19 no.4 Medellín Oct./Dec. 2006
El espermatozoide, desde la eyaculación hasta la fertilización
Martha Olivera1,2, MV, Dr Sci Agr; Tatiana Ruiz1,2, MV, Ms, PhD; Ariel Tarazona1,2,Z,Ms; Carlos Giraldo1,2,MV, Ms.
1Fisiología y Biotecnología de la Reproducción, 2Facultad de Ciencias Agrarias, Universidad de Antioquia, A.A 1226, Medellín, Colombia.
molivera@catios.udea.edu.co
(Recibido: 23 de junio, 2006; aceptado: 14 de septiembre, 2006)
Resumen
En los últimos años se ha hecho un inmenso progreso en el entendimiento de los mecanismos moleculares involucrados en la maduración del gameto masculino y su tránsito desde la gónada hasta la fertilización del oocito. A lo largo de este trayecto el espermatozoide modifica su morfología y sus componentes moleculares especialmente, y además ocurren procesos que conducen a la activación para la entrada al oocito, para activar a su vez los mecanismos que conducen a la formación del zigoto. Este artículo presenta, a partir de la interpretación de la literatura actual un modelo de los eventos que se suceden a partir de la eyaculación hasta la fertilización, con énfasis en los mecanismos celulares y moleculares conocidos, y señala algunos vacíos de información aún existentes.
Palabras clave: activación, adhesión, capacitación, fusión, reacción acrosomal.
Generalidades sobre la producción y anatomía espermática en mamíferos
La célula germinal masculina se produce en la gónada (testículo) mediante un proceso permanente de división de las células germinales o espermatogonias. El proceso de división meiótica denominado espermatogénesis está controlado hormonalmente por el eje hipófisis-hipotálamo-gónada. A partir de cada espermatogonia se producen cuatro espermatocitos haploides que permanecen unidos entre sí por puentes citoplasmáticos y a la vez están en comunicación con la célula nutricia o de Sertolli; estas últimas, a partir de moléculas señalizadoras, inducen el proceso denominado espermiogénesis o metamorfosis que convierte las espermátides en espermatozoides. Se reconocen cuatro fases características en esta transformación: la fase de Golgi, la de capuchón, la acrosomal y la de maduración.
En la fase de Golgi, la organela del mismo nombre se acerca al núcleo, desprende vesículas que se le sobreponen y poco a poco se unen para convertirse en la vesícula acrosomal que se localiza en la parte apical del núcleo (véase Figura 1). Los centríolos situados en forma de T, muy cercanos al aparato de Golgi, van migrando hacia lo que será la base del núcleo. El centríolo proximal se sitúa en la parte basal del núcleo y, a partir del centríolo distal crece el axonema (véase Figura 1) conformado por dos microtúbulos centrales y 9 pares de microtúbulos periféricos. En la fase de capuchón, la vesícula acrosomal se aplana formando una verdadera capucha sobre el núcleo. El núcleo se compacta mucho más al cambiar las histonas por protaminas, de tal forma que no puede haber ni replicación ni transcripción (Fase G0 del ciclo celular). La inactividad transcripcional del núcleo hace que el espermatozoide sea dependiente de modificaciones postranscripcionales como la fosforilación de proteínas necesarias para adaptar su función de acuerdo a las necesidades. Las fosforilaciones ocurren desde las células germinales, casi siempre en los residuos de tirosina, distinto a otras células eucarióticas en las que la mayoría de las fosforilaciones ocurren en residuos de serina y treonina.
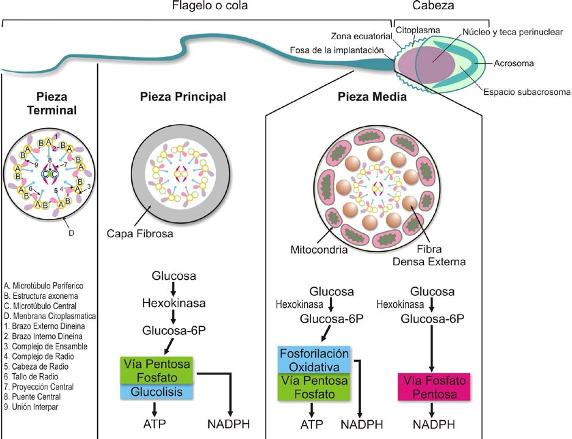
Figura 1. Morfología del espermatozoide y metabolismo energético que realiza esta célula a partir de glucosa en la pieza principal y en la pieza media.
En la fase acrosomal la espermátide gira de tal forma que el acrosoma queda en dirección de la membrana basal; se depositan gránulos en el acrosoma, el citoplasma se desplaza hacia la base de la cabeza y se localiza por debajo de la unión núcleo axonema; las mitocondrias se agrupan alrededor de este último en su parte cercana al núcleo, formando la pieza media. En esta fase el espermatozoide adquiere su morfología definitiva. Finalmente se sucede la fase de maduración, donde se observan las características finales de los espermatozoides: forma de la cabeza característica de cada especie (oval y plana), cubierta en sus dos terceras partes por el acrosoma; y la cola compuesta por las piezas media, principal y terminal (véase Figura 1); en la pieza media se encuentran las mitocondrias en forma de hélice. En esta fase se elimina gran parte del citoplasma por desplazamiento del mismo hacia la pieza terminal de la cola originando la llamada gota citoplasmática. El proceso de maduración termina con la espermiación, o liberación de los espermatozoides a la luz del túbulo seminífero. Mediante movimientos peristálticos los espermas son transportados de la rete testis a los ductos eferentes y de allí al epidídimo en cuya cola se almacenan.
Durante la eyaculación los espermatozoides junto con el plasma seminal pasan por la uretra y a través de movimientos peristálticos se liberan en el tracto genital femenino. La eyaculación es el reflejo de expulsión de los espermatozoides y el plasma seminal fuera del tracto reproductivo. El reflejo eyaculatorio es el resultado de la estimulación sensorial especialmente en el glande, lo que causa contracciones musculares coordinadas. Una vez se introduce el pene en la vagina se inicia el reflejo por impulsos que se transmiten del glande a través del nervio púdico hasta la región lumbosacra de la médula espinal. Así el semen es forzado a pasar a la uretra lo que induce la contracción de los músculos uretrales, isquicavernosos y bulboespongiosos. El eyaculado contiene, además, las secreciones de las glándulas anexas (vesículas seminales, próstata, glándulas bulbouretrales).
Movimiento del flagelo
El espermatozoide adquiere la capacidad de mover el flagelo en su tránsito por el epidídimo, pero el movimiento empieza después de la eyaculación. Este proceso es conocido como la activación del esperma. El movimiento del flagelo es característico y consiste en un bateo simétrico de la cola que hace que el espermatozoide se desplace en forma progresiva. El esperma pasa rápidamente a través del cuello y el útero de la hembra; a 10 minutos de la deposición del semen en el tracto femenino se encuentran espermatozoides en la unión útero-tubal (véase Figura 2). De aquí los espermatozoides pasan a lo que se conoce como el reservorio del oviducto (istmo) caracterizado por cilios luminales epiteliales y plegamientos de la mucosa que forman criptas (véase Figura 2). Los espermatozoides son retenidos en las criptas oviductales y allí pierden los factores decapacitantes como mucopolisacáridos y proteínas que habían aportado las glándulas anexas; éste es el comienzo del proceso conocido como capacitación (véanse Figuras 2 y 4), nombre que indica el potencial que adquiere el espermatozoide para hiperactivarse y para lograr la reacción acrosomal. Este proceso se lleva a cabo en las criptas del istmo donde se adosan los espermas, y termina con la liberación del mismo hacia el ámpula (véase Figura 2); aquí al encontrar el oocito ocurre el reconocimiento y la adherencia para que el espermatozoide empiece a atravesar la zona pelúcida.
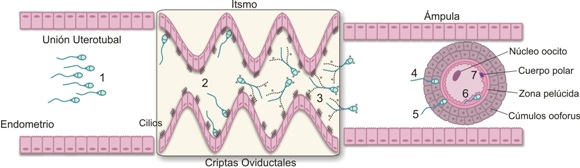
Figura 2. Secuencia de los procesos que sufre el espermatozoide en el tracto reproductivo de la hembra: 1) Activación, 2) Capacitación, 3) Hiperactivación, 4) Reconocimiento entre gametos, 5) Reacción acrosomal, 6) Adhesión y 7) Fusión.
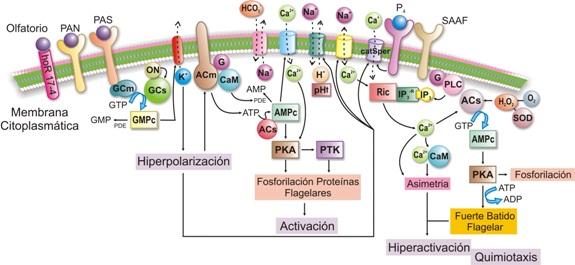
Figura 3. Modelo de los eventos moleculares que inducen la activación y la hiperactivación del flagelo.
