










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.22 no.2 Medellín Apr./June 2009
Evaluación de tres protocolos de tratamiento hormonal sobre el diámetro de ovocitos de sabaleta Brycon henni *
Evaluation of three protocols hormone treatment on the size of the oocytes of sabaleta Brycon henni
Avaliação de tres protocolos de tratamento hormonal sob o diámetro dos ovocitos da sabaleta Brycon henni
Gustavo A Lenis1*, Biólogo, cMSc; Luis F Restrepo1, Estadístico Esp ; Pablo E Cruz-Casallas, PhD2 .
1 Grupo de Investigación GRICA. Facultad de Ciencias Agrarias, Universidad de Antioquia, A.A. 1226, Medellín, Colombia. 2 Grupo de Reproducción y Toxicología de Organismos Acuáticos - GRITOX, Instituto de Acuicultura, Facultad de Ciencias Agropecuarias y Recursos Naturales, Universidad de los Llanos, A.A 110, Villavicencio, Meta - Colombia.
(Recibido: 29 octubre, 2008; aceptado: 7 mayo, 2009)
Resumen
El estudio fue realizado en el Municipio de San Carlos (Antioquia, Colombia), por la Facultad de Ciencias Agrarias de la Universidad de Antioquia, en convenio con la Corporación CORNARE, ISAGEN, durante la temporada reproductiva Mayo-Julio de 2008. 12 hembras y 24 machos de sabaleta Brycon henni de aproximadamente 100 g de peso corporal y 20 cm de longitud estándar, provenientes de un lote de 320 reproductores capturados del ambiente natural en Agosto del 2007 y mantenidos en cautiverio en un estanque de tierra de 200 m2, fueron seleccionados por sus características externas de madurez sexual como vientre abultado, papila genital enrojecida y expulsión de semen después de leve presión sobre la cavidad celómica. Además como carácter de dimorfismo sexual se consideró para las hembras la presencia de aleta anal lisa o con espinaciones en máximo el 50% de su longitud, mientras que en los machos es rugosa en toda su extensión. También se tuvieron en cuenta características internas, en las hembras se realizó biopsia ovárica por aspiración de una muestra de ovocitos para observar la posición de sus núcleos y medición del de diámetro.. Las hembras fueron seleccionadas con la condición que presentaran ovocitos con diámetros superiores a 900 µm. Los animales fueron distribuidos para recibir uno de tres tratamientos, T1 Extracto de Hipófisis de Carpa (Argent, USA) (EHC), n=4; T2 Ovopel (Sigma Co., St Louis, Missouri), n=4; T3 Ovaprim (Syndel Laboratories Ltd, Vancouver, BC Canadá), n=4, durante cuatro fechas de muestreo y fertilizadas cada una con el semen de dos machos, inyectados previamente con EHC. 11 de las 12 hembras respondieron a los tratamientos y presentaron diámetros ovocitarios promedio de 1428.8 y 1679.8 (T1), 1531.2 y 1879.2 (T2) y de 1712.8 a 1803.3 (T3), antes y después del tratamiento hormonal, respectivamente. Se observaron diferencias significativas al interior del T1 y T2 (p<0.05), pero no en el T3. La distribución de frecuencias del diámetro de los ovocitos presentó en T1, dos modas antes del tratamiento hormonal y dos después del tratamiento hormonal (bimodal); T2, dos modas antes y una después (bimodal y unimodal) y T3, dos antes y dos después (bimodal). Los porcentajes de fertilización fueron bajos en el T1, pero se presentaron altos porcentajes en T2 y T3 hasta las 12 h de incubación de los huevos.
Palabras clave: diámetro ovocitos, extracto hipofisiario, peces, reproducción.
Summary
The evaluation of three hormone treatments on the diameter size of ovocytes in Sabaleta was conducted during the breeding season from May to July 2008. Twelve Brycon henni females and 24 males, 100 g and a standard length of 24 cm, were selected according to their sexual maturity and external features such as bulging belly, red genital papilla and semen expulsion after softly pressing its coelomic cavity. The fish were selected from a 320 brood fish batch caught in the wild in August 2007 and kept in captivity in a 200 m2 earth pond. The presence of an anal fin with or without spines of a maximum length of 50% was considered as sexual dimorphism in females. Internal characteristics were also considered; e.g., an ovarian aspiration biopsy was performed in females in order to observe its nuclei position and diameter. Ovocytes from selected females were at least 900 µm in diameter. Animals were distributed into three treatment groups for four sampling dates: T1 carp pituitary extract (CPE) (Argent Chemical Laboratories Inc., USA), n = 4; T2 Ovopel (Sigma Co. USA), n = 4; and T3 Ovaprim (Syndel Laboratories Ltd., Canada), n = 4. Each female was fertilized with the semen of two males previously injected with CPE. 11 out of 12 females had a treatment response, and resulting ovocyte average diameters were 1428.8 and 1679.8 (T1), 1531.2 and 1879.2 (T2), and 1712.8 to 1803.3 (T3), before and after hormone treatment, respectively. Significant differences were found within T1 and T2 (p <0.05); however, this was not the case for T3. For T1, frequency distribution of the diameter of ovocytes before the hormonal treatment was bimodal. T2 had two modes before and one after the treatment (bimodal and unimodal), and T3 had two before and two after (bimodal). Fertilization rates were low in T1; nevertheless, T2 and T3 had high rates until 12 h eggs incubation.
Key words: Fish, ovocyte diameter, pituitary extract, reproduction.
