










Serviços Personalizados
Journal
Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
versão impressa ISSN 0120-0690versão On-line ISSN 2256-2958
Rev Colom Cienc Pecua v.22 n.2 Medellín abr.//jun. 2009
Reproducción inducida y producción de alevinos de Sabaleta Brycon henni: determinación del tiempo de latencia utilizando extracto de hipófisis de carpa *
Induced breeding and production of fingerlings of Sabaleta Brycon henni: Determining the time of latency using Carp Pituitary Extract
Reprodução induzida e produção de alevinos da Sabaleta Brycon henni: determinação do tempo de latência utilizando Extrato de Hipófise de Carpa
Gustavo A Lenis1*, Biólogo, cMSc; Luis F Restrepo1, Estadístico Esp; Juan C Rivera J, Ing Acuícola; Fabián Monsalve, Ing Acuícola; Pablo E Cruz-Casallas2, MVZ, MSc, PhD.
1 Grupo de Investigación GRICA. Facultad de Ciencias Agrarias, Universidad de Antioquia, AA 1226, Medellín, Colombia. 2 Grupo de Reproducción y Toxicología GRITOX, Instituto de Acuicultura, Facultad de Ciencias Agropecuarias y Recursos Naturales, Universidad de los Llanos, A.A 110, Villavicencio, Meta - Colombia.
(Recibido: 29 octubre, 2008; aceptado: 26 abril, 2009)
Resumen
Esta investigación fue realizada en la estación piscícola de San José del Nus (Antioquia). Fue iniciada en el año 2002 con la captura y adaptación de un lote de 100 reproductores silvestres, provenientes de tres microcuencas cercanas a la estación piscícola y fue continuada en el año 2003 con la captura de otros 80 ejemplares, con los cuales se hicieron los primeros ensayos de inducción hormonal estimulando la maduración final y la ovulación con dos dosis de Extracto Pituitario de Carpa (EPC) (0.5 + 5.0 mg/kg de peso corporal). Para la obtención de los gametos se utilizó extrusión en seco en los tres grupos de peces. La maduración final y la ovulación se indujo en 20 hembras y la espermiación en 20 machos. El resultado más relevante fue la ovulación y desove de ocho hembras, de las cuales sólo dos fueron completamente exitosas, con un tiempo de latencia de 13 y 11 h, respectivamente. De la primera hembra con un peso de 136.2 g, se extrajeron 1421 huevos, los cuales fueron fertilizados, mostrando una sobrevivencia de embrión a larva de 28% y de larva a alevino de 35.7%. La segunda hembra, de 90 g de peso corporal, desovó 1348 huevos mostrando un porcentaje de fertilidad de 74%, sobrevivencia larval del 40% y sobrevivencia hasta alevino comercial de 25%. En cada uno de los desoves se obtuvo un número de alevinos netos de 141 y 101, respectivamente. La temperatura de incubación fue de 24.5 ºC y de 25.3 ºC. Cabe destacar que el en mismo año se encontró una hembra ya desovada que presentó nuevamente madurez gonadal tres meses más tarde.
Palabras clave: desove, eclosión, fertilización, inducción hormonal, latencia, viabilidad.
Summary
This research was conducted in the San José del Nus fish farm, located in the Municipality of San Roque (department of Antioquia). This study began in 2002 with the capture and suitability of 100 wild breeding fish batches coming from three sub-basins near to San José del Nus fish farm. In 2003, another 80 specimens were captured and utilized for first hormonal induction trials by stimulating final maturation and ovulation with two doses of Carp pituitary extract (CPE) (0.5 + 5.0 mg/kg B.W.). Dry extrusion was used in the three groups of fish to obtain gametes. Spermiation and final maturation plus ovulation were induced in twenty males and females, respectively. The most relevant result was ovulation and spawning in 8 cases. Two of them were completely successful; their latency period was 13 and 11 h, respectively. From the first previously fertilized female (136.2 g. B.W.) 1421 eggs were extracted, with a survival of 28% from embryo to larva; and 35.7% from larva to alevin. The second female (90 g B.W.) spawned 1348 eggs, resulting in a fertilization rate of 74%, larval survival of 40% and survival until alevin for commercial use of 25%. In both cases, spawning resulted in an alevin net number of 141 and 101, respectively. Water incubation temperature was 24.5 ºC and 25.3 ºC. It is important to emphasize that in the same year a previously spawned female was found, and three months later this female presented gonadal maturity once again.
Key words: fertilization, hatching, hormonal induction, latency, spawning, viability.
Resumo
Esta pesquisa foi realizada na estação de piscicultura de São José del Nus (Antioquia). Foi iniciada no ano 2002 com a captura e adaptação de um lote de 100 reprodutores selvagens provenientes de três córregos perto da estação piscicola e foi continuada no ano 2003 com a captura de outros 80 exemplares, com os quais foram feitos os primeiros ensaios de indução hormonal estimulando a maduração final e a ovulação com duas doses de extrato de hipófise de carpa (0.5 + 5.0 mg / kg de peso corporal). Para a obtenção dos gametas se utilizo extrusão em seco para os três grupos de peixes. A maduração final e a ovulação foi induzida em 20 fêmeas e a espermiação em 20 machos. O resultado mais relevante foi a ovulação e desova de oito fêmeas, das quais apenas duas foram completamente bem sucedidas, o período de latência foi de 13 e 11 h, respectivamente.. Da primeira fêmea, com 136.2 g de peso corporal, obtiveram-se 1421, os quais foram fertilizados, com uma sobrevivência desde embrião a larva de 28% e de larva até alevin de 35.7%. A segunda fêmea de 90 g de peso corporal, desovou 1348 ovos mostrando uma percentagem de fertilidade de 74%, sobrevivência larval de 40% e sobrevivência desde larva a alevin comercial de 25%. Em cada uma das desovas foram obtidos 141 e 101 alevines, respectivamente. A temperatura da água de incubação foi de 24.5 ºC e 25.3 ºC, respectivamente. Outra observação importante foi que, durante o mesmo período encontrouse uma fêmea desovada que apresentou de novo maturação gonadal 3 meses mais tarde.
Palavras chave: desova, eclosão, fertilização, indução hormonal, latência, viabilidade.
* Para citar este artículo: Lenis GA, Restrepo LF, Rivera JC, Monsalve F, Cruz-Casallas PE. Reproducción inducida y producción de alevinos de sabaleta Brycon henni: determinación del tiempo de latencia utilizando extracto de hipófisis de carpa. Rev Colomb Cienc Pecu 2009; 22:143-155
* Autor para correspondencia: Facultad de Ciencias Agrarias, Carrera 75 No 65 -87, Ciudadela Robledo, Universidad de Antioquia, Medellín, Colombia. Email: g-lenis@hotmail.com.