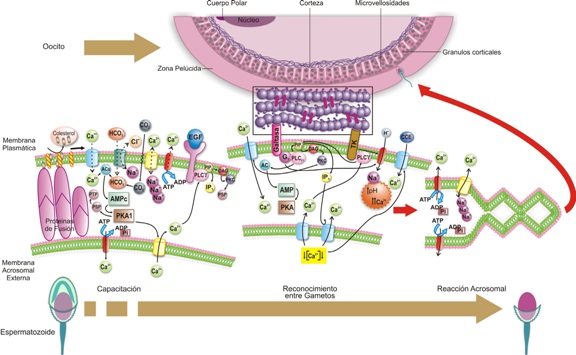
Figura 4. Modelo de eventos moleculares que se suceden durante la capacitación, el reconocimiento entre los gametos y la reacción acrosomal.
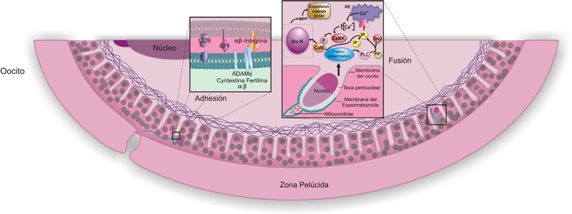
Figura 5. Fertilización: adhesión y fusión.
La reacción acrosomal es un proceso de fusión de la membrana citoplasmática externa, con la membrana acrosomal externa en la zona apical de la cabeza; de aquí se liberan enzimas que estaban almacenadas dentro de esta vesícula exocítica, esto es fundamental para el paso a través de la zona pelúcida.
Cuando el espermatozoide alcanza el istmo del oviducto inicia un movimiento asimétrico, amplio y acelerado del flagelo (característico de la hiperactivación), lo que lo lleva a moverse en círculos y lo ayuda a liberarse de las criptas oviductales para avanzar a través del lumen y alcanzar el ámpula, atravesar el cúmulus ooforus (células de la granulosa que rodean el oocito) y aponerse a la zona pelúcida donde es reconocido (reconocimiento entre gametos) (véanse Figuras 2 y 4). Una vez que el espermatozoide alcanza el espacio perivitelino, se produce la adherencia (véase Figura 5) entre la membrana plasmática de la zona ecuatorial espermática y las microvellosidades de la membrana citoplásmica del oocito; luego se fusionan las dos membranas (véase Figura 5); y de esta manera el núcleo y demás organelas de la célula espermática ingresan al ooplasma del oocito.
Los depósitos de Ca++ almacenados entre la teca perinuclear y el núcleo del espermatozoide son liberados al ooplasma, junto con la “oscilina” (factor espermático), que es una proteína espermática muy parecida a la enzima bacterial isomerasa glucosalina 6 fosfato (GNPI) o a la deaminasa (GNPDA). A este factor se le atribuye la acción de inducir las oscilaciones de calcio que conducen a la activación del oocito. Esta activación comprende: oscilaciones intracelulares de Ca++, la expulsión del segundo cuerpo polar, la formación del pronúcleo femenino, el reemplazo de las protaminas por histonas en el núcleo espermático, la formación del pronúcleo masculino y la ubicación del centríolo espermático para formar el aster masculino que es necesario para la migración de su pronúcleo. Las oscilaciones de Ca++ inducen también la exocitosis del contenido enzimático de los gránulos corticales almacenados en la periferia citosólica del oocito, modificando tanto la zona pelúcida como la membrana del oocito, para prevenir la poliespermia.
Activación e hiperactivación
La movilidad del esperma se desencadena por cambios en el medio iónico extracelular, por interacción con ligandos específicos y por glucosa, presentes en el líquido seminal y en el tracto reproductivo femenino; estos cambios inducen señales citosólicas flagelares, a través de la fosforilación de proteínas, de canales de Ca++ y de vías dependientes de nucleótidos cíclicos (GMPc y AMPc) (véase Figura 3).
Los ligandos específicos más conocidos son: la progesterona y el esteroide sulfatado SAAF (Sperm Activating and Attracting Factor) que inducen la entrada de Ca++; el péptido activador de espermatozoide (PAS) y el péptido atrial natriurético (PAN) que actúan, ya sea por medio de un receptor de membrana, o por activación directa de la guanilil ciclasa ligada a membrana (GCm).
Otros ligandos específicos son los factores de tipo olfatorio y odorante (hOR17-4) producidos por el oocito para inducir la quimiotaxis del espermatozoide.
Los cationes y los aniones también juegan un papel importante en la modulación de la movilidad espermática, el efecto mayor se atribuye a cationes tales como Ca++, Na+, K+ y H+. El tránsito de estos cationes inducido por diferencias de concentración extra e intracelular, lleva a cambios en la composición iónica intracelular y subsecuentemente a cambios en el potencial funcional y de movilidad del espermatozoide. Los cambios que produce la glucosa se describirán más adelante.
La activación se desencadena cuando las señales extracelulares de las que hablamos anteriormente activan las ciclasas, lo que produce un aumento transitorio de GMPc, de AMPc y la activación de la guanilil ciclasa transmembranal (GCm) o soluble (GCs). La GCm se activa por la unión del péptido PAS a su receptor, o por el aumento en la concentración de GMPc; a la GCs la activa el óxido nítrico (ON) producido por acción de la óxido nítrico sintasa. Consecutivamente se abren los canales de K+ dependientes de GMPc, lo que provoca la salida de K+ con la consecuente hiperpolarización de la membrana espermática dependiente de AMPc.
La hiperpolarización activa el intercambiador de voltaje dependiente Na+/H+ con la consecuente salida de H+ lo que induce la alcalinización del citosol y activa las dineínas (véase Figura 1).
El aumento de AMPc se debe a la activación de la adenilil ciclasa de dominio transmembranal (ACm), y a la activación de la fosfodiesterasa (PDE), reguladas, a su vez, por la subunidad alfa de la proteína G y por Ca++ /CaM (calmodulina) respectivamente. La forma soluble de la adenilil ciclasa (ACs) también produce AMPc, y en mayores cantidades que la ACm debido a que es independiente de proteína G. Esta ACs es dependiente de bicarbonato (HCO3-), que ingresa por mediación del cotransportador Na+/HCO3-. El Ca++ también ingresa a través del CatSper (canales catiónicos específicos del esperma). El siguiente paso en la activación de la movilidad flagelar es la fosforilación y la defosforilación de tirosinas, mediadas por la proteína quinasa dependiente de AMPc (PKA) (véase Figura 3). La subunidad catalítica de la PKA, posee una estructura única y específica en el espermatozoide y parece que se encuentra anclada a los microtúbulos. La PKA está ubicada muy cerca al brazo de dineína externo y esto podría explicar la rápida fosforilación que sufren los polipéptidos de la cadena liviana de este brazo (Tctex-2) en el momento de la activación de la movilidad. Estas PKA se fijan en dominios de algunas de las proteínas relacionadas con la activación y candidatas a ser fosforiladas; ellas son 1) las hexoquinasas, 2) las AKAP (AKAP 82, AKAP 220, AKAP 110, AKAP 3, AKAP 4, AKAP 97). Las AKAP están presentes en los radios y los pares de microtúbulos externos del axonema; la AKAP 82 se ha encontrado en abundancia en la capa fibrosa del flagelo (pieza principal) (véase Figura 1), 3) una proteína no caracterizada de 15 kDa en la base de flagelo y 4) la glicógeno sintasa-quinasa.
La defosforilación de las proteínas está a cargo de fosfatasas entre otras la tipo B2 dependiente de Ca++, que fue encontrada unida al axonema y relacionada con la regulación de la fosforilación de la dineína.
El espermatozoide, activado y capturado por las microvellosidades del istmo del oviducto (véase Figura 2), se capacita, y esto desencadena señales intracelulares que inducen la hiperactivación (véanse Figuras 2 y 3). La activación y la hiperactivación utilizan mecanismos moleculares similares para generar el movimiento del flagelo cuyo eje funcional es el axonema y cuya proteína motora principal es la dineína (véase Figura 1). El axonema, además, está compuesto por microtúbulos, moléculas chaperonas, proteínas fijadoras de calcio y proteínas quinasas/fosfatasas. Es una estructura de 9x2 pares de microtúbulos que corre a lo largo del flagelo rodeado, en la pieza media, por mitocondrias y capas de fibra densa; en la pieza principal está rodeado por una capa fibrosa, y en la final está en contacto directo con la membrana plasmática (véase Figura 1).
