










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.21 no.4 Medellín Oct./Dec. 2008
Artículos originales
Actividad de glutatión peroxidasa y superóxido dismutasa en sangre, plasma sanguíneo y plasma seminal en toros Normando¶
Activity of glutathione peroxydase and superoxide dismutase in whole blood, blood plasma and seminal plasma in normande bulls
Atividade de superóxido dismutase e glutationa peroxidase no sangue, plasma e plasma seminal em touros normando
Néstor A Villa1*, MVZ, MS; Wilson Moreno1, MVZ; Alejandro Ceballos1,2, MVZ, MS.
1Departamento de Sistemas de Producción, Grupo de Investigación Salud Productiva Veterinaria,
Universidad de Caldas, A.A. 275. Manizales, Colombia.
2Atlantic Veterinary College, Univeristy of Prince Edward Island. Charlottetown, PE C1A4P3 Canadá.
(Recibido: 26 noviembre, 2007; aceptado: 28 noviembre, 2008)
Resumen
Con el objeto de evaluar la actividad de las enzimas antioxidantes glutatión peroxidasa (GPx; EC 1.11.1.9), y superóxido dismutasa (SOD; EC 1.15.1.1) en sangre, plasma y plasma seminal y su relación con las características seminales: volumen, movilidad individual y en masa, concentración, relación vivos y muertos y prueba hiposmótica (PH), se tomaron muestras de sangre y semen en 20 toros Normando en la cordillera central del Departamento de Caldas (Colombia). De cada toro se tomó una muestra de semen mediante electroeyaculador y una muestra de sangre de la vena coccígea media. Los datos fueron analizados mediante un modelo lineal. La actividad sanguínea de GPx fue 80 ± 48 U/g Hb, 0.09 ± 0.1 U/ml y 0.9 ± 0.7 U/ml en sangre, en plasma sanguíneo y plasma seminal, respectivamente, sin tener correlación con las características seminales ni con la PH (p>0.05). La actividad de SOD fue 940 ± 374 U/g Hb, 120 ± 73 U/ml, y 5.1 ± 2.6 U/ml, en sangre, plasma sanguíneo y en plasma seminal, respectivamente. Una tendencia a una correlación negativa entre SOD en plasma seminal y la PH (p = 0.06) fue hallada, sin tener correlación con las características seminales (p>0.05). La actividad de ambas enzimas en sangre sugiere un subconsumo de Se, Cu y Zn. La actividad enzimática en sangre y plasma no se vio reflejada en la capacidad de defensa enzimática antioxidante del plasma seminal, lo que sugiere la presencia de isoenzimas no determinadas o compartimientos independientes entre los plasmas sanguíneo y seminal.
Palabras clave: calidad espermática, glutatión peroxidasa, minerales en bovinos, prueba hiposmótica, superóxido dismutasa.
Summary
The aim of the study was to evaluate the activity of the antioxidant enzymes glutathione peroxidase (GPx; EC 1.11.1.9), and superoxide dismutase (SOD; EC 1.15.1.1) in blood, blood plasma and seminal plasma, and to correlate them with seminal characteristics (i.e. volume, individual and total motility, concentration, ratio of live and death cells, and hyposmotic test) in Normande bulls. Semen and blood samples were collected from each bull (n = 20) from farms located in the high Andes mountains in Caldas State (Colombia). Data were analyzed by a linear regression model. The blood activity of GPx was 80 ± 48 U/g Hb, 0.09 ± 0.1 U/ml, and 0.9 ± 0.7 U/ml, in blood, blood plasma and seminal plasma, respectively. There was no relationship to either seminal characteristics or hyposmotic test (p>0.05). The activity of SOD was 940 ± 374 U/g Hb, 120 ± 73 U/ml, and 5.1 ± 2.6 U/ml, in blood, blood plasma, and seminal plasma, respectively. There was a negative trend (p = 0.06) in the relationship of SOD activity in seminal plasma to hyposmotic test; however, there was no association to other seminal characteristics (p>0.05). The blood activity of both enzymes suggested a suboptimal intake of Se, Cu, and Zn, and it was not translated into an adequate antioxidant defense of the seminal plasma. It suggested the activity of undetermined isoenzymes or and independency between blood and testicle compartments.
Key words: bovine minerals, glutathione peroxidase, hyposmotic test, sperm quality, superoxide dismutase.