Introducción
La reproducción inducida en peces consiste en administrar a los reproductores, hormonas de origen natural o sintético para provocar la maduración final de sus gónadas y la ovulación o espermiación, cuando bajo las condiciones de cautiverio no experimentan los estímulos ambientales necesarios para desencadenar las respuestas endocrinas que gobiernan el proceso reproductivo. Estos estímulos inciden sobre el hipotálamo provocando la liberación de la hormona liberadora de gonadotropinas (GnRH) (46), la cual estimula las células gonadotropas de la hipófisis, induciendo la producción de las hormona FSH, encargada de la formación de vitelo y de la hormona LH, responsable de la maduración final de los ovocitos (67). En este proceso también participan otras hormonas de origen gonadal, tales como testosterona, 17β estradiol (65), 17α-20β dihidroxi-progesterona (29), 17α, 20β-dihydroxi-4-pregnen-3-ona (17α, 20βP) y 17-20β-21 trihidroxi-4 pregnen-3 ona (1, 21, 28).
El uso de diferentes sustancias para inducir la vitelogénesis, maduración, ovulación y, finalmente el desove en las diferentes especies de peces en cautiverio ha sido evaluado por varios autores (8, 18, 19) y actualmente vienen empleándose con éxito en varias especies de peces marinos y estuarinos, tales como los lenguados Paralichthys tropicus con gonadotropina coriónica humana (hCG) (56) y el mero Epinephelus marginatus (38), entre muchas otras. Se han obtenido también resultados exitosos en especies de peces dulceacuícolas como Brycon cephalus (55) y Brycon insignis (3), B.amazonicus (34), utilizando en las dos especies Extracto de Hipófisis de Carpa (EHC) (66), y en Brycon opalinus (48) usando Extracto de Hipófisis de Salmón (EHS). En siluriformes, como Pseudoplatystoma fasciatum (35) también se ha reportado el uso exitoso de EHC, solo o combinado con hCG.
En carpas Cyprinus carpio, Hypophthalmichthys molitrix, Ctenopharyngodon idella y en el Bleak o mento Chalcalburnus calcoides (33), se ha reportado el uso de GnRHa sola o combinada con EHC (12). En Colombia hay reportes de resultados satisfactorios con hCG en bocachico Prochilodus magdalenae (57), EHC en cachama blanca y negra Piaractus brachypomus y Colossoma macropomum (53, 63, 66), blanquillo Sorubim cuspicaudus (8), yaque Leiarius marmoratus (15), curimbatá Prochilodus scrofa (42), yamú Brycon amazonicus (5, 6, 7), barbilla Rhamdia sebae (22) entre otras especies tropicales. Con Ovaprim, un producto comercial que incluye GnRHa combinado con un bloqueador de los receptores de dopamina (Domperidone) se tienen reportes de su uso exitoso en Sorubim cuspicaudus (9), barbilla Rhamdia sebae Cf. (22), moneda Methinnis spp (17) y tigrito Pimelodus pictus (Aya E., comunicación personal). Con Ovopel, una combinación de GnRHa y el antidopaminérgico Metoclopramide, se tienen reportes en las especies barbilla Rhamdia sebae Cf. y yamú Brycon amazonicus (5).
Las variables para medir la respuesta a la administración de las hormonas son especialmente la progresión de la maduración del ovocito, la ovulación y el desove, acompañado o no de cortejo entre los reproductores, así como la medición de varios parámetros de desempeño reproductivo como el tiempo de latencia, el número y el tamaño de los ovocitos pre y post desove, el porcentaje de fertilización, la sobrevivencia embrionaria y larval y finalmente el número de alevinos obtenido.
La sabaleta Brycon henni, una especie nativa de las quebradas de la zona cafetera colombiana, parece ser un importante bioindicador con relación a la calidad de las aguas corrientes y el decrecimiento de esta especie reofílica, al parecer, es producto de la fragmentación de las corrientes, por la construcción y operación de represas. Sin embargo, una de las dificultades para trabajar con esta especie, con potencial para la acuicultura nacional, es obtener cantidades comerciales de huevos viables y predecir con cierto grado de exactitud el tiempo de latencia, después de que los huevos por tratamiento hormonal, son ovulados y expulsados hacia la cavidad ovárica del pez, para proceder con el extrujamiento y la seminación artificial. Algunos problemas todavía persisten como la incertidumbre de las hembras para que respondan positivamente al tratamiento hormonal y parcialmente a la ovulación, lo cual hace prácticamente imposible la obtención de los huevos por una simple extrusión (68). En Brycon henni, se han realizado estudios de biología (36, 50, 52), ecología (20, 49, 51) y pesquerías, reportados por varios autores (20, 36, 50). En reproducción inducida y desarrollo gonadal, además de los trabajos preliminares a partir de protehormonas (54), de Builes y Uran (13) Zapata y Vanegas (71) y de Perdomo (50), se reportan algunos trabajos sobre desempeño reproductivo de la especie (10, 26, 41) y una revisión bibliográfica sobre el tema (43).
En cuanto a cultivo, únicamente se han reportado los trabajos de Álvarez y Duque (2), Arboleda et a. (4), Greco y Granda (27) y de Isaza con Brycon fowleri (30). En aspectos relacionados con la caracterización seminal y métodos de crioconservación y descripción de la gónada masculina, también se han reportado recientemente varios trabajos (60, 61, 62) y un trabajo comparado sobre conservación de semen para tres especies del género Brycon, incluyendo a B. henni (11).
Aunque la biología reproductiva es el tópico más estudiado sobre la especie, sin embargo se necesitan investigaciones que esclarezcan preguntas relacionadas con parámetros medioambientales que desencadenan diversas respuestas reproductivas, protocolos efectivos de inducción hormonal con diversas sustancias inductoras y descripción del desarrollo embrionario y larval (45). Realizando una descripción anatómica e histológica de las gónadas, haciendo énfasis en aspectos de su biología reproductiva como la gametogénesis y su relación con algunas variables ambientales a lo largo del año, según los resultados encontrados en este trabajo, B. henni desovaría en los meses de menor pluviosidad (44).
El desarrollo de un protocolo de reproducción de sabaleta es importante para la conservación de la especie, la cual se encuentra amenazada de extinción debido a la contaminación, destrucción de su hábitat, bloqueo en su proceso migratorio ocasionado por los embalses y a la sobrepesca a que se encuentra sometida la especie (37). Aparte de esto, la estandarización de un protocolo de reproducción inducida de sabaleta aportaría a los programas de repoblación de cuencas y microcuencas existentes y podría abrir las puertas a un nuevo mercado de este pez, que tradicionalmente ha sido uno de los más apreciados en Antioquia tanto por su valor deportivo como gastronómico.
Materiales y métodos
El trabajo se realizó en la estación piscícola de San José del Nus, de la Universidad de Antioquia, localizada en el municipio de San Roque Departamento de Antioquia, Colombia, a una latitud de 6º 2’ N, longitud de 74º 50’ 0, altura promedio de 830 msnm, precipitación promedio anual de 2200 mm, ubicada en una zona de bosque muy húmedo premontano (bmh - PM). Se utilizaron tres grupos de reproductores capturados entre septiembre de 2002 y abril de 2003 en las microcuencas La Clara (municipio de Gómez Plata) y las quebradas Quivocá y Ahumareda, localizadas en el Municipio de Puerto Berrío, en el corregimiento de Virginias. Se hicieron muestreos preliminares en los tres sitios con el objeto de comprobar la presencia de peces de la especie sabaleta, utilizando métodos artesanales de captura, especialmente una atarraya de ojo de malla de una pulgada y 2.5 m de diámetro, procurando capturar únicamente animales en edad reproductiva. Para las labores de reproducción se utilizaron las instalaciones del Laboratorio de la Estación Piscícola, con dos piletas circulares de 2.5 m de diámetro y una altura de 1.1 m y seis piletas para incubación con dimensiones de 6m de largo por 1m de ancho y 1.10 m de alto.