Resumo
O estudo foi realizado no Município de San Carlos (Antioquia, Colombia), pela Facultad de Ciencias Agrarias de la Universidad de Antioquia, conjuntamente com a corporação CORNARE, ISAGEN, entre os meses de maio e julho de 2008. Foram utilizadas 12 fêmeas e 24 machos de sabaleta Brycon henni de aproximadamente 100 g de peso corporal e 20 cm de comprimento, provenientes de um lote de reprodutores capturados em ambiente natural em agosto de 2007. Os animais receberam um dos três tratamentos: T1 extrato de hipófise de Hipófise de Carpa (Argent, USA) (EHC), n=4; T2 Ovopel (Sigma Co., St Louis, Missouri), n=4; T3 Ovaprim (Syndel Laboratories Ltd, Vancouver, BC Canadá), n=4, durante quatro datas de amostragem e fertilizadas cada uma com sêmen de dois machos, injetados previamente com EHC. 11 das 12 fêmeas responderam aos tratamentos e apresentaram ovócitos entre 1428.8 e 1679.8 (T1), 1531.2 e 1879.2 (T2) e de 1712.8 a 1803.3 (T3) de diâmetro, antes e depois do tratamento hormonal, respectivamente. Observaram-se diferencias significativas ao interior do T1 e T2 (p<0.05), mas não para o T3. A distribuição de freqüências do diâmetro do ovócito apresentou no T1, duas modas antes do tratamento hormonal e dois depois do tratamento hormonal (bimodal); T2, duas modas antes e 1 depois (bimodal e unimodal) e T3, duas antes e duas depois (bimodal). As porcentagens de fertilização foram baixas em T1, e foram altas em T2 e T3 até as 12 h de incubação dos ovos.
Palavras chave: diâmetro de ovócitos, extrato de hipófises, peixes, reprodução.
* Para citar este artículo: Lenis GA, Restrepo LF, Cruz-Casallas PE. Evaluación de tres protocolos de tratamiento hormonal sobre el diámetro de los ovocitos de sabaleta Brycon henni. Rev Colomb Cienc Pecu 2009; 22: 131-142.
* Autor para correspondencia: Facultad de Ciencias Agrarias, Carrera 75 No 65 -87, Ciudadela Robledo, Universidad de Antioquia, Medellín, Colombia. Email: g-lenis@hotmail.com.
Introducción
La sabaleta (Brycon henni) es un pez migratorio que habita en las cuencas y microcuencas de la región cafetera de Colombia y es tal vez la especie euritérmica más representativa, puesto que se encuentra en aguas limpias y transparentes con temperaturas entre los 18 y 28 ºC y concentraciones de oxígeno disuelto entre 6 y 10 mg/l (11, 12, 34). Sin embargo, estas condiciones ecológicas se han visto deterioradas por contaminación generada por los centros urbanos localizados en la región. Además, las necesidades de agua potable y de energía eléctrica han propiciado la construcción de embalses que afectan los procesos migratorios de la especie con la consecuente disminución de las poblaciones.
La sabaleta es una especie endémica de Colombia y en especial de la cuenca de los ríos Porce y sus afluentes como el Río Grande, Nus, San Rafael, San Carlos, todos localizados en la zona de embalses del Departamento de Antioquia, en donde la especie, a pesar de que aún no ha sido declarada en peligro de extinción, sigue siendo muy apetecida por la calidad de su carne, lo que ha provocado un descenso considerable en sus índices de captura Es también una especie de alta importancia económica, cultural y científica y aún no existen estudios completos sobre su biología, estado actual de las poblaciones, reproducción en el ambiente natural e inducida en ambientes artificiales como los laboratorios de piscicultura.
Según Montoya (31), gran parte de la información sobre B. henni necesita ser revisada, actualizada y ampliada. El estado real de las poblaciones de B. henni es desconocido, necesitándose investigaciones urgentes en este tópico. Además, se requieren investigaciones referentes al crecimiento en cautiverio de la especie y todavía no se logra resolver el problema de producción masiva de alevinos con diferentes finalidades como el repoblamiento o el cultivo en estanques. Aunque se conoce la respuesta positiva a sustancias inductoras como EHC (Argent, USA) y Primogonyl, no se han evaluado aún otras alternativas como Ovaprim® (Syndel laboratories Ltd, Vancouver, BC Canadá) y el Ovopel® (Sigma Chemical Co. St Louis, Missouri), que probablemente tengan una mayor eficiencia sobre el desempeño reproductivo de la especie. Inyecciones e implantes del análogo sintético de hormona liberadora de gonadotropina GnRHa (6, 15), aún no han sido evaluados, para conocer la respuesta comparada con otros inductores ya probados como el EHC. Los niveles de gonadotropina maduracional o LH (18, 32, 42), no han sido determinados en peces del medio natural ni en animales sometidos a procesos de amansamiento en cautiverio.
Para la sabaleta Brycon henni, según los resultados de la investigación realizada en la estación de piscicultura del Nus (Lenis et al, 2008, en revisión), el período de latencia se encuentra entre las 11-13 h, en las que se presentaron los huevos viables con los mayores porcentajes de fertilización, eclosión, sobrevivencia larval y alevinos, pero se deben hacer más investigaciones tomando biopsias en cortos períodos de tiempo para determinar el mejor momento del estrujamiento y relacionarlos con los diámetros ovocitarios y con las dos temporadas reproductivas que presenta la especie, al parecer, coincidentes con los picos de lluvias en la región (11, 14, 26). Los diámetros ovocitarios a partir de la biopsia ovárica y posterior a la hidratación y otros parámetros de desempeño reproductivo (35), aún no han sido determinados para recomendar el mejor momento de desarrollo gonadal que permita iniciar con mayor posibilidad de éxito un programa de reproducción inducida.
Uno de los más importantes problemas en acuicultura es obtener gametos buena calidad, por esta razón los tratamientos hormonales son usados para estimular su maduración en producciones comerciales de ciprínidos, siendo uno de los agentes más comúnmente utilizados el EHC, en algunos casos con adición de Gonadotropina Coriónica Humana (hCG) (39), o con análogos de Hormona Liberadora de Gonadotropina (GnRHa) y algunas veces con antagonistas Dopaminérgicos (DA) (24). Sin embargo, el alto valor comercial del EHC comparados con los análogos de GnRH, además de los resultados exitosos para estimular la ovulación en varias especies de peces (13, 27, 37), hacen más atractivo su uso.