Resumo
Com o objetivo de avaliar a atividade de enzimas antioxidantes glutationa peroxidase (GPx; EC 1.11.1.9), e superóxido dismutase (SOD; EC 1.15.1.1) sangue e plasma seminal plasma, e sua relação com as seguintes características seminais: volume, massa e de mobilidade individual, a concentração, relação vivos e mortos ela prova hiposmótica (PH), amostras de sangue e de sêmen de 20 touros Normando na cordilheira central do Estado de Caldas (Colombia) foram tomadas. Em cada touro teve uma amostra de sêmen através electroeyaculador e uma amostra de sangue da veia coccígea média. Os dados foram analisados usando um modelo linear. A atividade de GPx foi 80 ± 48 U/g Hb, 0.09 ± 0.1 U/ml e 0.9 ± 0.7 U/ml, na sangue, no plasma sanguíneo e plasma seminal, respectivamente, sem qualquer correlação com as características seminais ou com a pH (p>0.05). A atividade da SOD foi 940 ± 374 U/g Hb, 120 ± 73 U/ml, y 5.1 ± 2.6 U/ml, no sangue, no plasma sanguíneo e plasma seminal, respectivamente. Uma tendência para uma correlação negativa entre SOD no plasma seminal e PH (p = 0.06) foi encontrada, sem correlação com as características seminais (p>0.05). A atividade de duas enzimas no sangue sugere-se de um subconsumo, Cu e Zn. A atividade da enzima no plasma sangüíneo não foi reflectiu na capacidade de defesa enzimática antioxidante do plasma seminal, o que sugere a presença de isoenzimas não determinadas ou compartimentos separados entre o plasma sanguíneo e plasma seminal.
Palavras chave: a glutationa peroxidase, esperma qualidade, hiposmótica teste, minerais em bovinos, superóxido dismutase
Introducción
Los antioxidantes han sido agrupados en enzimáticos y no enzimáticas, y son las sustancias capaces de controlar en el organismo la producción de radicales libres (RL) que se generan como consecuencia del metabolismo aerobio, ya sea secuestrando el RL o estabilizándolo (14). Para contrarrestar los RL existe en primera instancia el sistema antioxidante enzimático, que incluye la seleno-enzima (GPx; 1.11.1.9) que actúa fundamentalmente reduciendo el peróxido de hidrógeno (H2O2), y la superóxido dismutasa (SOD; 1.15.1.1) que actúa sobre el anión superóxido (O2•-) transformándolo en un radical secundario (ej. H2O2) para una posterior acción de la GPx (14, 33). Estas enzimas se encuentran entre los principales antioxidantes presentes en el plasma seminal en humanos (12, 13, 27) y su deficiencia ha sido asociada con una posible aparición de signos centinela sugerentes de daño espermático (33).
La producción de RL en semen ha sido asociada con una disminución de la calidad espermática. Así, se han descrito alteraciones como baja movilidad, disminución en la capacidad de fusión oocito-espermatozoide y por lo tanto alteraciones de la fertilidad (27, 33). El H2O2 ha sido reconocido como uno de los RL más tóxicos para el semen (13). Los espermatozoides bovinos están pobremente adaptados al metabolismo de H2O2 y factores como la criopreservación disminuyen significativamente la cantidad de SOD hasta en un 50%, enzima antioxidante importante en el metabolismo y la presentación de este radical a otros antioxidantes (2).
Las enzimas GPx y SOD son metaloenzimas, lo que significa que dependen de la presencia de minerales como selenio (Se), cobre (Cu), zinc (Zn) y manganeso (Mn), que actúan como catalizadores para su síntesis (1, 25). A principios de la década de los 80, se reportó que la suplementación con Se no afectaba la concentración o la viabilidad de los espermatozoides (29); pese a ello, estudios provenientes de este mismo grupo demostraron que la suplementación con Se mejoraba la función reproductiva, como el número de espermatozoides por óvulo fertilizado en vacas suplementadas (30). Recientemente, se ha demostrado una correlación positiva entre la actividad de GPx y la movilidad espermática, mientras que fue negativa con el número de espermatozoides vivos (34).
El efecto benéfico de la suplementación con Se ha sido demostrado en estudios realizados en otras especies. Así, en humanos se ha demostrado la necesidad (9, 20) y el efecto benéfico de suplementar con Se sobre la función espermática, donde se ha visto en pacientes subfértiles suplementados un aumento significativo de la concentración del mineral en plasma seminal y la movilidad espermática, y por lo tanto la oportunidad de una concepción exitosa; pese a ello, los resultados obtenidos fueron variables (28). Igualmente, el incremento de la actividad de SOD ha demostrado su beneficio en la reducción del daño oxidativo sobre los espermatozoides (31).