Peces Experimentales
Para la selección y muestreo de los reproductores, se realizó un arrastre con chinchorro de nylon de ojo de malla de 1 cm, sin nudo, de 2 m de altura. Se preseleccionaron hembras por sus características externas como vientre abultado y papila genital ligeramente dilatada y enrojecida; los machos por salida de semen al ejercer suave presión sobre el vientre. Se transportaron dentro de bolsas plásticas con agua hacia las piletas circulares de concreto, aprovisionadas con agua limpia. Se pesaron utilizando balanza Ohaus con capacidad de 500 g y se identificaron en la aleta dorsal. Para la biopsia ovárica se utilizó una cánula nasofaríngea pediátrica Nº 6; las muestras de ovocitos se fijaron en suero fisiológico al 0.5% NaCl para contar el número de ovocitos por gramo, utilizando un estereomicroscopio marca Nikon. Luego se retiró el suero fisiológico y se adicionó solución Serra (85% alcohol etílico al 90% de pureza, 10% formol y 5% ácido acético glacial); Cinco minutos después se procedió a contar los ovocitos con núcleo central, en migración y atrésicos.
Inducción Hormonal
Para las inducciones se prepararon las dosis de extracto de hipófisis de carpa (EHC) para macho (3 mg/kg P.V.) y hembra (0.5 -5 mg/kg P.V.), con ayuda de una balanza analítica marca Ohaus de 0.0001 g. de precisión; la hormona liofilizada fue macerada en un vidrio de reloj adicionándole 0.5 ml de suero fisiológico. Cuando la solución se homogenizó se agregó 0.5 ml más de suero para lavar el vidrio y completar la dosis de 1 ml de dilución por cada aplicación. Las hembras se inyectaron con intervalos de 10 h entre la primera y la segunda dosis. Los machos se inyectaron simultáneamente con la segunda dosis de las hembras. Posteriormente se inició la medición de temperatura con un termómetro de máximas y mínimas, cada hora, con el objeto de determinar el “tiempo de latencia” al desove. También se registró la temperatura y la concentración de oxígeno disuelto en el agua de la pileta con un oxímetro digital marca WTW, desde el momento de la captura de los reproductores y durante todo el proceso de aplicación de hormona, para relacionarlos con el comportamiento de los reproductores.
Obtención de los Gametos
Seis horas después de la última dosis, se inició la observación del comportamiento reproductivo de los peces como acompañamiento y cortejo, de acuerdo con experiencias de otros Brycon (B. moorei y B. amazonicus). Entre las 8 y las 13 h pos inducción, las hembras fueron sacadas cada hora de la pileta, se hacía una suave presión abdominal hasta el momento en el cual se obtenían huevos y se procedía a sedarlos con 2-fenoxietanol (6 gotas en 10 L de agua) por tres minutos y se estrujaron, recogiendo los huevos en un plato plástico previamente tarado para pesarlos. Se contó en un gramo el número de unidades y los huevos obtenidos fueron repartidos en dos platos. El semen se colectó directamente por estrujamiento del macho, seminando los dos grupos de huevos, agregando agua fresca para activar la movilidad de los espermatozoides y mezclando cada grupo con una pluma por dos minutos. Se realizaron siete recambios de agua para retirar impurezas.
Evaluación del desempeño reproductivo
Los huevos seminados se depositaron en una incubadora tipo californiana o de bandeja. A las seis horas postseminación se determinó el porcentaje de fertilización, para lo cual se tomaron tres muestras y se observaron cuales estaban fertilizados. A las 30 h posfertilización, se calculó el porcentaje de sobrevivencia embrionaria, contando los huevos con larvas a punto de eclosionar y en proceso de desarrollo en la incubadora. Después de la eclosión que ocurrió entre las 40 y 60 h postfertiización, se procedió a recolectar las larvas, retirando la cubierta de la incubadora californiana con los restos de huevos y capturándolas con un recipiente plástico tipo beaker de 1000 ml;, se contaron, se sembraron en una pecera de vidrio y se le suministró nauplios de Artemia salina durante una semana, luego se trasladaron a un estanque en tierra de 40 m2, similar al que se utiliza para la adaptación de reproductores, previamente abonado con un bulto de 50 kg de bovinaza bien disuelta y filtrada. También utilizando plancton, colectado con malla de 300 µm de otros estanques abonados.
A los quince días de sembradas las larvas, se inició la alimentación exógena en las horas de la mañana (entre 5:00 y 6:00 am) y en las horas de la tarde (entre 17:00 y 18:00 pm) con alimento balanceado molido de 45% de proteína. Treinta días después se cosecharon los alevinos utilizando un chinchorro de 5 m de longitud, 1.20 m de altura y ojo de malla de 1/4 de pulgada y se cambiaron de estanque para dar mejores condiciones en cuanto a suministro de agua y alimentación. Cada hora se registró la temperatura y la concentración de oxígeno disuelto.
Análisis Estadístico
Se empleó el modelo lineal general (GLM) con el objetivo de probar si existía diferencia estadística en el efecto de la Latencia, entre los tres grupos de peces inducidos hormonalmente, durante las temporadas febrero-marzo, octubre-noviembre de 2003 y febrero de 2004, convalidándose los supuestos de normalidad, aleatoriedad e independencia de los errores experimentales y, adicionalmente, la homogeneidad de varianzas entre grupos de peces inducidos. Con el objetivo de comparar el efecto promedio, se empleó la prueba de Tukey, con un nivel de confiabilidad de 95%. De igual forma se llevó a cabo análisis descriptivo exploratorio de tipo unidimensional, con el objetivo de establecer la media aritmética, la desviación típica y el coeficiente de variación. Para todos los análisis se empleó el software de estadística SAS, versión 8.2.