Horvath et al en 1997 citado por Kucharczyk et al (24), propuso un nuevo protocolo para estimular la maduración final y la ovulación en ciprínidos usando análogos de GnRH contenidos en pellets (Ovopel) con peso promedio de 25 mg, que contienen el análogo de mamífero D-Ala 6, Pro 9 NEt-mGnRH en dosis de 18-20 µg y el antagonista de DA metoclopramide en dosis de 8-10 mg (24). En varias especies de peces, la aplicación de Ovopel dio buenos resultados para estimular la ovulación, por ejemplo en Perca fluviatilis L. (23, 25), Leuciscus idus (24), Cyprinus carpio (8, 10, 27), Silurus glanis (7, 9, 37), carpas de la India (13), Lucioperca L. (39) y Abramis brama L. (25), dando la posibilidad de precisar la dosificación sin la necesidad de pesar la preparación, con un método muy simple de almacenamiento de la inyección y con la eliminación de una inyección adicional del bloqueador de los receptores de DA, siendo justificado hacer más investigaciones usando Ovopel® (7). Con Ovaprim®, un análogo de GnRH con el antagonista de DA Domperidone, existen trabajos en especies como Esox lucius L., comparados con EHC (36), en los ciprínidos Tor tambroides y T. douronensis (20, 21) y silúridos como Pseudoplatystoma fasciatum (29).
El problema arriba mencionado es mucho más visible en el caso de peces silvestres como la sabaleta Brycon henni, la mayoría de las veces con reproductores capturados del medio natural y sometidos a procesos de amansamiento en estanques y que además comienzan a extinguirse, siendo necesario desarrollar técnicas de propagación controlada de esos peces.
Los primeros trabajos que avalaron la distribución de frecuencia porcentual del diámetro de los ovocitos como criterio para la escogencia de hembras reproductoras fueron realizados por Frenerich-Verani , Godinho y Narahara con Curimbatá Prohilodus scrofa (35). Estos autores observaron que el éxito de la fertilización ocurría solamente en hembras con diámetros superiores a
735.55 µm, con distribución unimodal o simétrica y que las hembras que presentaban ovocitos mas grandes pero con distribución polimodal, no respondían a los tratamientos hormonales (35). Lo anterior fue confirmado por Antoniutti et al (3) en hembras de curimbatá Prochilodus scrofa, cuando presentaron distribución de frecuencia porcentual del diámetro de los ovocitos con moda mayor o igual a 972.5 µm y por Andrade-Talmelli en hembras de piabanha, Brycon insignis, avalando el grado de desarrollo de los ovocitos obtenidos por la técnica de canulación asociado a características externas (35).
En Brycon cephalus (35) asociado a los tratamientos hormonales y con la finalidad de determinar el grado de desarrollo del ovario, muestras de ovocitos fueron retiradas antes de la primera y segunda dosis hormonal y en el momento de la ovulación, y de esta manera, se pudo verificar el efecto del EHC sobre la evolución del ovario, la ovulación y el desove bajo condiciones de confinamiento, en hembras seleccionadas para inducir a la reproducción con base en observaciones externas asociadas a la morfología (microscopía de la luz y electrónica ) y distribución de frecuencia del diámetro de los ovocitos (35), reportados también para B. insignis (2), B. opalinus (16, 17, 28) y B. amazonicus (33). En B. opalinus (16, 17), el tipo de desove de la especie fue estimado por graficación del diámetro de los ovocitos y su frecuencia, como desovante total con ovocitos maduros en tallas de 1346.4 µm, alcanzando su máximo a 2509.2 µm y un lote de reserva que incluyó ovocitos en diámetros de 61.2 µm a 367.2 µm, clasificado como sincrónico en dos grupos (16).
El objeto del presente estudio fue determinar la eficiencia de tres protocolos de tratamiento hormonal, a base de EHC, Ovopel y Ovaprim, en la estimulación a la ovulación, especialmente en los cambios en los diámetros ovocitarios antes y después del tratamiento hormonal en la especie sabaleta Brycon henni, durante la temporada reproductiva de Abril-Julio del 2008.
Materiales y métodos
Colección de reproductores y transporte
Los peces fueron capturados de la quebrada San Miguel en el municipio de San Carlos, utilizando para la captura un chinchorro de nylon de 15 m de longitud y un ojo de malla de una pulgada, buscando capturar solamente animales en la talla mínima y media de acuerdo con la clasificación de (5, 14, 26), ayudado con caretas de buceo para localizar los cardúmenes de peces en los charcos. Posteriormente se hacía un encierro y se sacaban selectivamente por tamaños con una nasa triangular, en tallas de 16 y 24 cm aproximadamente y se depositaban en bolsas plásticas, se les inyectaba oxígeno a saturación y se amarraban con bandas de caucho para su transporte a la estación de piscicultura del municipio de San Carlos (Antioquia). Allí fueron alojados en un estanque en tierra de 200 m2 con un recambio del 20% día, en donde permanecieron por cuatro meses en proceso de amansamiento, alimentados a saciedad con concentrado comercial de 32% de proteína y a voluntad con larvas y alevinos de tilapia roja, producto de la reproducción de 10 hembras y cinco machos no reversados de esta especie.
Chequeo de la maduración ovocitaria
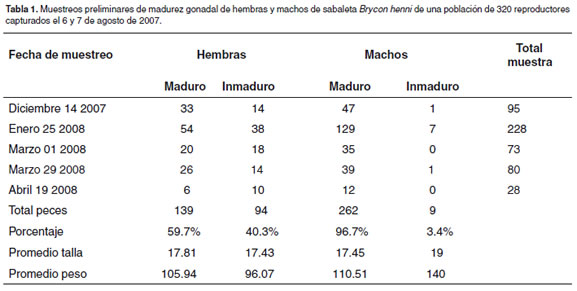
En el mes de diciembre de 2007 y hasta el mes de abril de 2008, se inició la revisión del lote de 320 reproductores hembras y machos y se procedió a revisar el estado de maduración gonadal por sus características externas, tales como vientre abultado y papila ligeramente dilatada y enrojecida en las hembras y expulsión de semen por suave presión abdominal en los machos. Adicionalmente se consideró el criterio de la aleta anal, lisa en las hembras maduras y rugosas o con espinaciones en los machos, propuesto por (11). (véase Tabla 1), se presentan los resultados de los muestreos previos. A partir de abril, se inició el proceso de selección de reproductores para inducción artificial, en las hembras por medio de una cánula nasofaríngea tomando dos muestras de ovocitos y depositándolos, la primera en solución salina para su medición antes del tratamiento hormonal y la segunda en líquido de Serra para clarificación del citoplasma por espacio de cinco minutos. La posición del núcleo de los ovocitos fue determinada usando 3 estadios de desarrollo: Estado 1, vesícula germinal en posición central; estado 2, vesícula germinal en migración (más de la mitad del radio, según (24), y estado 3, ovocitos atrofiados o deformes. Para los experimentos, solamente se tuvieron en cuenta las hembras con más del 50% de núcleos migrados.