En la zona del trópico alto del Viejo Caldas (Colombia), deficiencias de Se han sido detectadas, establecidas mediante una baja actividad de GPx (7). También han sido obtenidos valores para SOD que sirven como guía para determinar el balance metabólico nutricional de otros minerales como Cu y Zn (7, 16), los que fueron considerados como base para la interpretación de valores en estudios futuros.
En consonancia con lo anterior y, debido a la importancia económica que tiene la raza normando para la zona del trópico alto en el Viejo Caldas y el impacto del toro sobre los índices reproductivos, el objetivo de este estudio fue determinar la actividad de las enzimas antioxidantes GPx y SOD en diferentes compartimientos y su eventual relación con la calidad espermática.
Materiales y métodos
Tipo de estudio
Este fue un estudio observacional transversal realizado en la zona central del Departamento de Caldas (Colombia), en una zona caracterizada por una altitud de 3550 msnm, con una temperatura media de 9 ºC y una precipitación media anual de 1640 mm. Los sistemas de producción predominantes son cultivos de papa (Solanum tuberosum l.) y la ganadería extensiva o extensiva con suplementación, con base en la raza normando (4).
Animales
Siete predios fueron seleccionados con base en la disponibilidad de tener toros adultos de la raza normando, aptos para la reproducción; es decir, que tuvieran hijos(as) nacidos(as) y vacas en gestación al momento del estudio. Los toros (n = 20) tenían una edad promedio de 2.8 ± 1.2 años, no tuvieron reposo sexual previo a la evaluación andrológica. La base de la alimentación era pradera natural, donde predominaban los pastos riqueza (Anthoxanthum odoratum) y falsa poa (Holcus lanantus); además, recibían un suplemento mineral y agua a voluntad.
Muestras
Los toros fueron evaluados clínicamente para descartar la presencia de alteraciones morfológicas o anormalidades en el tracto genital. A cada toro se le midió la circunferencia escrotal con cinta métrica, y se le tomó una muestra de semen mediante el uso de un electroeyaculador (ElectroJac IV®, Neogen-Ideal Instruments, Lexington, KY, USA). El examen del semen incluyó volumen, movimiento en masa, movilidad individual progresiva, concentración y relación vivos/muertos. Además, se practicó a cada una de las muestras de semen una prueba hiposmótica (PH), utilizando una solución de citrato de sodio a 100 mOsm/l e incubando 0.1 ml de semen en 0.9 ml de la solución a 37 °C/30 min, según el protocolo descrito (35). Luego, el semen se centrifugó a 3000 rpm/15 min, para obtener el plasma, el cual se conservó congelado a -20°C hasta el día de la determinación enzimática.
Después de la colección del semen, a cada toro se le tomó una muestra de sangre mediante venopunción coccígea usando tubos al vacío con heparina. Posteriormente, se determinó la concentración de hemoglobina y una alícuota de la sangre se centrifugó para obtener el plasma, utilizando la sangre restante para la medición de GPx y SOD. Para la determinación de GPx la sangre se conservó diluida, mientras que para la determinación de SOD los eritrocitos fueron separados mediante centrifugación y lavados seriados con solución salina. Ambas diluciones fueron conservadas a -20 °C hasta el análisis.
La actividad de GPx fue determinada mediante un método cinético compuesto NADPH-dependiente, tanto para sangre como plasma sanguíneo y seminal, mientras que la actividad de SOD fue hecha mediante un método colorimétrico de punto final (6, 7). Las lecturas fueron realizadas en un fotómetro automático Bio Systems BTS-330 (BioSystems, Barcelona, España), empleando para los análisis reactivos comerciales (Ransel® y Ransod®; Randox Laboratories, Crumlin, UK). La actividad sanguínea de ambas enzimas fue expresada en U/g de Hb (U/g Hb) y en plasma sanguíneo y seminal en unidades por mililitro (U/ml). Una actividad inferior a 60 U/g Hb señalaría una deficiencia metabólica nutricional de Se (6), mientras que una actividad de SOD inferior a 1000 U/g Hb estaría indicando un consumo inadecuado de Cu y Zn (16).
Análisis estadístico
Los datos se describen mediante la obtención del promedio (![]() ), desviación estándar (DE), rango e intervalo de confianza (IC) al 95%. Inicialmente se establecieron asociaciones incondicionales para las variables en estudio y posteriormente se evaluó el efecto de la actividad enzimática en los diferentes compartimientos sobre aquellas características seminales con las cuales hubo una asociación significativa (p<0.10), mediante un modelo lineal general (8). Los análisis fueron realizados usando el programa estadístico STATA versión 10.1 (StataCorp, College Station, TX, USA).