Resultados
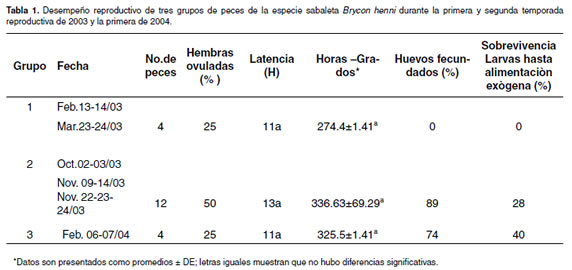
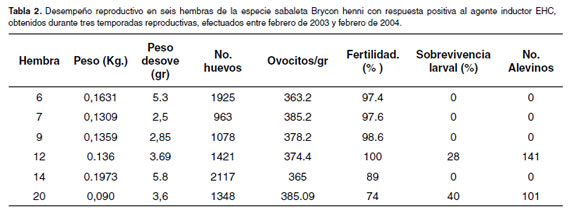
Luego de haber inducido 20 hembras y 20 machos, distribuidos en tres grupos de acuerdo con la temporada reproductiva por año (véase Tabla 1), se presentó respuesta en uno de los desoves realizados en los meses de febrero y marzo de 2003 (n=4) del grupo uno, con un tiempo de latencia de 11 h después de aplicada la dosis resolutoria, pero no se observó fertilidad. En el grupo dos, inducido durante la segunda temporada reproductiva (octubre-noviembre de 2003), se presentó respuesta positiva en el 50% de hembras inducidas (n=12), observándose un período de latencia de 13h, fertilidad de 89% y sobrevivencia larval de 28% (véase Tabla 1), de los cuáles se lograron obtener 141 alevinos en talla comercial (3 cm). En el tercer grupo, correspondiente a la primera temporada reproductiva de 2004 (febrero), 25% hembras presentaron diferentes tonos de color de las hembras ovularon (n=4) con un período de de sus ovocitos: Blancos, traslúcidos y de color latencia de 11 h, 74% de huevos fecundados, 40% de pardo. Los huevos de las hembras que presentaron sobrevivencia larval (véase Tabla 1), de las cuáles se color pardo fueron aquellos los que presentaron obtuvieron 101 alevinos de talla comercial. mejores tasas de fertilidad. Se logró la secuencia del desarrollo embrionario tomando fotografía digital El promedio de huevos por gramo fue de 375 en periodos de 30 minutos y durante 40 horas hasta (véase Tabla 2) y una particularidad es que algunas que eclosionaron las primeras larvas.
Discusión
De las 20 inducciones que se realizaron con la especie Brycon henni, ocho hembras (40%) respondieron bien a la inducción y ovularon con 0.5 y 5 mg por kg de peso vivo de EHC, seis de ellas con huevos fecundados, pero sólo dos tuvieron un desempeño reproductivo completo hasta la fase de alevino, lo que representa un porcentaje muy bajo para concluir que se ha establecido un protocolo de reproducción de la especie. Dosis similares han sido aplicadas en Brycon cephalus por Romagosa (55) con un intervalo de aplicación entre dosis también de 10 h y una única dosis en los machos de 2.5 mg por kg de peso corporal, con respuesta positiva del 45% ( 10 de 22 hembras). Andrade-Talmelli et al (3), en un tratamiento hormonal similar con la especie Brycon insignis, de 77 hembras seleccionadas por características externas, 43 (56%) respondieron positivamente a los tratamientos hormonales. Narahara et al (48), en reproducción inducida de Brycon opalinus con extracto de hipófisis de salmón, de 29 hembras tratadas, 14 (48%) respondieron positivamente al tratamiento hormonal. La respuesta al inductor en estas tres especies del género Brycon fue ligeramente superior a B. henni.
El tiempo de latencia entre la aplicación de la segunda dosis y la ovulación fue calculado en 13 h para el grupo 2 de hembras tratadas, durante la segunda temporada reproductiva (noviembre) de 2003 (n=12) y en 11 h para el grupo 3, tratado durante la primera temporada reproductiva (febrero) de 2004 (n=4) (véase Tabla 1). No se encontraron diferencias estadísticas entre los tres grupos con respecto a la variable latencia (p>0.05). El tiempo de latencia para la sabaleta fue mayor que para otros Brycon, tales como el yamú Brycon amazonicus y la dorada Brycon moorei, que normalmente es de 7 h (18, 40, 64). Arboleda 2006, resultados no publicados, reporta para la especie sabaleta Brycon henni un tiempo de latencia de 10.5 h bajo las condiciones de la estación piscícola del municipio de San Jerónimo, con temperatura promedio del agua de 22 ºC. Debería esperarse bajo estas condiciones, un incremento en el tiempo de latencia muy similar a lo encontrado en el grupo tres, pero con temperaturas superiores.
En Brycon cephalus (55), el momento de la liberación de los huevos y la extrusión ocurrió a las 12 h, en uno de los grupos tratados, en B.opalinus (48) y en B.insignis a las 10 h (3), muy similar a los resultados encontrados en este experimento con B. henni. Yaron en (67), sugiere que la latencia fue siempre más corta en peces tratados con EHC que en peces tratados con otras hormonas, lo cual no coincide con los resultados obtenidos en B. opalinus, B. insignis, B. cephalus y, en B .henni, pero si lo fue para B. amazonicus y para B, moorei. Esto puede ser explicado por el hecho de que la liberación de GnRH de la hipófisis y su respuesta en el ovario es un proceso secuencial, mientras que en los peces inyectados con EHC, la respuesta del ovario a la LH exógena es un proceso simple (31).
El porcentaje de huevos fecundados en B. henni fue alto en el Grupo 2, correspondiente a la segunda temporada reproductiva de 2003 (Octubre-Noviembre) con un promedio de 89% y sobrevivencia larval del 28% (véase Tabla 1), que correspondió con una de las hembras que tuvo un desempeño reproductivo completo hasta alevino. 50% de las hembras de este grupo respondieron positivamente al tratamiento con EHC, mientras que en los Grupos uno y tres correspondiente a la primera temporada reproductiva de 2003 y 2004, solamente el 25% de las hembras respondieron al tratamiento hormonal, con un desempeño reproductivo completo hasta la fase de alevino en el Grupo tres. Las tasas de fertilización y eclosión reportadas en Brycon cephalus (55) fueron del 95% y 85%, en B. insignis (3) de 76% y 42% y con B. opalinus (48) del 90% y del 40%, respectivamente. Desoves experimentales en el ciprínido Chondrostoma nasus, que recibieron inyecciones de extracto de hipófisis de carpa en dosis de 6 mg/kg, tuvieron índices de ovulación altos (86%) y el índice de fertilización fue 74.7% y 83.5%, respectivamente (59).
El porcentaje de respuesta ovulatoria, tiempo de latencia, porcentaje de fertilización y de sobrevivencia embrionaria en barbilla Rhamdia sebae cf, utilizando 5.5 mg.kg-1 de EHC en dosis de 10 y 90%, en intervalo de 12 h, fue 9/12, 7.00 ± 0.04 h, 60 ± 24% y 44 ± 35% respectivamente (22). En cachama blanca Piaractus brachypomus, utilizando una dosis similar, la sobrevivencia embrionaria fue de 89 ± 7.3 (53) y en yamú Brycon amazonicus con una dosis de 5.75 mg/ kg fue de 81 ± 3.1 (18). En este bricónido existen también reportes de sobrevivencia embrionaria de 65 ± 4 (40) y de 83 ± 1 (64). El porcentaje de sobrevivencia embrionaria para B. henni (eclosión), fue bajo de 28% y 40% (véase Tabla 1), para dos de los desoves con respuesta positiva completa. Según los parámetros aplicados a especies de aguas cálidas, este debe ser de 75% (16) y se encuentra por debajo del rango (43 ± 42 a 69 ± 5%) en dos investigaciones realizadas con la especie Brycon amazonicus (60.2 ± 6 y 65.9 ± 5.8) (7).