Propagación artificial
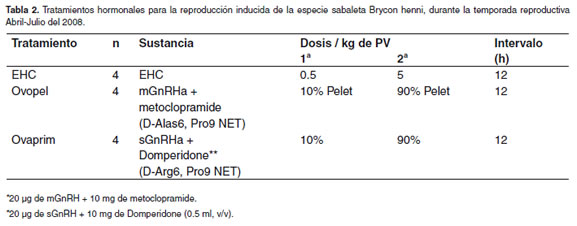
Los peces fueron recapturados del estanque en tierra, en cuatro muestreos periódicos y posteriormente trasladados al laboratorio de la estación piscícola y depositados en contenedores de concreto de 6 m3 con recambio permanente de agua en un sistema de recirculación, aclimatados durante 24 h a una temperatura promedio de 22 ± 1ºC. Mensualmente, se seleccionaron tres hembras y seis machos por muestreo (mayo, junio y julio), con doble muestreo en julio por mayor cantidad de hembras disponibles por características externas, para un total de 12 hembras (cuatro por tratamiento) y 24 machos divididos en tres grupos de tratamiento (T), así: T1 EHC (n=4), T2 Ovopel® (n=4) y T3 Ovaprim® (n=4). Durante los meses de mayo a Julio de 2008, correspondiente con la primera temporada reproductiva, (véaseTabla 2), una hembra y dos machos fueron colocados en cada estanque y fueron tratados con la respectiva inyección hormonal de: extracto hipofisiario de carpa (T1) (EHC-Argent, USA) pulverizados en un mortero, Ovopel (T2), de la misma forma que el EHC y diluidos en solución salina al 0.9% y con 0.5 ml de Ovaprim® (T3), extraído directamente de su empaque con la jeringa. Las inyecciones fueron intramusculares junto a la base de la aleta dorsal. Antes de las manipulaciones, los peces fueron anestesiados con Eugenol en solución de una parte de Eugenol en nueve partes de alcohol absoluto de 96% de pureza y luego disolviendo 0.4 partes de solución de Eugenol en ocho litros de agua, de acuerdo con Arias (comunicación personal). El intervalo de tiempo entre las inyecciones fue de 12 h para los tres tratamientos y los machos recibieron todos una única dosis de 3 mg/kg de peso corporal de EHC (véase Tabla 2), junto con la segunda dosis de las hembras.
Colección de gametos e incubación
Las hembras fueron chequeadas a partir de las 10 h y hasta las 12 -13 h después de la inyección resolutoria o definitiva y justo en el momento en que con una suave presión abdominal, expulsaban una pequeña cantidad de huevos; se procedía a estrujarlas y se depositaban en un recipiente plástico y se fertilizaron usando el método seco con el semen de dos machos. Los siguientes datos fueron tomados: peso de la hembra antes y después del extrujamiento y peso de los huevos extraídos. Adicionalmente se tomaban muestras de 0.1 g de huevos y se fijaban en solución salina para medir el diámetro después del tratamiento hormonal. Los huevos fueron incubados en bandejas tipo californiana, similares a las utilizadas para salmónidos, a la misma temperatura de los contenedores del laboratorio y a las 12 h se registró el porcentaje de huevos fertilizados.
Análisis estadístico
Se empleó el método lineal general (MLG) con el fin de evaluar la variable dependiente “Diámetro de los ovocitos” en función de los tratamientos, fechas de inducción dentro de la temporada reproductiva y muestras o tomas de huevos antes y después del tratamiento hormonal, convalidándose los supuestos adscritos con el diseño de estructura experimental (normalidad, aleatoriedad e independencia de los errores experimentales, homogeneidad de varianzas, aditividad del modelo y no relación entre medias y varianzas). Para contrastar el efecto de los tratamientos y demás fuentes de variabilidad e interés, se empleó el método Tukey con base en un 95% de confiabilidad y, adicionalmente, se efectuaron análisis descriptivos de tipo univariado por tratamiento, por fecha y muestra o toma de huevos antes y después, solo por tratamiento, tratamiento y muestra o toma. Se empleó el paquete estadístico SAS, versión 9.0.
Resultados
Los muestreos preliminares de madurez gonadal realizados entre diciembre de 2007 y abril de 2008, después de cuatro meses de amansamiento en contenedores, muestran que el 59.7% de las hembras y 96.7% de los machos se encontraban maduros y listos para iniciar un proceso de reproducción inducida con hormonas (véase Tabla 1).
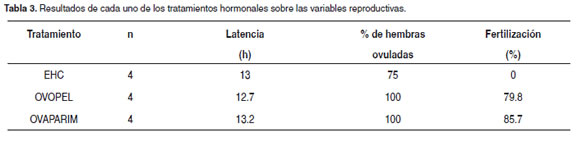
De las 12 inducciones realizadas, 11 hembras respondieron bien a la inducción y ovularon con EHC, Ovopel y Ovaprim (véase Tabla 3). Todas las hembras tratadas con Ovopel y Ovaprim, tuvieron buena respuesta en porcentaje de fertilización, mientras que las cuatro hembras tratadas con EHC no presentaron huevos fertilizados (véase Tabla 3). El periodo de latencia a la ovulación fue muy similar en los tres tratamientos, entre 12.7h y 13.2h (véase Tabla 3). El promedio de los ovocitos en el momento en el que se realizó la biopsia ovárica y justo antes de la administración de la primera dosis hormonal fue de 1428.8, 1531 y 1712 µm con EHC, Ovopel y Ovaprim, respectivamente.