), desviación estándar (DE), rango e intervalo de confianza (IC) al 95%. Inicialmente se establecieron asociaciones incondicionales para las variables en estudio y posteriormente se evaluó el efecto de la actividad enzimática en los diferentes compartimientos sobre aquellas características seminales con las cuales hubo una asociación significativa (p<0.10), mediante un modelo lineal general (8). Los análisis fueron realizados usando el programa estadístico STATA versión 10.1 (StataCorp, College Station, TX, USA).
Resultados
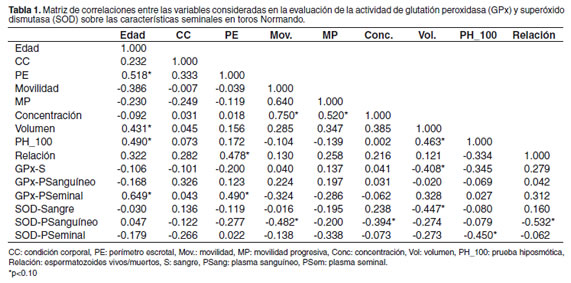
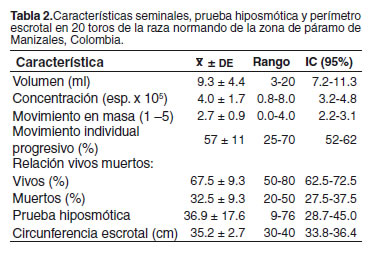
En la tabla 1 se presenta la matriz de correlaciones entre las variables estudiadas, así como aquellas que tuvieron un efecto significativo (p<0.10) en el modelo. Estas variables fueron posteriormente incluidas en el modelo lineal para evaluar su efecto sobre las características seminales. En la tabla 2 se resumen las características de la evaluación andrológica de los toros del estudio. En el análisis de correlación se encontró que la edad de los toros estaba asociada en forma significativa con el perímetro escrotal y algunas de las características seminales como volumen y la PH. Asimismo, estaba asociado positivamente el perímetro escrotal con la relación de espermatozoides vivos y muertos (véase Tabla 1).
Glutatión peroxidasa
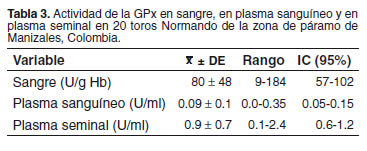
En sangre fue hallada una actividad de GPx inferior a 60 U/g Hb en ocho de los 20 toros evaluados, mientras que cinco de ellos presentaron valores marginales (61-100 U/g Hb). No hubo una correlación significativa entre la actividad en sangre con la actividad en los otros compartimientos (p>0.05). En la tabla 3 se describen los valores para la actividad de GPx encontrados en sangre y plasma sanguíneo y seminal. No se observaron correlaciones significativas entre la actividad de la enzima, las características seminales y la prueba hiposmótica, excepto una tendencia (p = 0.07) a la disminución del volumen espermático en los toros con mayor actividad de GPx en sangre (véase Tabla 1).
Superóxido dismutasa
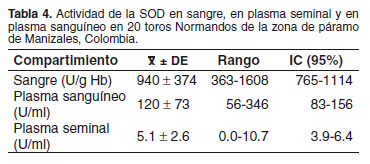
En 13 toros se encontraron valores de SOD por debajo de 1000 U/g Hb, mientras que los restantes tuvieron valores superiores, siendo el máximo 1608 U/g Hb. Igualmente, no se observaron correlaciones entre la actividad sanguínea de SOD y la actividad en los demás compartimientos evaluados. La actividad promedio para SOD encontrada en sangre, plasma sanguíneo y seminal se presenta en la tabla 4. Al igual que para la actividad sanguínea de GPx, se observó una tendencia (p = 0.05) a tener volúmenes más bajos en los toros con mayor actividad de SOD (véase Tabla 1). La actividad de SOD en sangre no estuvo correlacionada con otras variables.
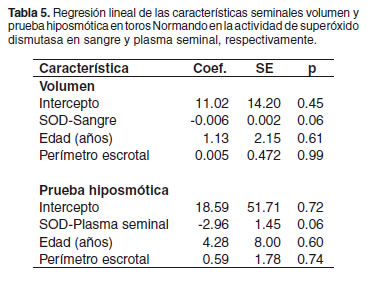
Las variables significativas en la asociación incondicional (p<0.10) fueron utilizadas posteriormente en un modelo lineal. La edad de los toros, así como la circunferencia escrotal, fueron incluidas como covariables. La actividad sanguínea de SOD mostró una tendencia hacia una relación negativa con el volumen, donde por cada unidad de incremento en la actividad enzimática en sangre, el volumen espermático decayó en 0.05 ml. Asimismo, la actividad de SOD en el plasma seminal mostró la tendencia hacia una relación negativa con el resultado de la PH, donde por cada unidad de incremento en la actividad enzimática, el porcentaje de dilatación decayó en 3.2 unidades. La edad del toro, así como la circunferencia escrotal no presentaron un efecto significativo en ambos modelos (véase Tabla 5).