La mayor dificultad que presenta el proceso de reproducción inducida para la especie Brycon henni, reside en la determinación del punto óptimo de madurez de las hembras, ya que por su pequeño tamaño resulta difícil observar el abultamiento abdominal producido por crecimiento de ovocitos en el saco ovárico. Las hembras se seleccionaron a partir de la biopsia ovárica y posición de vesícula germinal, que correspondían a los estados dos y tres, de acuerdo con la clasificación de Kucharczyk et al (32), de vesícula germinal en migración desde menos de la mitad y más de la mitad del radio, respectivamente, pero no fue registrado el diámetro de los ovocitos. En condiciones de confinamiento, similares a las que se emplearon con B. henni, se recomienda la caracterización de las hembras seleccionadas para inducción a la reproducción, basados en la morfología externa y el diámetro de los ovocitos (55) ; como resultado de ello, fueron registrados diámetros promedio en B. cephalus de 1001.6 µm (55), en B. insignis de 1250 µm (3), en B. opalinus de 1900 µm (48), y también se reportó con esta última , una talla de maduración ovocitaria de 1346.4 µm, alcanzando un máximo de 2570.4µm (24, 25). Marino et al (38), recomiendan la evaluación periódica de la progresión de la maduración gonadal con base en los promedios de distribución del diámetro de los ovocitos de cada hembra, después de la estimulación con EHC en el pez Teleósteo Epinephelus marginatus, con el fin de determinar si ovocitos en diferente estado de desarrollo madurarán simultáneamente en el ovario. Esto sugiere precisar el tipo de ovario que presenta la especie y que probablemente sea asincrónico (metacrónico) o que contiene ovocitos en todos los estadios de desarrollo (63, 69, 70). Lo anterior se sustenta en la observación de una hembra de sabaleta B. henni que presentó madurez gonadal tres meses después de ser desovada, lo que sugiere también la posibilidad de que exista un lote de reserva, similar a lo reportado en B. opalinus (24). Sin embargo, en B. cephalus es reportado un tipo de ovario sincrónico con desove total (55), en B. insignis sincrónico en grupos con desove total (3) y en B. opalinus sincrónico en dos grupos con desove total (24).
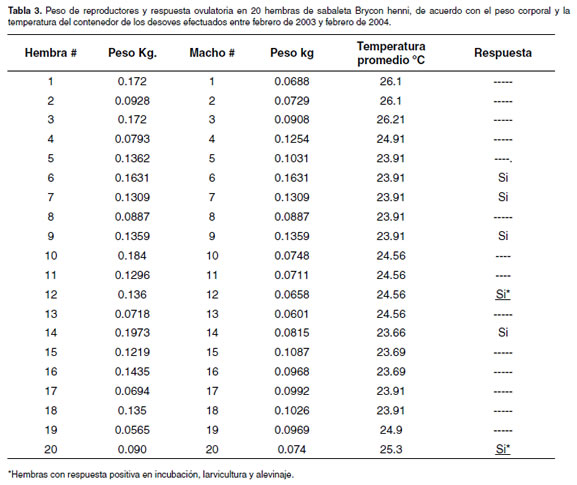
Algunos autores al estudiar los procesos de maduración final ovocitaria (FOM) y espermiación en teleósteos del género Morone (47), determinaron que la falla para alcanzar la FOM son los bajos niveles de hormona luteinizante (LH) en el plasma durante la estación de desove. Como resultado, los cambios esperados en los esteroides inductores de maduración no se observan. Para estas especies, la FOM fue separada en una fase temprana (vesícula germinal en migración) y en una fase tardía (ruptura de vesícula germinal). Para la sabaleta Brycon henni, fue tenido en cuenta el criterio de migración ovocitaria de más del 50% de los ovocitos (16), pero no fueron determinados los perfiles hormonales en el plasma después de la inducción. Probablemente las hembras completaron vitelogénesis pero fallaron en la fase tardía de FOM, por lo que ésta debe ser predicha con un alto grado de exactitud, con el fin de colectar huevos ovulados y proceder exitosamente con la seminación artificial (72, 73). Esto solamente se puede precisar si, adicionalmente al criterio de migración de vesícula germinal, se determinan los niveles de esteroides en plasma. Lo anterior probablemente explica el bajo desempeño reproductivo que presentó la especie después de seleccionar peces en proceso de FOM, con aparentemente buenos resultados en los porcentajes de fertilización pero bajos porcentajes de eclosión (véase Tabla 1). Por ejemplo, los niveles de gonadotropina maduracional o del tipo LH no han sido determinados en peces en el ambiente natural ni en animales sometidos a procesos de domesticación en cautiverio. En Brycon henni, que se adapta fácilmente a temperaturas entre 18 ºC y 28 ºC, no se ha precisado si utiliza los límites inferiores o superiores del amplio rango en que esta especie se mantiene en el medio y pueda alcanzar la FOM. Es probable que utilice, similar a los salmónidos, un estrecho rango de adaptación para la maduración final y el desove durante la época reproductiva y que este factor afecta el desarrollo gonadal. Probablemente los reproductores de la especie, cuando son mantenidos en el rango entre 24-26 ºC de la estación de piscicultura del Nus, experimentan vitelogénesis y alcanzan la fase temprana de FOM (vesícula germinal en migración), como ha sido demostrado para otros teleósteos (72) y como se observó en el momento de realizar las biopsias en las hembras maduras y cuyo efecto más notable es el bajo desempeño reproductivo observado en el porcentaje de eclosión y sobrevivencia larval (véase Tabla 1). En B. henni, cuatro desoves fueron obtenidos con temperaturas inferiores a 24 ºC (véase Tabla 3) y dos por encima de 24 ºC; sin embargo, la temperatura, al parecer, no es un factor limitante en la reproducción dentro de los rangos de confort establecidos para la de B. opalinus (24), puesto que individuos maduros especie (13, 14) y coincidieron con los valores de las fueron capturados en primavera y verano (altas cuencas y microcuencas de donde fueron capturados temperaturas) como en otoño (bajas temperaturas). los reproductores (datos no mostrados) y fueron los La temperatura y el oxígeno disuelto (véase Tabla 3) mismos empleados tanto en incubación, como en en los contenedores del laboratorio, se encontraron larvicultura y alevinaje.
El peso de madurez gonadal encontrado en cautiverio bajo las condiciones de la estación piscícola de San José del Nus, fue entre 53 g y 230 g. Lo que sugiere establecer una relación con la talla y comparar los resultados con otros autores (13, 23, 39). El promedio de ovocitos por gramo de 375.2 unidades (véase Tabla 2), dista mucho de lo reportado para otras especies del género Brycon, por debajo de los valores reportados por Arias (7), (1212 ± 19 y 1409 ± 15) para B. amazonicus, lo cual confirma las características bien diferentes de la especie y la necesidad de incubación con sistemas de bandejas californianas, típicas de huevos bentónicos. Andrade-Talmelli (3), reportan que los huevos de B. insignis son esféricos, demersales y no adhesivos lo que parece ser el caso usual en este género (3).