El movimiento del flagelo se da por la activación de los complejos de ensamble y de regulación de dineína; el ATP provee la fuerza que se requiere para el deslizamiento entre los brazos y los microtúbulos, y la hidrólisis del mismo, junto con la ruptura de la unión entre el microtúbulo B. La dineína garantiza que el movimiento continúe como reacción en cadena de los nueve pares de microtúbulos externos generando el movimiento en forma de bateo.
Simultáneamente se dan fenómenos de fosforilación y defosforilación asociados a la unión del brazo externo del microtúbulo A, al microtúbulo B del brazo adyacente. El par de microtúbulos centrales con dominio AKAP, dirigen el plano de inclinación flagelar, enviando señales a los radios que regulan el brazo interno de dineína por medio de fosforilación/defosforilación de proteínas. Los radios están compuestos por 22 polipéptidos con una proteína de 97 kDa con un dominio AKAP en su base. El par de microtúbulos centrales al dirigir el plano de inclinación, determinan la asimetría de la onda de movimiento flagelar,característica y diferencial de la hiperactivación.
La capa fibrosa de la pieza principal (véase Figura 1), está compuesta por al menos 18 polipéptidos y parece servir como andamio de algunas enzimas en el metabolismo energético, y como molécula de señal para desencadenar la movilidad espermática. Esta capa está compuesta por varias proteínas, entre ellas la hexoquinasa y las AKAP 3, AKAP 4, AKAP 80; relacionadas con la fosforilación de tirosina de otros componentes de la capa fibrosa.
El metabolismo energético a partir de glucosa (véase Figura 1) es requerido para la activación del axonema e incluye la producción y regeneración de ATP y de intermediarios como el NADPH, necesarios para las vías de señalización interna, que conducen a la fosforilación de las proteínas flagelares al movimiento. El ingreso de glucosa al citosol se da a través de transportadores como el GLUT3 en la pieza intermedia o el GLUT8 en la región acrosomal, o por reservas endógenas en forma de glicógeno. La glucosa es transformada en glucosa-6- fosfato por la enzima hexoquinasa, para generar ATP o NADPH, por una de tres vías, dependiendo del dominio del flagelo: la glicolítica, la vía pentosa fosfato y la vía mitocondrial de la fosforilación oxidativa; ésta última fuente principal de ATP (véase Figura 1).
El NADPH está involucrado en reacciones de oxido-reducción y es requerido para enzimas como la oxidasa que genera superóxido y la oxido nítrico sintasa que genera óxido nítrico.
La hiperactivación del flagelo se debe al Ca++ que se fija a sus proteínas fijadoras en el brazo externo de la dineína, lo que induce el movimiento asimétrico. Este Ca++ proviene de varias fuentes: de las reservas intracelulares de Ca++ (RIC) mediada por los receptores de inositol 1,4,5 trifosfato (IP3), del Ca++ proveniente de la mitocondria o de la teca perinuclear (véase Figura 3) o del Ca++ que ingresa gracias a sistemas antiporte Na+/H+ y Na+/Ca+++. Además del calcio, la hiperactivación requiere de AMPc, cuya producción parece estar modulada por las especies reactivas de oxígeno (ROS), entre las que se encuentran el superóxido (SO) y el óxido nítrico ON (véase Figura 3). El superóxido se origina en el transporte de electrones en la mitocondria y es trasformado a peróxido de hidrógeno por la enzima superóxido dismutasa (SOD). Este peróxido activa la adenililciclasa de membrana, que actúa corriente arriba en la ruta de PKA, para que ésta fosforile algunas proteínas flagelares. (véase Figura 3).
La capacitación
El espermatozoide, en la cola del epidídimo y en el vas deferens, sufre cambios en los dominios de los esteroles de membrana, en la cabeza y en la cola, confiriéndo una distribución heterogénea de los mismos a los largo de toda la membrana. Estos dominios llamados complejos de esterol-caveolina, sirven como andamio en la membrana para acoplar proteínas que inducirán diferentes rutas de señalización. En la cabeza hay dos subdominios de membrana plasmática, la acrosomal y la subacrosomal. La primera cubre la región del mismo nombre y se caracteriza por presentar islas de composición ordenada de colesterol y esfingolípidos anclados a caveolinas, inmersas en una membrana de composición “desordenada”. La segunda, o región subacrosomal, es rica en fosfolípidos. Las islas juegan un papel importante en la compartamentalización de las vías de señalización en regiones específicas de la célula; son diseños prefabricados durante la espermatocitogénesis ya que, como se mencionó anteriormente, el espermatozoide, una vez en el lumen testicular, no puede transcribir ni traducir. La capacitación, se caracteriza por la salida de colesterol de la membrana y el ingreso de Ca++ y HCO3- al citosol (véase Figura 4).
El fluido oviductal es rico en albúminas y HDL, capaces de retirar el colesterol de la membrana del esperma, lo que la hace más fluida al producir la ruptura de la unión de las caveolinas con las proteínas de fusión; estas últimas al quedar libres forman complejos de señalización de fusión de membranas (reacción acrosomal); el aumento de la fluidez también permite que proteínas integrales puedan interactuar con las proteínas ancladas a membrana. La pérdida de colesterol favorece además la translocación de algunas proteínas a la zona ecuatorial, donde son necesarias para que el espermatozoide posteriormente pueda adherirse al oocito.
La salida de colesterol durante la capacitación también induce la activación de los canales de Ca++ dependientes de voltaje y de los canales de HCO3-. El ingreso de éste último al citosol, activa la adenililciclasa dependiente de HCO3- (ACs) que aumenta las concentraciones de AMP cíclico, activando, a su vez, la proteína quinasa A1 (PKA1); esta última fosforila algunas proteínas en los residuos de serina desencadenando la fosforilación de las tirosinas de otras proteínas citosólicas. La PKA1 también activaría en la membrana acrosomal externa un canal de Ca++, con el consecuente aumento de las concentraciones citosólicas de este catión, las cuales gradualmente activan la fosfolipasa Cγ acoplada al receptor del factor de crecimiento epidermal.
Interacción espermatozoide zona pelúcida y reacción acrosomal
La zona pelúcida expone glicoproteínas de reconocimiento (véase Figura 4) que interactúan con la membrana del espermatozoide de varias formas: proteína –proteína, donde se reconoce el receptor de 95KDa del esperma (receptor tirosina quinasa, TK); y proteína–carbohidrato para el reconocimiento de la Galtasa como receptor acoplado a proteína G, que se une a la N acetil glucosamina de la ZP3. Una vez que acoplan estas moléculas se activa la fosfolipasa C β1 (PLCβ1) por una parte y, por la otra, posiblemente se activa la adenilil ciclasa, con el subsecuente aumento los niveles de AMP cíclico que activarían la proteína quinasa A (PKA), y permiten la apertura de un canal de Ca++, voltaje dependiente, en la membrana acrosomal externa. Este pequeño incremento de Ca++ intracitoplasmático activa la fosfolipasa Cγ que se une al receptor de tirosina quinasa para inducir una ruta de señalización vía PIP2.
Cuando se da la interacción proteína-carbohidratos, el receptor unido a proteína G, activa la fosfolipasa Cβ1, lo que induce la hidrólisis del fosfatidil inositol difosfato (PIP2) con generación de diacil glicerol (DAG) e inositol trifosfato (IP3). El DAG activa la proteina quinasa C, lo que lleva a la apertura de un canal dependiente voltaje en la membrana plasmática y permite el ingreso de altas cantidades de Ca++. El IP3 y la PKA abren un canal en la membrana acrosomal externa, lo que lleva a la deplesión del Ca++ en el espacio intraacrosomal, y esto activa el canal de Ca++ “capacitante” en la membrana citoplasmática (véase Figura 4).
El receptor de TK y la proteína G, pueden activar un intercambiador de Na+-H+ en la membrana plasmática, lo que alcaliniza el pH citosólico. El incremento de Ca++ regula su propia salida tanto en la membrana plasmática como en la acrosomal, mediante la activación de canales de Ca++, dependiente de ATP, y con la activación de los intercambiadores Na+- Ca++. La PKA fosforila los residuos de serina de las proteínas (PSP); éstas, a su vez, fosforilan los residuos de tirosina de las proteínas citosólicas que junto con el aumento del pH y de Ca++ citosólico, permiten la fusión de membranas (citoplasmática y acrosomal externa) (véase Figura 4), y de esta manera se produce la exocitosis del contenido enzimático del acrosoma. El esperma atraviesa la zona pelúcida (ZP) y alcanza el espacio perivitelino, los mecanismos involucrados en este paso aún se desconocen.