Discusión
La concentración espermática observada en este estudio fue baja. Este fenómeno pudo deberse a la toma de la muestra de semen con el electroeyaculador, ya que su uso generalmente produce un mayor volumen seminal y una menor concentración. La dilatación espermática observada 36.9 ± 17.6% (véase Tabla 2) indica que los espermatozoides tuvieron una buena capacidad para reaccionar en el medio hiposmótico, lo que se debe a la presencia de mecanismos iónicos activos en la membrana celular, como la bomba Na-K (3).
Glutatión peroxidasa
La actividad sanguínea de GPx fue baja, y una actividad enzimática inferior a 130 U/g Hb es compatible con una concentración sanguínea inferior al valor señalado como mínimo en sangre para bovinos, lo que sugiere un balance metabólico nutricional de Se inadecuado (5, 6); esto significa que los animales evaluados podrían estar consumiendo dietas con una concentración de Se inferior a 0.3 ppm, valor que ha sido fijado como la concentración adecuada en la ración para bovinos (21).
En otros estudios realizados con toros de la raza brahman (37), se ha observado una actividad de GPx en sangre cercana a 300 U/g Hb. Así mismo, estudios realizados en Caldas (7) han reportado que la actividad de GPx en sangre es más baja en novilla y se observa una mayor frecuencia de valores bajos en zonas geográficas por encima de 2000 msnm. La baja actividad de GPx se debe a un aporte insuficiente de Se en la dieta, ya que la relación planta-animal indica que un bajo contenido de minerales en la pradera se verá reflejado en una concentración de Se inadecuada, lo que a su vez se puede establecer mediante una disminución en la actividad de GPx, dadas las características estructurales de la enzima donde hay cuatro átomos de Se por cada mol de enzima (5, 6, 39).
La actividad de GPx fue baja en el plasma sanguíneo y no se relacionó con la actividad en los eritrocitos. El plasma podría considerarse como un compartimiento de distribución del Se donde no se almacena este mineral, mientras que los eritrocitos lo captan desde el plasma, tienen un compartimiento de movilización más lenta y poseen aproximadamente un 70% del Se total de la sangre (19). La actividad de GPx en plasma seminal de los toros en estudio fue inferior a la hallada en toros brahman en pastoreo (36). Estudios realizados en otras especies, como los porcinos, han hallado valores hasta ocho veces superiores a los obtenidos en este estudio (26), lo que posiblemente esté relacionado con una mayor absorción del Se comparado con lo observado en rumiantes (5, 19).
Considerando que la mayor parte de los animales en este estudio tuvo una actividad de GPx baja-marginal, es probable que la actividad sanguínea no haya sido suficiente para trasladarse a un mejoramiento de las características seminales, lo que se corroboró al observar asociaciones no significativas entre la GPx y la evaluación andrológica (véase Tabla 1). Sin embargo, en otras especies, como humanos, las evidencias indican que hay una asociación positiva entre la suplementación con Se y el mejoramiento de las características seminales (17, 28). Aún así, es posible observar resultados contradictorios tanto en bovinos (29) como en humanos (15).
Pese a lo anterior, el Se es importante en la fisiología espermática, cuya participación va desde la protección contra el daño oxidativo (23) hasta su efecto directo en la espermatogénesis al formar parte de la cubierta mitocondrial del espermio (9, 20). Una enzima fundamental en este proceso es la fosfolípido hidroperóxido glutatión peroxidasa (GPx4; EC 1.11.1.12), cuyo contenido y actividad en el testículo son altos, habiéndose señalado como deficitaria en pacientes con alteraciones de la movilidad espermática (10). Lo anterior sugiere la necesidad de evaluar las características seminales en toros que posean un balance nutricional de Se adecuado o posterior a la suplementación y emplear para ello biomarcadores más específicos relacionados con la fisiología del espermatozoide.
Superóxido dismutasa
Al igual que lo observado para la actividad de GPx, se encontró una alta proporción de animales con valores inferiores a los reportados para toros de otras razas (37) o para bovinos lecheros en Caldas (7). Lo anterior podría ser debido a deficiencias en el aporte de nutrientes o a una diferencia en los suplementos minerales utilizados en las diferentes regiones del departamento de Caldas, donde se han realizado los estudios. Pese a contar con algunos antecedentes (16), es necesario realizar más estudios que permitan correlacionar la actividad de la enzima con el consumo de los minerales precursores de la misma.