Este experimento de reproducción inducida de sabaleta ha tenido éxito con respecto a otros reportados por la literatura en el país, en términos de obtención de alevinos, pero todavía no establece un protocolo efectivo para reproducir masivamente la especie, por los cual este trabajo sugiere una falencia que debe estandarizarse mejor, con verificación de parámetros plasmáticos y cuantitativos que permitan continuar las investigaciones y ser exitosas. Otros factores hormonales tienen que estar influyendo en la especie para que se presente disrupción endocrina y disfunción reproductiva, representada en la baja respuesta a la inducción, a pesar de presentar características externas e internas aparentemente favorables. No se conoce con exactitud el comportamiento social en cautiverio y la relación idónea de hembras y machos para esta especie, puesto que se observó en uno de los desoves, al ser utilizadas simultáneamente dos hembras, una de ellas agredió a la otra hasta causarle la muerte.
La temporada reproductiva de la especie se presenta en los meses de noviembre, diciembre, enero y febrero, coincidiendo con algunos autores en lo que respecta a los picos de máxima madurez de los machos y/o de las hembras, lo que refleja probablemente asincronismo en la especie. Sin embargo, se observaron hembras y machos maduros prácticamente durante todo el año en condiciones de cautiverio, lo que sugiere que posiblemente tenga varias temporadas reproductivas o haya maduración por cohortes. Se puede pensar que probablemente otros factores estén influyendo en el comportamiento reproductivo de la especie.
Agradecimientos
A la Corporación para el Desarrollo Sostenible de las Cuencas Rionegro-Nare, CORNARE e ISAGEN, por la financiación de las investigaciones que se llevan a cabo con esta especie en la región del Oriente Medio Antioqueño, quienes con el acompañamiento de la Universidad de Antioquia, a través del Centro de Investigaciones Agrarias y el CODI, hicieron posible la realización de esta investigación.
Referencias
1. Afonso OB, Iwama GK, Smiyh J, Donaldson EM. Effects of the aromatase inhibitor Fadrozole on plasma sex esteroid secretion and ovulation rate in female Coho salmon, Oncorhynchus kisutch, close final maduration. Gen Comp Endocrinol 1999; 113:221-229. [ Links ]
2. Álvarez LF y Duque JP. La sabaleta y su cultivo en estanques. Seminario (Zootecnia); Medellín Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia; 1989. P.1-34. [ Links ]
3. AndradeTalmelli, E.F.A, Kavamoto, EM., Narahara, MY. and Fenerich-Verani, Reprodução induzida da piabanha, Brycon insignis, mantida em cativeiro. Rev Brás Zootec 2002; 31:803-811. [ Links ]
4. Arboleda L, Olivera ÁM, Tabares SJ, Echeverri A, Serna DF Madurez gonadal en hembras de sabaleta (Brycon henni) y su relación con variables medioambientales. Politécnica. 2005; 1:95-103. [ Links ]
5. Arias CJA, Aya BE, Velasco SJ. Inducción reproductiva de yamú con Ovopel©. Memorias II Congreso Col Acuicultura; Villavicencio; 2004. P.131-132. [ Links ]
6. Arias CJA. Estado actual del conocimiento sobre el yamú, Brycon amazonicus. Rev Colomb Cienc Pecu 2006; 19:125-133. [ Links ]
7. Arias CJA, Zaniboni FE, Pardo CSC, Atencio GV, Torres VW. Desarrollo gonadal y efectos de la domesticación y la restricción alimenticia de yamú (Brycon siebenthalae) en cautiverio; Memorias VIII jornada de acuicultura; Villavicencio; 2002; 55-63. [ Links ]
8. Atencio García V. Producción de alevinos de especies nativas Rev MVZ CÓRDOBA 2001; 6:9-14. [ Links ]
9. Atencio García V, Cura E, Cordero A, Pertuz V, Martínez C, et al. Reproducción inducida del bocachico (Prochilodus magdalenae) y el blanquillo (Sorubim cuspicaudus) con OvaprimÒ. IV Seminario Internacional de Acuicultura; Bogotá: Facultad de Medicina Veterinaria y Zootecnia; Universidad Nacional de Colombia; 2004. Memoria digital. [ Links ]
10. Beltrán CN, Beltrán I, Sierra R. Notas preliminares sobre la reproducción y cultivo de la sabaleta (Brycon henni, Eigenman 1913). Bogotá, Instituto de recursos naturales. 1978; 27p. [ Links ]
11. Betancur C, Ortega M, Hernán RQ. Conservación de semen de yamú (Brycon amazonicus), dorada (Brycon moorei sp) y sabaleta (Brycon henni) mediante la técnica de la criopreservacion. Pregrado, Facultad de Ciencias Agrarias, Universidad de Antioquia, Medellín, 2001. 34p. [ Links ]
12. Brzuska E. Artifi cial spawning of herbivorous fish: use of an LHRH-a to induce ovulation in grass carp Ctenopharyngodon idella (Valenciennes) and silcer carp Hypophthalmichthys molitrix (Valenciennes). Aquac Res 1999; 30: 849-856. [ Links ]
13. Builes J, Uran A. Estudio del ciclo sexual de la sabaleta (Brycon henni 1913) su comportamiento y fecundación artificial. Rev Actual biol 1974; 3: 2-12. [ Links ]
14. Builes J, Lara L. Informe sobre las actividades realizadas durante el año 1980 en el programa de piscicultura agrícola de la sabaleta (Brycon henni). Corpourabá, Medellín, 1980; 310-324. [ Links ]
15. Castillo A, Ramírez R, Rodríguez A. Ensayos de reproducción y alevinaje en Yaque Leiarius marmoratus (Gill, 1870) (pisces: siluriformes:pimelodidae), en la Orinoquía Colombiana. IV Seminario Internacional de Acuicultura; Bogotá: Facultad de Medicina Veterinaria y Zootecnia; Universidad Nacional de Colombia; 2004. Memoria digital. [ Links ]
16. Chaparro N. Reproducción Artifi cial y Manipulación Genética en Peces. Barranquilla: Mejoras, 1994. [ Links ]
17. Clavijo A, Aya BE, Arias CJA. Desarrollo embrionario de moneda Metynnis sp. II Congreso Colombiano de Acuicultura. X Jornada de Acuicultura IALL; Villavicencio: Universidad de los Llanos; 2004. P. 39. [ Links ]
18. Cruz Casallas PE, Pardo Carrasco S, Arias Castellanos JA, Lombo-Castellanos PE, Lombo Rodríguez DA, et al. Cryopreservation of Yamú Brycon siebenthalae Milt J World Aquaculture Soc 2004; 35:529-535. [ Links ]