Para evitar la poliespermia se suceden dos mecanismos, uno inmediato que es el cambio de potencial de membrana del oocito, que a su vez cambia la polaridad interna de la ZP; y otro subsiguiente que es la remodelación de la ZP, que ocurre por exocitosis del contenido enzimático de los gránulos corticales al espacio perivitelino; esta exocitosis está regulada por PKC dependiente de factores lipídicos e independiente de los iones de calcio. Se han identificado dos poblaciones de gránulos corticales que son liberados a tiempos y en lugares diferentes: el primer grupo, se libera durante la expulsión del primer cuerpo polar y el segundo muy cerca del sitio en donde comienza la citoquinesis de la primera división celular.
El mecanismo propuesto para la remodelación de la zona pelúcida, es a partir de la alteración de los residuos de galactosa alfa y beta de la misma, particularmente las enzimas hidrolíticas y las glicoproteínas liberadas por los gránulos corticales. Los residuos de GlcNAc de la ZP3 que contiene la zona pelúcida (receptores de la Galtasa espermática) serían removidos por la N-acetilglucaminidasa, una de las enzimas que contienen los gránulos corticales, lo que no permitiría que más espermatozoides se adhirieran a la ZP3.
La adhesión del espermatozoide y la fusión de las membranas oocito-esperma
Las vellosidades del oocito tienen integrinas α6 y β1 (véase Figura 5), a las que se adhiere el espermatozoide mediante la exposición de proteínas como la fertilina α y β y ciritestina, (proteínas de la familia ADAMS con dominios disintegrina y metaloproteasa).
Recientemente se han candidatizado las proteínas de anclaje del oocito CD9 y glicosilfosfatidil inositol (GPI) y la proteína epididimal DE, en el espermatozoide, como las moléculas candidatas a ser las responsables de la fusión membranal entre el esperma y el oocito. Sin embargo aún no se han podido analizar claramente las interacciones proteína-proteína en esta fusión, por lo que el mecanismo molecular permanece sin dilucidar.
La fusión de membranas continúa a través (véase Figura 5) de una proteína presente en el oocito, de la familia de las tetraspaninas conocida como CD9. Esta proteína se caracteriza por tener cuatro dominios transmembranales y dos bucles extracelulares; en el mayor de estos bucles se fijan los complejos de fusión del esperma. Las microvellosidades se encuentran por toda la superficie del oocito, exceptuando la zona por donde se expulsará el segundo cuerpo polar. Al fusionarse las membranas del oocito y del esperma se libera, del espermatozoide, una molécula de superficie que parece ser una proteína de 30-100 kDA llamada “oscilina” o factor espermático, que activa el oocito produciendo la liberación de Ca++ en forma de ondas (véase Figura 5). Cada vez que se sucede una onda de Ca++, cambia su concentración intracitoplasmática de 50 a 600-1000 nmol/litro.
Estas ondas se dan periódicamente cada 2 - 30 minutos dependiendo de la especie y pueden durar varias horas. El inicio de las ondas de Ca++ induce la activación del oocito que termina con la formación de los pronúcleos. En los cambios que ocurren en el oocito para que se libere el Ca++ intracelular y se produzcan los eventos corriente abajo, podrían estar involucradas las quinasas de la familia de las Src (Src-K); entre éstas la c-Fyn, y la c-Yes que están distribuidas en la corteza del oocito (véase Figura 5).
La c-Fyn también se encuentra alrededor del huso mitótico; la activación de esta quinasa resulta en la activación de la fosfolipasa C (PLC) que cataliza la hidrólisis del PIP2 a DAG e IP3. IP3 actúa en el receptor de IP3 (IP3R) que se encuentra en el retículo endoplásmico y hace liberar el calcio. El mismo calcio participa en su autorregulación y junto con otros moduladores del receptor de IP3 induce las oscilaciones intracelulares descritas, y que tienen como objetivo la activación del oocito.
Sin embargo, cómo se activan las Src-K es una pregunta que se permanece sin respuesta.
Glosario: abreviaturas usadas en las figuras
MP: membrana plasmática
MAE: membrana acrosomal externa
Ca++: ión calcio
PTP: Proteína tirosina fosfatada
PSP: proteína serina fosfatada
PKA1:Proteína quinasa A1
AMPc: adenosin monofosfato cíclico
ATP: adenosin trifosfato
ADP: adenosin difosfato
Pi: fósforo inorgánico
HCO3: ión bicarbonato
sAC: adenilato ciclasa activada por bicarbonato
Na+: ión Sodio
CO2-: ión dioxido de carbono
Cl-: ión cloro
EGF: factor de crecimiento epidermal
PLC γ: Fosfolipasa C gamma
PIP2: Fosfatidil inositol difosfato
DAG: Diacil glicerol
IP3: Inositol trifosfato
GALtasa:Galactosil transferasa
TK: tirosina quinasa
G1: proteína G
CCE: Calcio capacitante
CaM: calmodulina
CaMK: calmodulina quinasa
REnd: retículo endoplásmico
Ubiq: ubiquitina
MPF: factor promotor de la maduración
CD9: receptor familia citoquinas
DE: proteína epididimal
GPI: glicosil fosfatidil inositol, proteína de anclaje
AKAP: Proteínas de dominios de fijación de PKA
PAN: Péptido Atrial Natriurético.
PAS: Peptido Activador de Esperma.
ON: Óxido Nítrico.
ACm: Adenil Ciclasa de Membrana.
GCm: Guanil Ciclasa de membrana.
CaM: Calmodulina.
ACs: Adenil Ciclasa Soluble.
PKA: Proteína quinasa dependiente de AMPc
PTK: Proteina Tirosina Quinasa.
CCp: Canal de Calcio Putativo.
PLC Fosfolipasa C
IP3: Inositol Trifosfato.
RCaIC: Reserva de Calcio Intracitoplasmático
P4: progesterona
SAAF: Factor espermático de activación y atracción (sperm activating and attracting factor)
PASA: péptido activador de espermas
GCm: Guanil Ciclasa ligada a membrana
hOR17-4: factores odorantes
GCs: guanil ciclasa soluble
ON: óxido nítrico
ACm: adenilciclasa de dominio transmembranal
ACs: adenil ciclasa soluble
CatSperm: canales catiónicos del esperma
Summary
The sperm cell: from eyaculation to fertilization During the last recent years there has been a great increase of information regarding the molecular mechanisms involved in the maturation of the male gamete as well as its progression from the gonad up to fertilizing the oocyte. Along this way the sperm completes its maturation adding some molecular components; additionally along this trail take place all the processes leading to activation of the sperm for the entrance into the oocyte to initiates molecular cascades for the formation of a zygote. This article, based on updated literature proposes a model that integrates known cellular and molecular interactions and pinpoints some steps still requiring further research.
Key words: acrosome reaction, activation, adhesion, capacitation, fusion.
Referencias
1. Inaba K. Molecular architecture of the sperm flagella: molecules for motility and signaling. Zoolog Sci 2003; 20: 1043-1056. [ Links ]
2. Urner F, Sakkas D. Protein phosphorylation in mammalian spermatozoa. Reproduction 2003; 125: 17-26. [ Links ]
3. Sakkas D, Leppens-Luisier G, Lucas H, Chardonnens D, Campana A, Franken DR,. et.al. Localization of tyrosine phosphorylated proteins in human sperm and relation to capacitation and zona pellucida binding. Biol Reprod 2003; 68: 1463-1469. [ Links ]
4. Vijayaraghavan S, Mohan J, Gray H, Khatra B, Carr DW. A role for phosphorylation of glycogen synthase kinase-3alpha in bovine sperm motility regulation. Biol Reprod 2000; 62: 1647-1654. [ Links ]
5. Galindo BE, Nishigaki T, Rodriguez E, Sanchez D, Beltran C, Darszon A. Speract-receptor interaction and the modulation of ion transport in Strongylocentrotus purpuratus sea urchin sperm. Zygote 2000;8 Suppl 1:S20-1. [ Links ]
6. Galindo BE, Beltran C, Cragoe EJ, Jr., Darszon A.Participation of a K(+) channel modulated directly by cGMP in the speract-induced signaling cascade of strongylocentrotus purpuratus sea urchin sperm. Dev Biol 2000;221:285-294. [ Links ]
7. Tardif S, Lefievre L, Gagnon C, Bailey JL. Implication of cAMP during porcine sperm capacitation andprotein tyrosine phosphorylation. Mol Reprod Dev 2004; 69:428-435. [ Links ]
8. Ho HC, Suarez SS. Hyperactivation of mammalian spermatozoa: function and regulation. Reproduction 2001;122:519-526. [ Links ]
9. Baker MA, Lewis B, Hetherington L, Aitken RJ. Development of the signalling pathways associated with sperm capacitation during epididymal maturation. Mol Reprod Dev 2003; 64 :446-457. [ Links ]
10. Baker MA, Krutskikh A, Aitken RJ. Biochemical entities involved in reactive oxygen species generation by human spermatozoa. Protoplasma 2003; 221:145-151. [ Links ]
11. Travis AJ, Kopf GS. The role of cholesterol efflux in regulating the fertilization potential of mammalian spermatozoa. J Clin Invest 2002;110:731-736. [ Links ]
12. Williams CJ. Signalling mechanisms of mammalian oocyte activation. Hum Reprod Update 2002;8:313-321. [ Links ]
13. Brewis IA, Wong CH. Gamete recognition: sperm proteins that interact with the egg zona pellucida. Rev Reprod 1999;4:135-142. [ Links ]
14. Flesch FM, Gadella BM. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim Biophys Acta 2000;1469:197-235. [ Links ]
15. White D, Weerachatyanukul W, Gadella B, Kamolvarin N, Attar M, Tanphaichitr N. Role of sperm sulfogalactosylglycerolipid in mouse sperm-zona pellucida binding. Biol Reprod 2000;63:147-155. [ Links ]
16. Gadella BM, Harrison RA. The capacitating agent bicarbonate induces protein kinase A-dependent changes in phospholipid transbilayer behavior in the sperm plasma membrane. Development 2000;127: 2407-2420. [ Links ]
17. Sun QY, Breitbart H, Schatten H. Role of the MAPK cascade in mammalian germ cells. Reprod Fertil Dev 1999;11:443-450. [ Links ]
18. Breitbart H, Naor Z. Protein kinases in mammalian sperm capacitation and the acrosome reaction. RevReprod 1999; 4:151-159. [ Links ]
Martha Olivera1,2, MV, Dr Sci Agr; Tatiana Ruiz1,2, MV, Ms, PhD; Ariel Tarazona1,2,Z,Ms; Carlos Giraldo1,2,MV, Ms.