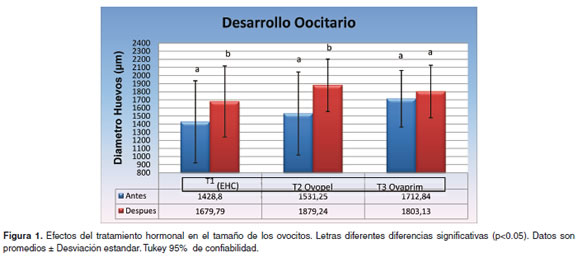
Después de aplicada la segunda dosis hormonal en las hembras y que los huevos fueran extrujados, este diámetro alcanzó 1679.79, 1879.24 y 1803.13 µm, respectivamente para cada tratamiento (véase Figura 1), encontrando diferencias significativas al interior de los tratamientos T1 y T2 (p<0.05), pero no en T3 y un aumento en los diámetros ovocitarios después del tratamiento hormonal de 251 µm, 340 µm y 91 µm respectivamente. En cuanto a la frecuencia de distribución del diámetro de los ovocitos se presentaron en T1 dos modas de 1500 y 2100 µm (véase Figura 2A), antes del tratamiento hormonal (bimodal) y, en el momento de la extrusión, de 900 y 2100, también de tipo bimodal. En el T2 antes del tratamiento hormonal se presentaron dos modas de 900 y 1800 µm (bimodal) y después del tratamiento ovocitos con diámetros de 1800 pasaron a ocupar el 50% del total (véase Figura 2B). En T3 antes del tratamiento se presentaron dos modas de 900 y 1800 µm (bimodal) y fue similar después del tratamiento pero los ovocitos con diámetro de 1800 µm representaron el 55% del total (véase Figura 2C).
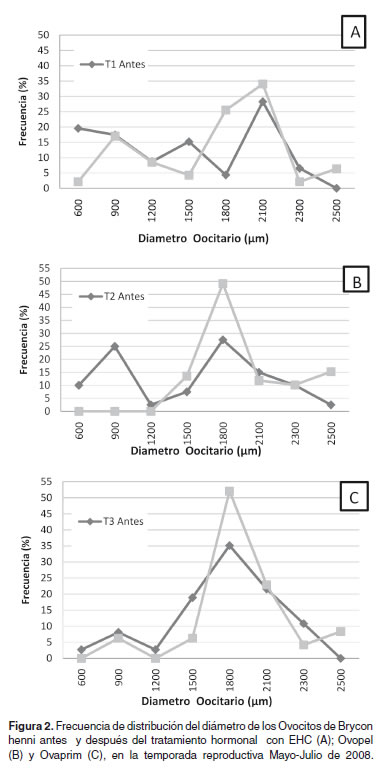
Discusión
El tiempo de latencia entre la aplicación de la segunda dosis y la ovulación de en Brycon cephalus (35), ocurrió entre las 4 y 8 h en los grupos A y B, pero a las 12 h en el grupo C, muy similar al ocurrido en B. opalinus (28), en B. insignis a las 10 h (2) y en este trabajo con B. henni. En B. amazonicus (33) fue mucho más bajo, de 6.7 ± 0.14 h.
Con respecto a la respuesta al tratamiento hormonal y el porcentaje de fertilizacion, con EHC en B.henni, 75% de las hembras respondieron al tratamiento y ovularon, mientras que dosis similares de EHC fueron aplicadas en Brycon cephalus (35) con respuesta positiva en 10 (45.4%) de 22 hembras tratadas con hormona y bajas tasas de fertilización en el grupo C, asociado a varias modas; en Brycon insignis (2) en un tratamiento hormonal similar con EHC, el 27.8% de las hembras seleccionadas por: características externas, verificación del diámetro promedio de mayor frecuencia (1175 µm) y posición del núcleo (60% excéntricos y 40% centrales), respondieron positivamente a los tratamientos hormonales, mientras que el 81.8% lo hicieron con una dosis única de hCG y el 64.7% con una mezcla de ambas hormonas. Después del tratamiento con EHC se presentó una moda con diámetros entre1450 y 1500 (unimodal), asociado con altas tasas de fertilización (2).
En matrinxa B.orbignyanus con dosis de 0.25, 0.5 y 5 de EHC el 66.7% de las hembras respondieron al tratamiento, pero la fertilización fue también muy baja (12.8%) (40), similar a B.henni y en Brycon opalinus (28), con extracto pituitario de salmón, de las 29 hembras tratadas, 14 (48.3%) respondieron positivamente al tratamiento hormonal, con una moda de 1900 y tasas de fertilización de 90%.
En cuanto a la frecuencia de distribución del diámetro promedio de los ovocitos en B. cephalus (35), (grupos A y B) inducidos con EHC presentaron dos modas distintas, antes de la primera dosis hormonal de 939.0 y de 1001.6 µm y después del tratamiento hormonal la frecuencia de ovocitos con diámetros de 1001.6 µm pasaron a representar el 60% del total. En el grupo C la frecuencia porcentual del diámetro de los ovocitos fue de tipo polimodal, asociado a bajas tasas de fertilización (40%). En B. insignis (2), encontraron que el diámetro de mayor frecuencia antes del tratamiento fue de 1250 µm y en el momento de la extrusión de 1450 µm a 1500 µm con tendencia unimodal y con tasas de fertilización superiores al 70%, aumentando 200 a 250 µm el valor del diámetro de los ovocitos de la muestra inicial, similar al T1 en B.henni. Las hembras que no respondieron a los tratamientos, presentaron inicialmente distribución de frecuencia porcentual del diámetro de los ovocitos con modas entre 1200 y 1550 µm y distribución bimodal o polimodal (2).
Con B. opalinus (28), encontraron que las hembras seleccionadas para inducción, presentaban ovocitos con diámetros variando de 1650 a 2150 µm, muy similares a los encontrados en esta investigación con B. henni con modas de 1975 µm para T1 y 1725.5 µm para T2 y T3 después del tratamiento hormonal (véase Figura 2A, 2B, 2C), pero también muy superiores a los encontrados en B. cephalus (35) y, en B. insignis (2). En B. opalinus (16), reportaron un rango alto en talla de maduración ovocitaria de 1346.4 µm, alcanzando un máximo de 2570.4 µm. Sin embargo en B.amazonicus (33) el diámetro ovocitario inicial para todas las hembras, tuvo una distribución unimodal, la cual osciló entre 1000 y 1125 µm y solamente las hembras tratadas con EHC presentaron un crecimiento significativo del diámetro ovocitario.