19. Cruz Casallas PE. Manejo hormonal de la función reproductiva de peces tropicales bajo condiciones de cautiverio. Memorias III Seminario Nacional de Ingeniería en Producción Acuícola, Universidad de Nariño. San Juan de Pasto, 31 de Mayo al 2 de Junio de 2006. [ Links ]
20. De Greif S, Montoya F. Estudio genético y bioquímico de cuatro especies del género Brycon de origen colombiano. Trabajo de promoción, Facultad de Ciencias, Universidad Nacional de Colombia, Medellín, 1988. p. 1-85. [ Links ]
21. Devlin R, Nagahama Y. Sex determination and sex differentiation in fish an overview of genetic, physiological, and environmental influences. Aquaculture 2002; 208:191-364. [ Links ]
22. Díaz SE, Arias CJA, Aya BE. Comparación del OVAPRIM y del extracto de hipófisis de carpa (EHC) en la inducción a la ovulación y desove de Rhamdia sebae cf (Pisces Pimelodidae). II Congreso Colombiano de Acuicultura. X Jornada de Acuicultura IALL. 2004; p.118. [ Links ] Flórez Brand, PE Estudio biológico-pesquero preliminar de tres especies icticas del alto del Río Cauca Embalse de Salvajina. Rev Cespedesia 1999; 23:47-61. [ Links ]
24. Gomiero, LM, Braga FMS. Gonadossomatic relation and reproductive strategy of Brycon opalinus (Cuvier, 1819) in the Serra do Mar State Park -Núcleo Santa Virgínia, Atlantic Forest, Brazil Braz J Biol 2007; 67:727-733 [ Links ]
25. Gomiero LM, Braga FMS. Reproduction of Pirapitinga do Sul (Brycon opalinus Cuvier, 1819) in the Parque Estadual da Serra do Mar-Núcleo Santa Virgínia, São Paulo, Brazi J Biol 2007; 67:541-549 [ Links ]
26. Granados H, López A, Galvis D, Ante J, López J et al. Reproducción inducida en: Brycon henni, Eingenman, 1913 (Sabaleta) y Rhamdia wagneri, Gunther, 1869 (Barbudo). Rev Vet y Zoot de Caldas 1986; 5:68-91. [ Links ]
27. Grecco GA, Granda J. Producción de larvas de moscas de establo (insectos Muscidae y Colliphovidae) y su palatibilidad en sabaleta Brycon henni, colocadas en jaulas. Trabajo de grado, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellin. 1989. p.1- 93. [ Links ]
28. Haddy JA, Pankhurst NW. 2000. The efficacy of exogenous hormones in stimulating changes in plasma steroids and ovulation in wild black bream Acanthopagrus butcheri is improved by treatment at capture. Aquaculture 2000; 191:351-366. [ Links ]
29. Harvey B, Hoar W. Teoría y Práctica de la Reproducción Inducida en los Peces. CIID. Otawa, Canadá; 1980 [ Links ]
30. Isaza BM. Ensayo de cultivo de sabaleta Brycon fowleri en Urabá. Trabajo de investigación. Departamento de Biología, Universidad de Antioquia, Medellín, 1983. 83p. [ Links ]
31. Kucharczyk D, Kujawa R, Mamcarz A, Wyszomirska E, Ulikowsky D. Artifi cial spawning of ide (Leuciscus idus) under controlled conditions. EJPAU. Electronic J Polish Agric Universities. 1999; (noviembre de 2007). URL: http://www.ejpau.media.pl/volume2/issue2/fisheries/art-05.html [ Links ]
32. Kucharczyk D, Szczerbowski A, Luczynski MJ, Kujawa R, Mamcarz A, et al. Artifi cial spawning of Eurasian perch, Perca fluviatilis L. using Ovopel. Arch Pol Fish 2001; 9:39-49. [ Links ]
33. Lahnsteiner FR, UrbanyI B, Horvath A, Weisman T. Biomarkers for egg quality determination in cyprinid fish. Aquaculture 2001; 195:331-352. [ Links ]
34. Landinez P, Miguel A. Inducción de la reproducción del yamú (Brycon siebenthalae) a partir de extracto de hipófisis de carpa (EPC); Tesis, Universidad Nacional de Colombia, Bogotá, 1994. 105p. [ Links ]
35. Leonardo A, Romagosa E, Borella M, Batlouni S. Induced spawining of hatchery-raised Brazilian catfish, cachara Pseudoplatystoma fasciatum (Linnaeus, 1766). Aquaculture 2004; 240:451-461. [ Links ]
36. Magallanes H. Evaluación íctica del sistema Oriente Antioqueño. Trabajo de grado, Departamento de Biología, Universidad de Antioquia, Medellín, 1989. 47p. [ Links ]
37. Magallanes MH, Tabarez MM. Informe fi nal del estudio de los efectos del proyecto hidroeléctrico porce II, sobre la fauna íctica. Empresas Publicas de Medellín, gerencia generación de energía, división porce II, Departamento gestión ambiental. 1999. 108 p. [ Links ]
38. Marino G, PaninI E, LongobardI A, Mandich A, Finoia MG, et al. Induction of ovulation in captive-reared dusky grouper, Epinephelus marginatus (Lowe, 1834), with a sustained-release GnRHa implant. Aquaculture 2003; 219:841- 858. [ Links ]
39. Martínez OH, Vásquez GZ. Aspectos reproductivos de la sabaleta Brycon henni (Pisces Characidae), en el embalse La Salvajina, Colombia. Rev Asoc Colomb Ictiol 2001; 4:75-82. [ Links ]
40. Medina Robles, VM, Velasco Santamaría YM, Cruz Casallas PE. Tasa de congelación descongelación de semen de yamú (Brycon amazonicus) empacado en pajillas de diferentes volúmenes y su efecto sobre la calidad espermática postdescongelación. Rev Colomb Cienc Pecu 2005; 18: 331-XX. [ Links ]
41. Millan GJ. Contribución al estudio del comportamiento reproductivo y otros aspectos del ciclo vital de la sabaleta Brycon henni (Eigenman) en el Embalse Calima, Palmira, Valle. Tesis de pregrado Zootecnia, 1981; 112 p. [ Links ]
42. Mojica B.H.O. Efecto de LHRHa2 combinada con Domperidone (Método Linpe) y de la Hipófi sis de Carpa (HC), en la maduración fi nal y ovulación de Curimbatá Prochilodus scrofa (Steindachner, 1881) (PISCES: CHARACIDADE); (Noviembre de 2007) URL: http://www.iiap.org.pe/publicaciones/CDs/MEMORIAS_VALIDAS/contenidos. [ Links ]
43. Molina B. Reproducción inducida de la sabaleta Brycon henni. Seminario como trabajo de grado: Medellín: Facultad de Agronomía, Universidad Nacional de Colombia; 1982. p.1-32. [ Links ]
44. Montoya AF. Descripción anatómica e histológica de las gónadas en Brycon henni (PISCES: CHARACIDAE). Trabajo de grado. Facultad de Ciencias Agrarias, Universidad de Antioquia, Medellín, 2004. 24 p. [ Links ]
45. Montoya A, Carrillo LM, Olivera-Angel M. Algunos aspectos biológicos y del manejo en cautiverio de la sabaleta Brycon henni Eigenmann,1913 (Pisces Characidae). Rev Colomb Cienc Pecu 2006; 19:180-185 [ Links ]