1Fisiología y Biotecnología de la Reproducción, 2Facultad de Ciencias Agrarias, Universidad de Antioquia, A.A 1226, Medellín, Colombia.
molivera@catios.udea.edu.co
(Recibido: 23 de junio, 2006; aceptado: 14 de septiembre, 2006)
Resumen
En los últimos años se ha hecho un inmenso progreso en el entendimiento de los mecanismos moleculares involucrados en la maduración del gameto masculino y su tránsito desde la gónada hasta la fertilización del oocito. A lo largo de este trayecto el espermatozoide modifica su morfología y sus componentes moleculares especialmente, y además ocurren procesos que conducen a la activación para la entrada al oocito, para activar a su vez los mecanismos que conducen a la formación del zigoto. Este artículo presenta, a partir de la interpretación de la literatura actual un modelo de los eventos que se suceden a partir de la eyaculación hasta la fertilización, con énfasis en los mecanismos celulares y moleculares conocidos, y señala algunos vacíos de información aún existentes.
Palabras clave: activación, adhesión, capacitación, fusión, reacción acrosomal.
Generalidades sobre la producción y anatomía espermática en mamíferos
La célula germinal masculina se produce en la gónada (testículo) mediante un proceso permanente de división de las células germinales o espermatogonias. El proceso de división meiótica denominado espermatogénesis está controlado hormonalmente por el eje hipófisis-hipotálamo-gónada. A partir de cada espermatogonia se producen cuatro espermatocitos haploides que permanecen unidos entre sí por puentes citoplasmáticos y a la vez están en comunicación con la célula nutricia o de Sertolli; estas últimas, a partir de moléculas señalizadoras, inducen el proceso denominado espermiogénesis o metamorfosis que convierte las espermátides en espermatozoides. Se reconocen cuatro fases características en esta transformación: la fase de Golgi, la de capuchón, la acrosomal y la de maduración.
En la fase de Golgi, la organela del mismo nombre se acerca al núcleo, desprende vesículas que se le sobreponen y poco a poco se unen para convertirse en la vesícula acrosomal que se localiza en la parte apical del núcleo (véase Figura 1). Los centríolos situados en forma de T, muy cercanos al aparato de Golgi, van migrando hacia lo que será la base del núcleo. El centríolo proximal se sitúa en la parte basal del núcleo y, a partir del centríolo distal crece el axonema (véase Figura 1) conformado por dos microtúbulos centrales y 9 pares de microtúbulos periféricos. En la fase de capuchón, la vesícula acrosomal se aplana formando una verdadera capucha sobre el núcleo. El núcleo se compacta mucho más al cambiar las histonas por protaminas, de tal forma que no puede haber ni replicación ni transcripción (Fase G0 del ciclo celular). La inactividad transcripcional del núcleo hace que el espermatozoide sea dependiente de modificaciones postranscripcionales como la fosforilación de proteínas necesarias para adaptar su función de acuerdo a las necesidades. Las fosforilaciones ocurren desde las células germinales, casi siempre en los residuos de tirosina, distinto a otras células eucarióticas en las que la mayoría de las fosforilaciones ocurren en residuos de serina y treonina.
Figura 1. Morfología del espermatozoide y metabolismo energético que realiza esta célula a partir de glucosa en la pieza principal y en la pieza media.
En la fase acrosomal la espermátide gira de tal forma que el acrosoma queda en dirección de la membrana basal; se depositan gránulos en el acrosoma, el citoplasma se desplaza hacia la base de la cabeza y se localiza por debajo de la unión núcleo axonema; las mitocondrias se agrupan alrededor de este último en su parte cercana al núcleo, formando la pieza media. En esta fase el espermatozoide adquiere su morfología definitiva. Finalmente se sucede la fase de maduración, donde se observan las características finales de los espermatozoides: forma de la cabeza característica de cada especie (oval y plana), cubierta en sus dos terceras partes por el acrosoma; y la cola compuesta por las piezas media, principal y terminal (véase Figura 1); en la pieza media se encuentran las mitocondrias en forma de hélice. En esta fase se elimina gran parte del citoplasma por desplazamiento del mismo hacia la pieza terminal de la cola originando la llamada gota citoplasmática. El proceso de maduración termina con la espermiación, o liberación de los espermatozoides a la luz del túbulo seminífero. Mediante movimientos peristálticos los espermas son transportados de la rete testis a los ductos eferentes y de allí al epidídimo en cuya cola se almacenan.
Durante la eyaculación los espermatozoides junto con el plasma seminal pasan por la uretra y a través de movimientos peristálticos se liberan en el tracto genital femenino. La eyaculación es el reflejo de expulsión de los espermatozoides y el plasma seminal fuera del tracto reproductivo. El reflejo eyaculatorio es el resultado de la estimulación sensorial especialmente en el glande, lo que causa contracciones musculares coordinadas. Una vez se introduce el pene en la vagina se inicia el reflejo por impulsos que se transmiten del glande a través del nervio púdico hasta la región lumbosacra de la médula espinal. Así el semen es forzado a pasar a la uretra lo que induce la contracción de los músculos uretrales, isquicavernosos y bulboespongiosos. El eyaculado contiene, además, las secreciones de las glándulas anexas (vesículas seminales, próstata, glándulas bulbouretrales).
Movimiento del flagelo
El espermatozoide adquiere la capacidad de mover el flagelo en su tránsito por el epidídimo, pero el movimiento empieza después de la eyaculación. Este proceso es conocido como la activación del esperma. El movimiento del flagelo es característico y consiste en un bateo simétrico de la cola que hace que el espermatozoide se desplace en forma progresiva. El esperma pasa rápidamente a través del cuello y el útero de la hembra; a 10 minutos de la deposición del semen en el tracto femenino se encuentran espermatozoides en la unión útero-tubal (véase Figura 2). De aquí los espermatozoides pasan a lo que se conoce como el reservorio del oviducto (istmo) caracterizado por cilios luminales epiteliales y plegamientos de la mucosa que forman criptas (véase Figura 2). Los espermatozoides son retenidos en las criptas oviductales y allí pierden los factores decapacitantes como mucopolisacáridos y proteínas que habían aportado las glándulas anexas; éste es el comienzo del proceso conocido como capacitación (véanse Figuras 2 y 4), nombre que indica el potencial que adquiere el espermatozoide para hiperactivarse y para lograr la reacción acrosomal. Este proceso se lleva a cabo en las criptas del istmo donde se adosan los espermas, y termina con la liberación del mismo hacia el ámpula (véase Figura 2); aquí al encontrar el oocito ocurre el reconocimiento y la adherencia para que el espermatozoide empiece a atravesar la zona pelúcida.
Figura 2. Secuencia de los procesos que sufre el espermatozoide en el tracto reproductivo de la hembra: 1) Activación, 2) Capacitación, 3) Hiperactivación, 4) Reconocimiento entre gametos, 5) Reacción acrosomal, 6) Adhesión y 7) Fusión.
Figura 3. Modelo de los eventos moleculares que inducen la activación y la hiperactivación del flagelo.
Figura 4. Modelo de eventos moleculares que se suceden durante la capacitación, el reconocimiento entre los gametos y la reacción acrosomal.