El análisis de distribución de frecuencias del diámetro de los ovocitos de Salminus hilarii, mostró un lote de ovocitos con picos en una frecuencia de valores entre 1291 ± 24.48 µm y 1353.0 ± 16.5 µm, considerados como vitelogénicos y otro lote con picos entre 202.5 ± 4.5 y 189.8 ± 19.87 µm, claramente revelando que esta especie es un desovante total y que el lote de diámetro pequeño puede ser considerado como de reserva, el cual debería ser desovado en el próximo ciclo reproductivo (19). Cuando la migración es completamente bloqueada, las hembras tienen una baja fecundidad absoluta, pero un número superior de ovocitos maduros, los cuáles son más grandes que en los animales silvestres. La fecundidad es baja en peces en cautiverio, pero el diámetro de los ovocitos es mayor, como se comprobó en Salminus hilarii, donde la fecundidad en cautiverio declinó, pero los animales pudieron compensar esa pérdida produciendo huevos grandes (19).
En un desovador anual, es observada una moda en la talla de los ovocitos (estado III), mientras que en desovadores múltiples, ovocitos vitelogénicos (estado II) de diferentes tallas pueden ser observados en o dentro de un gran número de ovocitos en estado III, según la clasificación de Nuñez y Duponchelle (30). Este parece ser el caso de B.henni en el que aparecen modas de ovocitos en diferentes tallas en el período reproductivo evaluado.
Con referencia a los lotes de ovocitos de mayor talla encontrados en B.henni, es conocido que las hembras en cautiverio producen huevos grandes y liberan pocas cantidades de huevos en cada desove (fecundidad absoluta), cuando son comparados con animales que están en la capacidad de migrar (19). Sin embargo, los estados de maduración alcanzados son adecuados para inducción con manipulación hormonal (19).
Se confirma entonces que la distinción del grado de madurez del animal, debe hacerse por un método que de un alto grado de exactitud como es el diámetro de los ovocitos y que según lo encontrado por Beltrán et al (5) en B.henni, dio como resultado un promedio de 1.5 mm para el estadio III, clasificado como maduro. Isaza (22), con la especie Brycon fowleri reporto ovocitos de 1.5 mm de diámetro.
Álvarez y Duque (1) con B.henni, reportan que antes de la fecundación, el tamaño oscila entre 1.5 y 2 mm y que una vez fecundados y avanzado el desarrollo embrionario, el diámetro aumenta hasta 3 mm. Zapata y Vanegas (41), encontraron que en una hembra de B.henni con un 47% de núcleos excéntricos, se presentó un diámetro promedio de 1.2 mm. Martínez y Vásquez (26), estimaron el diámetro de los ovocitos de B.henni en rangos de tamaño de 0.96 -1.38 mm para el estadio III y de 1.38- 1.8 mm para el estadio IV y en su etapa tardía con tamaños entre 1.80-1.95 mm de diámetro.
Lo anterior sugiere, por lo encontrado en este trabajo, que B.henni podría tener su mejor rango de diámetro ovocitario entre 1700 y 1800 µm y demuestra la importancia de la utilización de este parámetro como criterio de selección de hembras para inducir su reproducción y muy similar a lo reportado con B. opalinus (28), pero debido a la amplitud de la variación observada en los valores del diámetro medio de los ovocitos, no se puede utilizar únicamente este criterio, pero si un valor modal de distribución de frecuencia porcentual.
Con relación al período reproductivo evaluado (abril-Julio), en los embalses del El Peñol y Troneras en Antioquia (5) se presentaron ejemplares sexualmente maduros, correspondiendo el mayor porcentaje en mayo, coincidiendo con Perdomo (34) en que para esta especie hay dos épocas de desove en el año, con picos máximos en abril y mayo (primera época) y en octubre-noviembre (segunda época).
En sabaleta B.henni, según Florez-Brand (14) en el embalse de Salvajina, se distinguen dos períodos en los cuáles la mayoría de los ejemplares se encuentran sexualmente maduros, la primera época entre marzo y junio y la segunda entre octubre y enero, que coinciden con los períodos lluviosos o de aguas altas, incrementándose en los períodos de marzo, abril y mayo y octubre, noviembre, diciembre. Para el mismo sitio, Martínez y Vásquez (26) detectaron con base en el índice Gonadosomático (IGS) los meses de abril, julio y agosto como épocas de desove y que tanto hembras como machos presentaron estadio IV (maduros), casi todo el año, con máximos en abril (80%), julio (71.4%) y diciembre (75%).
En esta investigación se encontraron ejemplares maduros durante toda la temporada evaluada de abril a julio, coincidiendo con los autores citados. En B. amazonicus (4) el inicio de la estación de lluvias fue el estímulo determinante para la finalización de la maduración y desove en el mes de abril en el que se registró igualmente el mayor valor del IGS. Otros trabajos con Salminus hilarii (19), sugieren que para esta especie y con base en el IGS y la frecuencia de distribución del diámetro de los ovocitos, que el período reproductivo de tabarana ocurrió de septiembre a febrero.
En cuanto a los tratamientos hormonales con Ovopel y Ovaprim (T2 y T3), en Perca fluviatilis (23), utilizando una mezcla de FSH y LH con Pimozide (P) o Metoclopramide (M), encontraron que este último fue más potente en hembras. Todos los machos estuvieron espermiando al tiempo que ellos fueron capturados y por lo general, los tratamientos hormonales no ocasionaron un significativo cambio en la cantidad de esperma obtenida de los machos. En B.henni se encontró un comportamiento completamente igual y en la tabla 1 puede verse que permanecieron maduros durante todos los muestreos más del 96% de los machos. En Golfish Carassius auratus, M fue menos potente que P (38). En Perca la ovulación de la hembra fue muy sincrónica, la mayoría ovularon entre 16 y 22 h después de la inyección resolutoria y la latencia en los peces que se les administro FSH y LH con 10 mg de M fue más corta (16 - 18 h). Es posible que la solubilidad del M y por tanto su rápido absorción en el sitio de la inyección, sean una explicación para la más rápida respuesta al desove.