La actividad enzimática de SOD en el plasma sanguíneo fue 120 ± 73 U/ml; sin embargo, no se encontraron antecedentes en la literatura en esta u otras especies que permitieran una interpretación de este resultado. Al igual que para la GPx la mayor parte de la actividad de esta enzima debería encontrarse en los eritrocitos (1) y no el plasma, ya que es en esta célula donde se requiere la acción antioxidante. La actividad de SOD en plasma seminal de los toros en estudio (véase Tabla 4) fue similar a la obtenida en toros de la raza brahman (37). No obstante, los valores observados tanto en este estudio como en toros brahman fueron inferiores a los obtenidos previamente en toros en pastoreo (11.8 ± 9.4 U/ml) y suplementados (20.4 ± 12.7 U/ml) (36). Los resultados para la actividad de SOD en el plasma seminal obtenidos en otras especies han arrojado valores variables. En porcinos se han obtenido valores en machos fértiles de 2.1 ± 1.8 U/ml (26), lo que es inferior a lo encontrado en otras especies. En estudios con plasma seminal humano (11), se halló una actividad enzimática de 6.9 ± 2.8 U/ml, determinándose que posiblemente la SOD se origina del espermatozoide debido a la correlación observada entre la actividad de la SOD y la concentración espermática (r = 0.27, p<0.05). Pese a esta asociación positiva, este efecto no se encontró en este estudio.
Igualmente, se ha establecido la actividad de SOD en pacientes con alteraciones de la función espermática: en un estudio realizado en pacientes oligozoospérmicos y astenozoospérmicos, se encontró una actividad de 11 ± 4 U/ml en plasma seminal, lo cual fue bajo e indicó la presencia de niveles incrementados de O2•-; como una posible causa se argumentó el estrés oxidativo (32). En otro estudio en humanos se observó una actividad de SOD en plasma seminal sensiblemente más elevada (366.8 ± 20.9 U/ml), pero no se correlacionó con la concentración y movilidad espermáticas (18). La determinación de la actividad de SOD en plasma seminal ha demostrado ser un indicador útil en la evaluación de la defensa antioxidante del semen. Resultados de investigaciones en humanos sugieren que las secreciones prostáticas son una importante fuente de defensa contra O2•- (11). Estos mismos autores sugieren que la actividad de la enzima en hombres infértiles con infecciones de glándulas sexuales accesorias tiene una baja actividad comparada con lo observado en hombres infértiles sin signos de infección, lo que indica que el sistema de defensa antioxidante capaz de neutralizar el O2•- depende de la actividad secretoria de las glándulas sexuales accesorias (11). Lo anterior ha sido corroborado en otros estudios que también sugieren que la protección antioxidante del plasma seminal es de origen prostático (24, 38). Igualmente, se ha señalado que la Cu-Zn-SOD en los órganos genitales masculinos juega un papel importante en la diferenciación celular, incluyendo la espermatogénesis (22).
En este estudio se encontraron correlaciones negativas entre la actividad de SOD en plasma seminal y la PH (véase Tabla 5), lo que sugiere que una mayor dilatación espermática debería esperarse en animales con una menor actividad de SOD en el plasma seminal, posiblemente ello se deba a una depleción de la actividad enzimática en la protección antioxidante para mantener la integridad de la membrana plasmática, o el consumo de Cu y Zn no fue suficiente para lograr un mayor aumento de la actividad enzimática en el plasma seminal, ya que hubo una alta frecuencia de valores inferiores a 1000 U/g en la actividad sanguínea, que sugiere un consumo inadecuado de estos minerales. No obstante, ambas hipótesis requieren la realización de estudios adicionales.
La falta de una correlación entre la actividad sanguínea con la actividad en el plasma seminal desde el punto de vista estadístico, no descarta que haya algún tipo de asociación biológica entre ellas; pero, es difícil evaluarlo bajo las condiciones de este estudio donde diversos factores, entre ellos el subconsumo de minerales, pudieron causar algún efecto no determinado en esta asociación. Cabe también la hipótesis que los compartimientos sanguíneo y testicular sean independientes en cuanto al metabolismo de Cu y Zn, pudiendo el testículo tener isoenzimas específicas, determinaciones que escapan al objetivo de este estudio.
La falta de más antecedentes en la literatura respecto de los estudios con estas enzimas en la función espermática en toros, hace difícil la adecuada interpretación de los resultados obtenidos; sin embargo, los antecedentes obtenidos en otras especies pueden ser una guía, sin dejar de reconocer las diferencias que existen en el metabolismo mineral entre especies.