Figura 5. Fertilización: adhesión y fusión.
La reacción acrosomal es un proceso de fusión de la membrana citoplasmática externa, con la membrana acrosomal externa en la zona apical de la cabeza; de aquí se liberan enzimas que estaban almacenadas dentro de esta vesícula exocítica, esto es fundamental para el paso a través de la zona pelúcida.
Cuando el espermatozoide alcanza el istmo del oviducto inicia un movimiento asimétrico, amplio y acelerado del flagelo (característico de la hiperactivación), lo que lo lleva a moverse en círculos y lo ayuda a liberarse de las criptas oviductales para avanzar a través del lumen y alcanzar el ámpula, atravesar el cúmulus ooforus (células de la granulosa que rodean el oocito) y aponerse a la zona pelúcida donde es reconocido (reconocimiento entre gametos) (véanse Figuras 2 y 4). Una vez que el espermatozoide alcanza el espacio perivitelino, se produce la adherencia (véase Figura 5) entre la membrana plasmática de la zona ecuatorial espermática y las microvellosidades de la membrana citoplásmica del oocito; luego se fusionan las dos membranas (véase Figura 5); y de esta manera el núcleo y demás organelas de la célula espermática ingresan al ooplasma del oocito.
Los depósitos de Ca++ almacenados entre la teca perinuclear y el núcleo del espermatozoide son liberados al ooplasma, junto con la “oscilina” (factor espermático), que es una proteína espermática muy parecida a la enzima bacterial isomerasa glucosalina 6 fosfato (GNPI) o a la deaminasa (GNPDA). A este factor se le atribuye la acción de inducir las oscilaciones de calcio que conducen a la activación del oocito. Esta activación comprende: oscilaciones intracelulares de Ca++, la expulsión del segundo cuerpo polar, la formación del pronúcleo femenino, el reemplazo de las protaminas por histonas en el núcleo espermático, la formación del pronúcleo masculino y la ubicación del centríolo espermático para formar el aster masculino que es necesario para la migración de su pronúcleo. Las oscilaciones de Ca++ inducen también la exocitosis del contenido enzimático de los gránulos corticales almacenados en la periferia citosólica del oocito, modificando tanto la zona pelúcida como la membrana del oocito, para prevenir la poliespermia.
Activación e hiperactivación
La movilidad del esperma se desencadena por cambios en el medio iónico extracelular, por interacción con ligandos específicos y por glucosa, presentes en el líquido seminal y en el tracto reproductivo femenino; estos cambios inducen señales citosólicas flagelares, a través de la fosforilación de proteínas, de canales de Ca++ y de vías dependientes de nucleótidos cíclicos (GMPc y AMPc) (véase Figura 3).
Los ligandos específicos más conocidos son: la progesterona y el esteroide sulfatado SAAF (Sperm Activating and Attracting Factor) que inducen la entrada de Ca++; el péptido activador de espermatozoide (PAS) y el péptido atrial natriurético (PAN) que actúan, ya sea por medio de un receptor de membrana, o por activación directa de la guanilil ciclasa ligada a membrana (GCm).
Otros ligandos específicos son los factores de tipo olfatorio y odorante (hOR17-4) producidos por el oocito para inducir la quimiotaxis del espermatozoide.
Los cationes y los aniones también juegan un papel importante en la modulación de la movilidad espermática, el efecto mayor se atribuye a cationes tales como Ca++, Na+, K+ y H+. El tránsito de estos cationes inducido por diferencias de concentración extra e intracelular, lleva a cambios en la composición iónica intracelular y subsecuentemente a cambios en el potencial funcional y de movilidad del espermatozoide. Los cambios que produce la glucosa se describirán más adelante.
La activación se desencadena cuando las señales extracelulares de las que hablamos anteriormente activan las ciclasas, lo que produce un aumento transitorio de GMPc, de AMPc y la activación de la guanilil ciclasa transmembranal (GCm) o soluble (GCs). La GCm se activa por la unión del péptido PAS a su receptor, o por el aumento en la concentración de GMPc; a la GCs la activa el óxido nítrico (ON) producido por acción de la óxido nítrico sintasa. Consecutivamente se abren los canales de K+ dependientes de GMPc, lo que provoca la salida de K+ con la consecuente hiperpolarización de la membrana espermática dependiente de AMPc.
La hiperpolarización activa el intercambiador de voltaje dependiente Na+/H+ con la consecuente salida de H+ lo que induce la alcalinización del citosol y activa las dineínas (véase Figura 1).
El aumento de AMPc se debe a la activación de la adenilil ciclasa de dominio transmembranal (ACm), y a la activación de la fosfodiesterasa (PDE), reguladas, a su vez, por la subunidad alfa de la proteína G y por Ca++ /CaM (calmodulina) respectivamente. La forma soluble de la adenilil ciclasa (ACs) también produce AMPc, y en mayores cantidades que la ACm debido a que es independiente de proteína G. Esta ACs es dependiente de bicarbonato (HCO3-), que ingresa por mediación del cotransportador Na+/HCO3-. El Ca++ también ingresa a través del CatSper (canales catiónicos específicos del esperma). El siguiente paso en la activación de la movilidad flagelar es la fosforilación y la defosforilación de tirosinas, mediadas por la proteína quinasa dependiente de AMPc (PKA) (véase Figura 3). La subunidad catalítica de la PKA, posee una estructura única y específica en el espermatozoide y parece que se encuentra anclada a los microtúbulos. La PKA está ubicada muy cerca al brazo de dineína externo y esto podría explicar la rápida fosforilación que sufren los polipéptidos de la cadena liviana de este brazo (Tctex-2) en el momento de la activación de la movilidad. Estas PKA se fijan en dominios de algunas de las proteínas relacionadas con la activación y candidatas a ser fosforiladas; ellas son 1) las hexoquinasas, 2) las AKAP (AKAP 82, AKAP 220, AKAP 110, AKAP 3, AKAP 4, AKAP 97). Las AKAP están presentes en los radios y los pares de microtúbulos externos del axonema; la AKAP 82 se ha encontrado en abundancia en la capa fibrosa del flagelo (pieza principal) (véase Figura 1), 3) una proteína no caracterizada de 15 kDa en la base de flagelo y 4) la glicógeno sintasa-quinasa.
La defosforilación de las proteínas está a cargo de fosfatasas entre otras la tipo B2 dependiente de Ca++, que fue encontrada unida al axonema y relacionada con la regulación de la fosforilación de la dineína.
El espermatozoide, activado y capturado por las microvellosidades del istmo del oviducto (véase Figura 2), se capacita, y esto desencadena señales intracelulares que inducen la hiperactivación (véanse Figuras 2 y 3). La activación y la hiperactivación utilizan mecanismos moleculares similares para generar el movimiento del flagelo cuyo eje funcional es el axonema y cuya proteína motora principal es la dineína (véase Figura 1). El axonema, además, está compuesto por microtúbulos, moléculas chaperonas, proteínas fijadoras de calcio y proteínas quinasas/fosfatasas. Es una estructura de 9x2 pares de microtúbulos que corre a lo largo del flagelo rodeado, en la pieza media, por mitocondrias y capas de fibra densa; en la pieza principal está rodeado por una capa fibrosa, y en la final está en contacto directo con la membrana plasmática (véase Figura 1).
El movimiento del flagelo se da por la activación de los complejos de ensamble y de regulación de dineína; el ATP provee la fuerza que se requiere para el deslizamiento entre los brazos y los microtúbulos, y la hidrólisis del mismo, junto con la ruptura de la unión entre el microtúbulo B. La dineína garantiza que el movimiento continúe como reacción en cadena de los nueve pares de microtúbulos externos generando el movimiento en forma de bateo.
Simultáneamente se dan fenómenos de fosforilación y defosforilación asociados a la unión del brazo externo del microtúbulo A, al microtúbulo B del brazo adyacente. El par de microtúbulos centrales con dominio AKAP, dirigen el plano de inclinación flagelar, enviando señales a los radios que regulan el brazo interno de dineína por medio de fosforilación/defosforilación de proteínas. Los radios están compuestos por 22 polipéptidos con una proteína de 97 kDa con un dominio AKAP en su base. El par de microtúbulos centrales al dirigir el plano de inclinación, determinan la asimetría de la onda de movimiento flagelar,característica y diferencial de la hiperactivación.
La capa fibrosa de la pieza principal (véase Figura 1), está compuesta por al menos 18 polipéptidos y parece servir como andamio de algunas enzimas en el metabolismo energético, y como molécula de señal para desencadenar la movilidad espermática. Esta capa está compuesta por varias proteínas, entre ellas la hexoquinasa y las AKAP 3, AKAP 4, AKAP 80; relacionadas con la fosforilación de tirosina de otros componentes de la capa fibrosa.