46. Muñoz Cueto J.A, González Martínez D., Z Mora N., Saligaut D., Zanuy S.,Elizur A., et al. New insight in development origins of different GnRH (Gonadotrophin-Releasing hormone) system in Perciform fish:an inmunohistochemical study in the european sea bass (Dicentrarchus labrax). J.Chem. Neuroanat 2004; 28:1-15. [ Links ]
47. Mylonas C, Zohar Y. Endocrine regulation and artificial induction ofoocyte maturation and spermiation in basses of the genus Morone. Aquaculture 2001; 202:205-220. [ Links ]
48. Narahara, MY, Talmeli A, Kavamoto EFE, Godinho HM. Reprodução induzida da pirapitinga do sul, Brycon opalinus, mantida em condições de confinamento. Rev Bras Zootec 2002; 31:1070-1075. [ Links ]
49. Pareja AV. Estudio de la variación genética de Brycon henni del departamento de Antioquia mediante RAPDPCR. Trabajo de grado, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, 2003. 44p. [ Links ]
50. Perdomo JM. La sabaleta (Brycon henni, Eigenman 1913). Observaciones bioecológicas y su importancia como especie de cultivo. Rev div pesq 1978; 11:1-46. [ Links ]
51. Pineda SH, Pareja MD, Olivera-Ángel M. Contribución a la relación taxonómica entre cuatro especies de peces de la familia Characidae mediante el polimorfismo de ADN Amplificado al Azar (RAPD). Rev Colomb Cienc Pecu 2004; 17 Suplemento: 30 - 37 [ Links ]
52. Plata J. Diferenciación sexual y desarrollo de los alevinos de la Sabaleta Brycon henni Eigenman. Trabajo de grado, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, 1977. 22 p. [ Links ]
53. Ramírez MJA, Velasco Santamaría YM, Medina Robles, VM, Cruz Casallas PE. Crioconservación de semen de cachama blanca (Piaractus brachypomus cuvier, 1818): efectos del volumen de empaque y de la sustancia crioprotectora sobre la calidad seminal. Rev Colomb Cienc Pecu 2005; 18:331. [ Links ]
54. Ramos V. Ensayos de reproducción inducida en algunas especies ícticas nativas a partir de protehormonas. Tesis de pregrado, Universidad Jorge Tadeo Lozano, Bogotá, 1978. 70p. [ Links ]
55. Romagosa E, Narahara MY, Borella MI, Fenerich VN. Seleção e caracterização de fêmeas de matrinxã, Brycon cephalus induzidas a reprodução. Bol. Inst. Pesca 2001; 27:139-147. [ Links ]
56. Rosas J, Arana D, Cabrera T, Millán J, Darryl J. The potencial use of the Caribbean Flounder Paralichthys tropicus as an aquaculture species. Aquaculture 1999; 176: 51-54. [ Links ]
57. Solano J. M. Reproducción inducida de la dorada (Brycon moorei sinuensis) y el bocachico(Prochilodus magdalenae). Montería: CINPIC/Universidad de Córdoba, 1992; 5p. [ Links ]
58. Sotelo FGA, Arias CJA, Aya BE. Inducción a la ovulación y el desove de la barbilla Rhamdia sebae c.f (Pisces Pimelodidae), con Ovopel. II Congreso Colombiano de Acuicultura. X Jornada de Acuicultura IALL; Villavicencio, Universidad de los Llanos, 2004. p. 1-132. [ Links ]
59. Szabo´ T, Medgyasszay C, Horváth L. Ovulation induction in nase (Chondrostoma nasus,Cyprinidae) using pituitary extract or GnRH analogue combined with domperidone. Aquaculture 2002; 203:389-395. [ Links ]
60. Tabares SCJ. Evaluación del efecto de algunos iones sobre la activación de la movilidad espermática y el potencial de membrana en Brycon henni (Eigenmann 1913), tesis, Facultad de Ciencias Agrarias, Universidad de Antioquia, Medellín, 2005; 68p. [ Links ]
61. Tabares CJ, Montoya AF, Arboleda L, Echeverri A, Restrepo LF, et al. Efecto de la pluviosidad y el brillo solar sobre la producción y características del semen en Brycon henni (Pisces Characidae). Rev Biol Trop 2006; 54:179-187. [ Links ]
62. Tabares J, Ruiz T, Arboleda L, Olivera Angel M. Effect of some ions on sperm activation in Brycon henni (Eigenmann 1913). Efecto de algunos iones sobre la activación espermática en Brycon henni (Eigenmann 1913). Acta biol Colomb 2007 ;12: 87-98. [ Links ]
63. Vasquez W. Efeito de dietas com níveis crescentes de proteína e energía na evolucao ovocitaria da pirapitinga, Piaractus brachypomus (CUVIER, 1818). Tesis de maestria, Centro de Ciencias Agrárias, Universidad Federal de Santa Catarina, Brasil, 1994. 86 p. [ Links ]
64. Velasco Santamaría YM, Medina Robles VM, Cruz Casallas PE. Cryo -preservation of yamú (Brycon amazonicus) sperm for large scale fertilization. Aquaculture 2006; 256: 264-271. [ Links ]
65. Weber GM, King VW, Clark RW, Hodson RG, Sullivan CV. Morpho-physiological predictors of ovulatory succes in captive striped bass (Morone saxatilis). Aquaculture 2000; 188:133-146. [ Links ]
66. Woynarovich E. La Propagación de Peces. Informe Técnico Nº 72. Venezuela. 1977. 48 p. [ Links ]
67. Yaron Z. Endocrine control of gametogenesis and spawing induction in the carp. Aquaculture1995; 129:49-73. [ Links ]
68. Zaniboni FE. Priming hormone administration to induce spawning of some brazilian migratory fish. Rev Bras Biol 1996; 56:655-659. [ Links ]
69. Zanuy S, Kah O., E.Mañanos, I.Anglade y M.Carrillo . Distribution of salmon gonadotrophin releasing hormone in the brain and pituitary of the sea bass (Dicentrarchus labrax). An immunocytochemical and immunoenzymoassay study. Cell Tissue Res 1991; 266:129-136. [ Links ]
70. Zanuy S, Carrillo M. Structural and functional effects of early exposure to estradiol-17 and 17-ethynylestradiol on the gonads of the gonochoristic teleost Dicentrarchus labrax. Fish Physiol Biochem 1998; 18: 37-47. [ Links ]
71. Zapata L, Vanegas R. Aspectos importantes sobre la reproducción inducida y el metabolismo de las gónadas de sabaleta Brycon henni. Trabajo de grado, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, 1993. 67p. [ Links ]
72. Zohar Y, Mylonas C. Endocrine manipulations of spawning in cultured fish: from hormones to genes. Aquaculture 2001; 197: 99-136. [ Links ]
73. Zohar Y, Zanuy S. Spawning induction of individual European sea bass females (Dicentrarchus labrax) using different GnRHa-delivery systems. Aquaculture 2001; 202: 221-234. [ Links ]