Agradecimientos
Los autores agradecen a los propietarios de las ganaderías que facilitaron los animales para esta investigación y a la Vicerrectoría de Investigaciones y Postgrados de la Universidad de Caldas.
Referencias
1. Bannister J, Bannister W, Woo E. Bovine erythrocyte cupro-zinc protein. 1. Isolation and general characterization. Eur J Biochem 1971; 18:178-186. [ Links ]
2. Bilodeau JF, Chatterjee S, Sirard MA, Gagnon C. Levels of antioxidant defenses are decreased in bovine spermatozoa after a cycle of freezing and thawing. Mol Reprod Dev 2000; 55:282-288. [ Links ]
3. Caiza FI, Rigau T, Bonet S, Miró J, Briz M, et al. Subjecting horse spermatozoa to hypoosmotic incubation:effects of ouabain. Theriogenology 1997; 47:765-784. [ Links ]
4. Castro R, Granobles J, Cruz G. Caracterización y tipificación de los sistemas de producción del cultivo de la papa (Solanum tuberosum l.) en la zona Tesorito-Páramo de Letras. Rev Sist Prod 2000; 10:3-20. [ Links ]
5. Ceballos A, Wittwer F. Metabolismo del Se en rumiantes. Arch Med Vet 1996; 28:5-18. [ Links ]
6. Ceballos A, Wittwer F, Contreras PA. Boehmwald H. Actividad sanguínea de glutatión peroxidasa en rebaños lecheros a pastoreo: variación según edad y época del año. Arch Med Vet 1998; 30:13-22. [ Links ]
7. Ceballos A, Correa H, Loaiza J, Villa NA. Actividad sanguínea de glutatión peroxida como indicador del balance metabólico nutricional de selenio en rebaños lecheros de Manizales, Colombia. Rev Colomb Cienc Pecu 2003; 16:19-25. [ Links ]
8. Dohoo I, Martin W, Stryhn H. Veterinary epidemiologic research. Charlottetown, PE: AVC Inc; 2003. [ Links ]
9. Flohé L. Selenium in mammalian spermiogenesis. Biol Chem 2007; 388:987-995. [ Links ]
10. Foresta C, Flohé L, Garolla A, Roveri A, Ursini F, et al. Male fertility is linked to the selenoprotein phospholipid hydroperoxide glutathione peroxidase. Biol Reprod 2002; 67:967-971. [ Links ]
11. Gavella M, Lipovac V, Vucic M, Rocic B. Superoxide anion scavenging capacity of human seminal plasma. Int J Androl 1996; 19:82-90. [ Links ]
12. Griveau JF, Dumont P, Renard J, Callegari JP, Le Lannou D. Reactive oxygen species, lipid peroxidation and enzymatic defence systems in human spermatozoa. J Reprod Fertil 1995; 103:17-26. [ Links ]
13. Griveau JF, Le Lannou D. Reactive oxygen species and human spermatozoa: physiology and pathology. Int J Androl 1997; 20:61-69. [ Links ]
14. Halliwell B, Whiteman M. Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean? Br J Pharmacol 2004; 142:231-255. [ Links ]
15. Iwanier K, Zachara BA. Selenium supplementation enhances the element concentration in blood and seminal fluid but does not change the spermatozoal quality characteristics in subfertile men. J Androl 1995; 16:441-447. [ Links ]
16. Jaramillo S, Villa NA, Pineda AF, Gallego AB, Tabares P, et al. Actividad sanguínea de superóxido dismutasa y glutatión peroxidasa en novillas a pastoreo. Pesq Agropec Bras 2005; 40:1115-1121. [ Links ]
17. Keskes-Ammar L, Feki-Chakroun N, Rebai T, Sahnoun Z, Ghozzi H, et al. Sperm oxidative stress and the effect of an oral vitamin E and selenium supplement on semen quality in infertile men. Arch Androl 2003; 49:83-94. [ Links ]
18. Kobayashi T, Miyazaki T, Natori M, Nozawa S. Protective role of superoxide dismutase in human sperm motility:superoxide dismutase activity and lipid peroxide in human seminal plasma and spermatozoa. Hum Reprod 1991; 6:987-991. [ Links ]
19. Krishnamurti CR, Ramberg CF Jr, Shariff MA. Kinetic modeling of selenium metabolism in nonpregnant ewes. J Nutr 1989; 119:1146-1155. [ Links ]
20. Maiorino M, Roveri A, Ursini F, Brigelius-Flohé R, Flohé L. Selenium in male reproduction. In: Hatfield DL, Berry MJ, Gladyshev VN (Eds). Selenium: Its molecular biology and role in human health. 2ª ed. New York: Springer; 1995. p. 323-331. [ Links ]