Con B. henni las ovulaciones ocurrieron entre 12 y 13 horas para los tres tratamientos EHC, Ovopel que incluye M y Ovaprim, con el antagonista de DA, Domperidone (D). Yaron (28), sugiere que la latencia fue siempre más corta en peces tratados con EHC que con otras hormonas; esto puede ser explicado por el hecho que la liberación de GnRH y su respuesta en el ovario es un proceso secuencial, mientras que en los peces inyectados con EHC, la respuesta del ovario a la gonadotropina (GtH) exógena es un proceso simple (23). En el ciprínido Leuciscus idus (24), el mayor problema fue la sobrevivencia de la ovas, especialmente de los peces tratados con Ovopel. Esta combinación hormonal (mGnRHa y M) provee muy buenos resultados pero el problema de mortalidad mostró que son necesarios más estudios.
En B.henni se encontró que el tratamiento con Ovopel tiene altos porcentajes de fertilización (véase Tabla 3), pero después de 12 h de incubación hay mortalidad total de las ovas. Kucharczyk (24), encuentra una buena calidad de los gametos después de la estimulación hormonal y sugiere que debería estar correlacionada con la temperatura del agua, tanto en el desove como en los huevos incubados. En B.henni este procedimiento se efectuó a temperaturas de 22 ± 1ºC, muy similar a lo reportado con B. opalinus (28), de 21 ± 1ºC.
Más investigaciones son requeridas para determinar el mejor momento de la estimulación hormonal para B.henni, considerando otros criterios adicionales al diámetro ovocitario, la segunda temporada reproductiva, el factor de condición y la temperatura de incubación entre otros.
Agradecimientos
A la Corporación para el Desarrollo Sostenible de las Cuencas Rionegro-Nare, CORNARE e ISAGEN, por la financiación de las investigaciones que se llevan a cabo con esta especie en la región del Oriente Medio Antioqueño. A Ligia Ramírez, Rodrigo Cardona y al Alcalde del municipio de San Carlos Juan Alberto García, por el apoyo logístico en la estación de piscicultura; al Ingeniero Acuícola Alfredo Gómez Zarate y al estudiante de Zootecnia Danny Andrés Isaza por el acompañamiento en el trabajo de campo, quienes con la Universidad de Antioquia, a través del Centro de Investigaciones Agrarias y el CODI, hicieron posible la realización de esta investigación.
Referencias
1. Álvarez LF y Duque JP. La sabaleta y su cultivo en estanques. Seminario (Zootecnia); Medellín:Facultad de Ciencias Agropecuarias,Universidad Nacional de Colombia;1989. P.1-34 . [ Links ]
2. Andrade Talmelli EFA, Kavamoto EM, Narahara MY, Fenerich-Verani, Reprodução induzida da piabanha, Brycon insignis, mantida em cativeiro. Rev Brás Zootec 2002;31:803-811. [ Links ]
3. Antoniuti DM Naraha MY, Romagosa E. Reproducao induzida ecusto operacional de producao de alevinosde curimbata, Prochilodus scrofa (Steindachener, 1881). B Inst Pesca Sao Paulo 1995;22:41-47. [ Links ]
4. Arias CJA, Zaniboni E, Aya BE Indicadores Del ciclo reproductivo Del Yamú Bricon amazonicus, em cautiverio. Revista Orinoquia 2006;10:24-34 [ Links ]
5. Beltrán CN, Beltrán I, Sierra R. Notas preliminares sobre la reproducción y cultivo de la sabaleta (Brycon henni, Eigenman 1913). Inderena, 1978:27p. [ Links ]
6. Berlinsky DL, Smith T IJ, McVey DC, Jenkins WE, Denson MR, et al. Broodstock management and spawning of southern fl ounder, Paralichthys lethostigma Aquaculture 1998;176: 87-99. [ Links ]
7. Brzuska E. Artifcial spawning of European catfsh Silurus glanis L.: differences between propagation results after stimulation of ovulation with carp pituitary and Ovopel. Aquac Res 2001;32:11-19. [ Links ]
8. Brzusca E, Bialowas H. Artifi cial spawning of carp, Cyprinus carpio (L.) Aquac Res 2002; 33:753-765. [ Links ]
9. Brzuska E. Artifi cial spawning of female Polish line 3 carp (Cyprinus carpio L.) after treatment with pituitary homogenate and/or Ovopel. Aquaculture Research. 2003;34: 1321-1327. [ Links ]
10. Brzuska E. Artifi cial spawning of female Lithuanian strain B carp (Cyprinus carpio L.) after treatment with carp pituitary homogenate, Ovopel or [d-Tle6, ProNHEt9] GnRH-a (Lecirelin). Aquac Res 2006:37: 264-271. [ Links ]
11. Builes J, Uran A. Estudio del ciclo sexual de la sabaleta (Brycon henni 1913) su comportamiento y fecundación artificial. Rev Actual Biol 1974; 3: 2-12. [ Links ]
12. Builes J, Lara L. Informe sobre las actividades realizadas durante el año 1980 en el programa de piscicultura agrícola de la sabaleta (Brycon henni). Corpourabá, Medellín, 1980; 310-324. [ Links ]
13. Das SK. Evaluation of a New Spawning Agent, Ovopel inInduced Breeding of Indian Carps. Asian Fish Sci 2004;17: 313-322. [ Links ]
14. Flórez Brand, P.E. Estudio biológico-pesquero preliminar de tres especies icticas del alto del Río Cauca Embalse de Salvajina. Rev Cespedesia 1999; 23:47-61. [ Links ]
15. Forniés MA, Mañanós E, Carrillo M, Rocha A, Laureau S, et al. Spawning induction of individual European sea bass femalesž Dicentrarchus labrax/using different GnRHadelivery systems. Aquaculture 2001;202: 221-234. [ Links ]
16. Gomiero LM, Braga FMS. Gonadossomatic relation and reproductive strategy of Brycon opalinus (Cuvier, 1819) in the Serra do Mar State Park -Núcleo Santa Virgínia, Atlantic Forest, Brazil. Braz J Biol 2007;67: 727-733. [ Links ]