El metabolismo energético a partir de glucosa (véase Figura 1) es requerido para la activación del axonema e incluye la producción y regeneración de ATP y de intermediarios como el NADPH, necesarios para las vías de señalización interna, que conducen a la fosforilación de las proteínas flagelares al movimiento. El ingreso de glucosa al citosol se da a través de transportadores como el GLUT3 en la pieza intermedia o el GLUT8 en la región acrosomal, o por reservas endógenas en forma de glicógeno. La glucosa es transformada en glucosa-6- fosfato por la enzima hexoquinasa, para generar ATP o NADPH, por una de tres vías, dependiendo del dominio del flagelo: la glicolítica, la vía pentosa fosfato y la vía mitocondrial de la fosforilación oxidativa; ésta última fuente principal de ATP (véase Figura 1).
El NADPH está involucrado en reacciones de oxido-reducción y es requerido para enzimas como la oxidasa que genera superóxido y la oxido nítrico sintasa que genera óxido nítrico.
La hiperactivación del flagelo se debe al Ca++ que se fija a sus proteínas fijadoras en el brazo externo de la dineína, lo que induce el movimiento asimétrico. Este Ca++ proviene de varias fuentes: de las reservas intracelulares de Ca++ (RIC) mediada por los receptores de inositol 1,4,5 trifosfato (IP3), del Ca++ proveniente de la mitocondria o de la teca perinuclear (véase Figura 3) o del Ca++ que ingresa gracias a sistemas antiporte Na+/H+ y Na+/Ca+++. Además del calcio, la hiperactivación requiere de AMPc, cuya producción parece estar modulada por las especies reactivas de oxígeno (ROS), entre las que se encuentran el superóxido (SO) y el óxido nítrico ON (véase Figura 3). El superóxido se origina en el transporte de electrones en la mitocondria y es trasformado a peróxido de hidrógeno por la enzima superóxido dismutasa (SOD). Este peróxido activa la adenililciclasa de membrana, que actúa corriente arriba en la ruta de PKA, para que ésta fosforile algunas proteínas flagelares. (véase Figura 3).
La capacitación
El espermatozoide, en la cola del epidídimo y en el vas deferens, sufre cambios en los dominios de los esteroles de membrana, en la cabeza y en la cola, confiriéndo una distribución heterogénea de los mismos a los largo de toda la membrana. Estos dominios llamados complejos de esterol-caveolina, sirven como andamio en la membrana para acoplar proteínas que inducirán diferentes rutas de señalización. En la cabeza hay dos subdominios de membrana plasmática, la acrosomal y la subacrosomal. La primera cubre la región del mismo nombre y se caracteriza por presentar islas de composición ordenada de colesterol y esfingolípidos anclados a caveolinas, inmersas en una membrana de composición “desordenada”. La segunda, o región subacrosomal, es rica en fosfolípidos. Las islas juegan un papel importante en la compartamentalización de las vías de señalización en regiones específicas de la célula; son diseños prefabricados durante la espermatocitogénesis ya que, como se mencionó anteriormente, el espermatozoide, una vez en el lumen testicular, no puede transcribir ni traducir. La capacitación, se caracteriza por la salida de colesterol de la membrana y el ingreso de Ca++ y HCO3- al citosol (véase Figura 4).
El fluido oviductal es rico en albúminas y HDL, capaces de retirar el colesterol de la membrana del esperma, lo que la hace más fluida al producir la ruptura de la unión de las caveolinas con las proteínas de fusión; estas últimas al quedar libres forman complejos de señalización de fusión de membranas (reacción acrosomal); el aumento de la fluidez también permite que proteínas integrales puedan interactuar con las proteínas ancladas a membrana. La pérdida de colesterol favorece además la translocación de algunas proteínas a la zona ecuatorial, donde son necesarias para que el espermatozoide posteriormente pueda adherirse al oocito.
La salida de colesterol durante la capacitación también induce la activación de los canales de Ca++ dependientes de voltaje y de los canales de HCO3-. El ingreso de éste último al citosol, activa la adenililciclasa dependiente de HCO3- (ACs) que aumenta las concentraciones de AMP cíclico, activando, a su vez, la proteína quinasa A1 (PKA1); esta última fosforila algunas proteínas en los residuos de serina desencadenando la fosforilación de las tirosinas de otras proteínas citosólicas. La PKA1 también activaría en la membrana acrosomal externa un canal de Ca++, con el consecuente aumento de las concentraciones citosólicas de este catión, las cuales gradualmente activan la fosfolipasa Cγ acoplada al receptor del factor de crecimiento epidermal.
Interacción espermatozoide zona pelúcida y reacción acrosomal
La zona pelúcida expone glicoproteínas de reconocimiento (véase Figura 4) que interactúan con la membrana del espermatozoide de varias formas: proteína –proteína, donde se reconoce el receptor de 95KDa del esperma (receptor tirosina quinasa, TK); y proteína–carbohidrato para el reconocimiento de la Galtasa como receptor acoplado a proteína G, que se une a la N acetil glucosamina de la ZP3. Una vez que acoplan estas moléculas se activa la fosfolipasa C β1 (PLCβ1) por una parte y, por la otra, posiblemente se activa la adenilil ciclasa, con el subsecuente aumento los niveles de AMP cíclico que activarían la proteína quinasa A (PKA), y permiten la apertura de un canal de Ca++, voltaje dependiente, en la membrana acrosomal externa. Este pequeño incremento de Ca++ intracitoplasmático activa la fosfolipasa Cγ que se une al receptor de tirosina quinasa para inducir una ruta de señalización vía PIP2.
Cuando se da la interacción proteína-carbohidratos, el receptor unido a proteína G, activa la fosfolipasa Cβ1, lo que induce la hidrólisis del fosfatidil inositol difosfato (PIP2) con generación de diacil glicerol (DAG) e inositol trifosfato (IP3). El DAG activa la proteina quinasa C, lo que lleva a la apertura de un canal dependiente voltaje en la membrana plasmática y permite el ingreso de altas cantidades de Ca++. El IP3 y la PKA abren un canal en la membrana acrosomal externa, lo que lleva a la deplesión del Ca++ en el espacio intraacrosomal, y esto activa el canal de Ca++ “capacitante” en la membrana citoplasmática (véase Figura 4).
El receptor de TK y la proteína G, pueden activar un intercambiador de Na+-H+ en la membrana plasmática, lo que alcaliniza el pH citosólico. El incremento de Ca++ regula su propia salida tanto en la membrana plasmática como en la acrosomal, mediante la activación de canales de Ca++, dependiente de ATP, y con la activación de los intercambiadores Na+- Ca++. La PKA fosforila los residuos de serina de las proteínas (PSP); éstas, a su vez, fosforilan los residuos de tirosina de las proteínas citosólicas que junto con el aumento del pH y de Ca++ citosólico, permiten la fusión de membranas (citoplasmática y acrosomal externa) (véase Figura 4), y de esta manera se produce la exocitosis del contenido enzimático del acrosoma. El esperma atraviesa la zona pelúcida (ZP) y alcanza el espacio perivitelino, los mecanismos involucrados en este paso aún se desconocen.
Para evitar la poliespermia se suceden dos mecanismos, uno inmediato que es el cambio de potencial de membrana del oocito, que a su vez cambia la polaridad interna de la ZP; y otro subsiguiente que es la remodelación de la ZP, que ocurre por exocitosis del contenido enzimático de los gránulos corticales al espacio perivitelino; esta exocitosis está regulada por PKC dependiente de factores lipídicos e independiente de los iones de calcio. Se han identificado dos poblaciones de gránulos corticales que son liberados a tiempos y en lugares diferentes: el primer grupo, se libera durante la expulsión del primer cuerpo polar y el segundo muy cerca del sitio en donde comienza la citoquinesis de la primera división celular.
El mecanismo propuesto para la remodelación de la zona pelúcida, es a partir de la alteración de los residuos de galactosa alfa y beta de la misma, particularmente las enzimas hidrolíticas y las glicoproteínas liberadas por los gránulos corticales. Los residuos de GlcNAc de la ZP3 que contiene la zona pelúcida (receptores de la Galtasa espermática) serían removidos por la N-acetilglucaminidasa, una de las enzimas que contienen los gránulos corticales, lo que no permitiría que más espermatozoides se adhirieran a la ZP3.
La adhesión del espermatozoide y la fusión de las membranas oocito-esperma
Las vellosidades del oocito tienen integrinas α6 y β1 (véase Figura 5), a las que se adhiere el espermatozoide mediante la exposición de proteínas como la fertilina α y β y ciritestina, (proteínas de la familia ADAMS con dominios disintegrina y metaloproteasa).