21. National Research Council (NRC). Nutritional requirements of dairy cattle. 7a ed. Washington DC: National Academy Press; 2001. [ Links ]
22. Nonogaki T, Noda Y, Narimoto K, Shiotani M, Mori T, et al. Localization of Cu-Zn-superoxide dismutase in the human male genital organs. Hum Reprod 1992; 7:81-85. [ Links ]
23. Oldereid NB, Thomassen Y, Purvis K. Selenium in human male reproductive organs. Hum Reprod 1998; 13:2172-2176. [ Links ]
24. Peeker R, Abramsson L, Marklund SL. Superoxide dismutase isoenzymes in human seminal plasma and spermatozoa. Mol Hum Reprod 1997; 3:1061-1066. [ Links ]
25. Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, et al. Selenium: biochemical role as a component of glutathione peroxidase. Science 1973; 179:588-590. [ Links ]
26. Sánchez, L. Determinación del estado antioxidante en sangre y plasma seminal en cerdos (Tesis de pregrado), Facultad de Ciencias Agropecuarias, Universidad de Caldas, Manizales, Colombia 2000. [ Links ]
27. Sanocka D, Kurpisz M. Reactive oxygen species and sperm cells. Reprod Biol Endocrinol 2004; 2:12-18. [ Links ]
28. Scott R, MacPherson A, Yates RW. The effect of oral selenium supplementation on human sperm motility. Br J Urol 1998; 82:76-80. [ Links ]
29. Segerson EC, Johnson BH. Selenium and reproductive function in yearling Angus bulls. J Anim Sci 1980; 51:395-401. [ Links ]
30. Segerson EC, Libby DW. Ova fertilization and sperm number per fertilized ovum for selenium and vitamin E-treated charolais cattle. Theriongenology 1982; 17:333-341. [ Links ]
31. Sheweita SA, Tilmisany AM, Al-Sawaf H. Mechanisms of male infertility: role of antioxidants. Curr Drug Metab 2005; 6:495-501. [ Links ]
32. Siciliano L, Tarantino P, Longabardi F, Rago V, De Stefano C, et al. Impaired seminal antioxidant capacity in human semen with hyperviscosity or oligoasthenozoospermia. J Androl 2001; 22:798-803. [ Links ]
33. Tremellen K. Oxidative stress and male infertility-A clinical perspective. Hum Reprod Update 2008; 14:243-258. [ Links ]
34. Vaisberg CN, Jelezarsky LV, Dishlianova B, Chaushev TA. Activity, substrate detection and immunolocalization of glutathione peroxidase (GPx) in bovine reproductive organs and semen. Theriogenology 2005; 64:416-428. [ Links ]
35. Vázquez J, Martínez E, Martínez P. Hypoosmotic swelling of boar spermatozoa compared to other methods for analysing the sperm membrane. Departament of pathology. Theriogenology 1997; 47:913-922. [ Links ]
36. Villa NA, Ceballos A, Correa S, López E, Salazar J. Evaluación del estado antioxidante en plasma seminal de toros brahman (Bos indicus) en pastoreo y suplementados. Rev Colomb Cienc Pecu 1999; 12(Supl):187.a [ Links ]
37. Villa NA, Ceballos A, García G, Franco F. Actividad sanguínea de glutatión peroxidasa y superóxido dismutasa y su correlación con la actividad en plasma seminal de toros brahman (Bos indicus) en pastoreo. Rev Colomb Cienc Pecu 1999; 12(Supl):185.b [ Links ]
38. Yeung CH, Cooper TG, De Geyter M, De Geyter C, Rolf C, et al. Studies on the origin of redox enzymes in seminal plasma and the relationship with results of in-vitro fertilization. Mol Hum Reprod 1998; 4:835-839. [ Links ]
39. Wichtel JJ. A review of selenium deficiency in grazing ruminants. Part 1: new roles for selenium in ruminant metabolism. N Z Vet J 1998; 46:47-52. [ Links ]
¶ Para citar este artículo: Villa NA, Moreno W, Ceballos A. Actividad de glutatión peroxidasa y superóxido dismutasa en sangre, plasma sanguíneo y plasma seminal en toros normando. Rev Colomb Cienc Pecu 2008; 21:537-545.
* Autor para el envío de la correspondencia y la solicitud de separatas: Departamento de Sistemas de Producción, Grupo de Investigación Salud Productiva Veterinaria, Universidad de Caldas, A.A. 275. Manizales, Colombia. E-mail: navilla@ucaldas.edu.co