17. Gomiero LM, Braga FMS. Reproduction of Pirapitinga do Sul (Brycon opalinus Cuvier, 1819) in the Parque Estadual da Serra do Mar-Núcleo Santa Virgínia, São Paulo, Brazil. Braz J Biol 2007; 67: 541-549. [ Links ]
18. Haddy JA, Pankhurst NW. The effi cacy of exogenous hormones in stimulating changes in plasma steroids and ovulation in wild black bream Acanthopagrus butcheri is improved by treatment at capture. Aquaculture 2000;191: 351-366. [ Links ]
19. Honji RM, Narcizo AM, Boella MI. Patterns of oocyte development in natural habitad and captive Salminus hilarii Valenciennes. Fish Physiol Biochem 2009;35:109- 123. [ Links ]
20. Ingram B, Sungan S, Gooley G, Sim SY, Tinggi D, et al. Induced spawning, larval development and rearing of two indigenous Malaysian mahseer, Tor tambroides and T. douronensis, Aquac Res 2005; 36:1001-1014. [ Links ]
21. Ingram B, Sungan S, Tinggi D, Sim SY, De Silva S. Breeding performance of Malaysian mahseer, Tor tambroides and T. douronensis broodfish in captivity, Aquac Res 2007; 38:809-818. [ Links ]
22. Isaza BM. Ensayo de cultivo de sabaleta Brycon fowleri en Urabá. Trabajo de investigación. Departamento de Biología, Universidad de Antioquia, Medellín, 1983. 83p. [ Links ]
23. Kucharczyk D, Kujawa R, Mamcarz A, Wyszomirska E, Ulikowsky D. Induced spawning in perch, Perca fluviatilis L. using FSH and LH with pimozide or metoclopramide. Aquac Res 1998;29:131-136. [ Links ]
24. Kucharczyk D, Kujawa R, Mamcarz A, Wyszomirska E, Ulikowsky D. Artifi cial spawning of ide (Leuciscus idus) under controlled conditions. EJPAU. Electronic Journal of Polish Agricultural Universities. 1999; (noviembre de 2007) URL: http://www.ejpau.media.pl/volume2/issue2/fisheries/art-05.html. [ Links ]
25. Kucharczyk D, Szczerbowski A, Luczynski MJ, Kujawa R, Mamcarz A, et. al. Artifi cial spawning of Eurasian perch, Perca fluviatilis L. using Ovopel. Arch Pol Fish 2001; 9: 39-49. [ Links ]
26. Martínez OH, Vásquez GZ. Aspectos reproductivos de la sabaleta Brycon henni (Pisces Characidae), en el embalse La Salvajina, Colombia. Rev Asoc Colomb Ictiol 2001;4:75-82. [ Links ]
27. Mikolajczyk T, Roelants I, Epler P, Ollevier F, Chyb J, et al. Modifi ed absorption of sGNRH-a following rectal and oral delivery to common carp, Cyprinus carpio L. Aquaculture 2002; 203: 375-388. [ Links ]
28. Narahara, MY, Talmeli A, Kavamoto EFE, Godinho HM. Reprodução induzida da pirapitinga do sul, Brycon opalinus, mantida em condições de confi namento. Rev Bras Zootec 2002; 31:1070-1075. [ Links ]
29. Nuñez J, Dugue R, Corcuy NA, Duponchelle F, Renno J, et al. Induced breeding and larval rearing of Surubí, Pseudoplatystoma fasciatum (Linnaeus, 1766), from the Bolivian Amazon. Aquac Res 2008; 39:764-776. [ Links ]
30. Nuñez J, Duponchelle F. Towards a universal scale to assess sexualmaturation and relatedlife history traits in oviparous teleost fishes. Fish Physiol Biochem 2009;35:167-180. [ Links ]
31. Montoya A, Carrillo LM, Olivera-Angel M. Algunos aspectos biológicos y del manejo en cautiverio de la sabaleta Brycon henni Eigenmann, 1913 (Pisces Characidae). Rev Colomb Cienc Pecua 2006;19: 180-185. [ Links ]
32. Pankhurst N, Thomas P. Maintenance at elevated temperature delays the steroidogenic and ovulatory responsiveness of rainbow trout Oncorhynchus mykiss to luteinizing hormone releasing hormone analogue. Aquaculture 1998;166:163-177. [ Links ]
33. Pardo S, Arias JA, Suárez-Mahecha H, Cruz-Casallas P, Vasquez-Torres W, et al. Inducción a la maduración fi nal y ovulación del Yamú Brycon henni con EPC y mGnRHa. Rev Colomb Cienc Pecua 2006; 19:160-166. [ Links ]
34. Perdomo JM. La sabaleta (Brycon henni, Eigenman 1913). Observaciones bioecológicas y su importancia como especie de cultivo. Rev div pesq 1978; 11:1-46. [ Links ]
35. Romagosa E, Narahara MY, Borella MI, Fenerich VN. Seleção e caracterização de fêmeas de matrinxã, Brycon cephalus induzidas a reprodução. Bo Inst Pesca 2001; 27:139-147. [ Links ]
36. Szabo´ T, Medgyasszay C, Horváth L. Ovulation induction in nase (Chondrostoma nasus,Cyprinidae) using pituitary extract or GnRH analogue combined with domperidone. Aquaculture. 2002;203: 389-395. [ Links ]
37. Ulikowski D. European Catfish (Silurus glanis L.). Reproduction outside of the spawning season. Arch Pol Fish 2004;12: 121-131. [ Links ]
38. Yaron Z. Endocrine control of gametogenesis and spawing induction in the carp. Aquaculture. 1995;129: 49-73. [ Links ]
39. Zakes Z , Demska K . Artifi cial Spawning of Pikeperch (Sander lucioperca L.), Estimulated with Human Chorionic Gonadotropin (HCG) and Mammalian GnRH Analogue with a Dopamine Inhibitor. Arch Pol Fish 2005; 13:1:63- 75. [ Links ]
40. Zanoboni Filho E, De Campos Barbosa ND. Priming hormone administration to induce spawning of some brazilian migratory fish. REVISTA 1996;vol:63-67. [ Links ]
41. Zapata L, Vanegas R. Aspectos importantes sobre la reproducción inducida y el metabolismo de las gónadas de sabaleta Brycon henni. Trabajo de grado, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, 1993. 67p. [ Links ]
42. Zohar Y, Mylonas C. Endocrine manipulations of spawning in cultured fish: from hormones to genes. Aquaculture 2001; 197: 99-136. [ Links ]