Recientemente se han candidatizado las proteínas de anclaje del oocito CD9 y glicosilfosfatidil inositol (GPI) y la proteína epididimal DE, en el espermatozoide, como las moléculas candidatas a ser las responsables de la fusión membranal entre el esperma y el oocito. Sin embargo aún no se han podido analizar claramente las interacciones proteína-proteína en esta fusión, por lo que el mecanismo molecular permanece sin dilucidar.
La fusión de membranas continúa a través (véase Figura 5) de una proteína presente en el oocito, de la familia de las tetraspaninas conocida como CD9. Esta proteína se caracteriza por tener cuatro dominios transmembranales y dos bucles extracelulares; en el mayor de estos bucles se fijan los complejos de fusión del esperma. Las microvellosidades se encuentran por toda la superficie del oocito, exceptuando la zona por donde se expulsará el segundo cuerpo polar. Al fusionarse las membranas del oocito y del esperma se libera, del espermatozoide, una molécula de superficie que parece ser una proteína de 30-100 kDA llamada “oscilina” o factor espermático, que activa el oocito produciendo la liberación de Ca++ en forma de ondas (véase Figura 5). Cada vez que se sucede una onda de Ca++, cambia su concentración intracitoplasmática de 50 a 600-1000 nmol/litro.
Estas ondas se dan periódicamente cada 2 - 30 minutos dependiendo de la especie y pueden durar varias horas. El inicio de las ondas de Ca++ induce la activación del oocito que termina con la formación de los pronúcleos. En los cambios que ocurren en el oocito para que se libere el Ca++ intracelular y se produzcan los eventos corriente abajo, podrían estar involucradas las quinasas de la familia de las Src (Src-K); entre éstas la c-Fyn, y la c-Yes que están distribuidas en la corteza del oocito (véase Figura 5).
La c-Fyn también se encuentra alrededor del huso mitótico; la activación de esta quinasa resulta en la activación de la fosfolipasa C (PLC) que cataliza la hidrólisis del PIP2 a DAG e IP3. IP3 actúa en el receptor de IP3 (IP3R) que se encuentra en el retículo endoplásmico y hace liberar el calcio. El mismo calcio participa en su autorregulación y junto con otros moduladores del receptor de IP3 induce las oscilaciones intracelulares descritas, y que tienen como objetivo la activación del oocito.
Sin embargo, cómo se activan las Src-K es una pregunta que se permanece sin respuesta.
Glosario: abreviaturas usadas en las figuras
MP: membrana plasmática
MAE: membrana acrosomal externa
Ca++: ión calcio
PTP: Proteína tirosina fosfatada
PSP: proteína serina fosfatada
PKA1:Proteína quinasa A1
AMPc: adenosin monofosfato cíclico
ATP: adenosin trifosfato
ADP: adenosin difosfato
Pi: fósforo inorgánico
HCO3: ión bicarbonato
sAC: adenilato ciclasa activada por bicarbonato
Na+: ión Sodio
CO2-: ión dioxido de carbono
Cl-: ión cloro
EGF: factor de crecimiento epidermal
PLC γ: Fosfolipasa C gamma
PIP2: Fosfatidil inositol difosfato
DAG: Diacil glicerol
IP3: Inositol trifosfato
GALtasa:Galactosil transferasa
TK: tirosina quinasa
G1: proteína G
CCE: Calcio capacitante
CaM: calmodulina
CaMK: calmodulina quinasa
REnd: retículo endoplásmico
Ubiq: ubiquitina
MPF: factor promotor de la maduración
CD9: receptor familia citoquinas
DE: proteína epididimal
GPI: glicosil fosfatidil inositol, proteína de anclaje
AKAP: Proteínas de dominios de fijación de PKA
PAN: Péptido Atrial Natriurético.
PAS: Peptido Activador de Esperma.
ON: Óxido Nítrico.
ACm: Adenil Ciclasa de Membrana.
GCm: Guanil Ciclasa de membrana.
CaM: Calmodulina.
ACs: Adenil Ciclasa Soluble.
PKA: Proteína quinasa dependiente de AMPc
PTK: Proteina Tirosina Quinasa.
CCp: Canal de Calcio Putativo.
PLC Fosfolipasa C
IP3: Inositol Trifosfato.
RCaIC: Reserva de Calcio Intracitoplasmático
P4: progesterona
SAAF: Factor espermático de activación y atracción (sperm activating and attracting factor)
PASA: péptido activador de espermas
GCm: Guanil Ciclasa ligada a membrana
hOR17-4: factores odorantes
GCs: guanil ciclasa soluble
ON: óxido nítrico
ACm: adenilciclasa de dominio transmembranal
ACs: adenil ciclasa soluble
CatSperm: canales catiónicos del esperma
Summary
The sperm cell: from eyaculation to fertilization During the last recent years there has been a great increase of information regarding the molecular mechanisms involved in the maturation of the male gamete as well as its progression from the gonad up to fertilizing the oocyte. Along this way the sperm completes its maturation adding some molecular components; additionally along this trail take place all the processes leading to activation of the sperm for the entrance into the oocyte to initiates molecular cascades for the formation of a zygote. This article, based on updated literature proposes a model that integrates known cellular and molecular interactions and pinpoints some steps still requiring further research.
Key words: acrosome reaction, activation, adhesion, capacitation, fusion.
Referencias
1. Inaba K. Molecular architecture of the sperm flagella: molecules for motility and signaling. Zoolog Sci 2003; 20: 1043-1056. [ Links ]
2. Urner F, Sakkas D. Protein phosphorylation in mammalian spermatozoa. Reproduction 2003; 125: 17-26. [ Links ]
3. Sakkas D, Leppens-Luisier G, Lucas H, Chardonnens D, Campana A, Franken DR,. et.al. Localization of tyrosine phosphorylated proteins in human sperm and relation to capacitation and zona pellucida binding. Biol Reprod 2003; 68: 1463-1469. [ Links ]
4. Vijayaraghavan S, Mohan J, Gray H, Khatra B, Carr DW. A role for phosphorylation of glycogen synthase kinase-3alpha in bovine sperm motility regulation. Biol Reprod 2000; 62: 1647-1654. [ Links ]
5. Galindo BE, Nishigaki T, Rodriguez E, Sanchez D, Beltran C, Darszon A. Speract-receptor interaction and the modulation of ion transport in Strongylocentrotus purpuratus sea urchin sperm. Zygote 2000;8 Suppl 1:S20-1. [ Links ]
6. Galindo BE, Beltran C, Cragoe EJ, Jr., Darszon A.Participation of a K(+) channel modulated directly by cGMP in the speract-induced signaling cascade of strongylocentrotus purpuratus sea urchin sperm. Dev Biol 2000;221:285-294. [ Links ]
7. Tardif S, Lefievre L, Gagnon C, Bailey JL. Implication of cAMP during porcine sperm capacitation andprotein tyrosine phosphorylation. Mol Reprod Dev 2004; 69:428-435. [ Links ]
8. Ho HC, Suarez SS. Hyperactivation of mammalian spermatozoa: function and regulation. Reproduction 2001;122:519-526. [ Links ]
9. Baker MA, Lewis B, Hetherington L, Aitken RJ. Development of the signalling pathways associated with sperm capacitation during epididymal maturation. Mol Reprod Dev 2003; 64 :446-457. [ Links ]
10. Baker MA, Krutskikh A, Aitken RJ. Biochemical entities involved in reactive oxygen species generation by human spermatozoa. Protoplasma 2003; 221:145-151. [ Links ]
11. Travis AJ, Kopf GS. The role of cholesterol efflux in regulating the fertilization potential of mammalian spermatozoa. J Clin Invest 2002;110:731-736. [ Links ]
12. Williams CJ. Signalling mechanisms of mammalian oocyte activation. Hum Reprod Update 2002;8:313-321. [ Links ]
13. Brewis IA, Wong CH. Gamete recognition: sperm proteins that interact with the egg zona pellucida. Rev Reprod 1999;4:135-142. [ Links ]
14. Flesch FM, Gadella BM. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim Biophys Acta 2000;1469:197-235. [ Links ]
15. White D, Weerachatyanukul W, Gadella B, Kamolvarin N, Attar M, Tanphaichitr N. Role of sperm sulfogalactosylglycerolipid in mouse sperm-zona pellucida binding. Biol Reprod 2000;63:147-155. [ Links ]
16. Gadella BM, Harrison RA. The capacitating agent bicarbonate induces protein kinase A-dependent changes in phospholipid transbilayer behavior in the sperm plasma membrane. Development 2000;127: 2407-2420. [ Links ]
17. Sun QY, Breitbart H, Schatten H. Role of the MAPK cascade in mammalian germ cells. Reprod Fertil Dev 1999;11:443-450. [ Links ]
18. Breitbart H, Naor Z. Protein kinases in mammalian sperm capacitation and the acrosome reaction. RevReprod 1999; 4:151-159. [ Links ]

