










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.24 no.2 Medellín Apr./June 2011
Body weight is inversely associated with ammonia excretion in red tilapia (Oreochromis sp)¤
El peso corporal esta inversamente relacionado con la excreción de amonio en tilapia roja (Oreochromis sp)
O peso corporal está inversamente relacionado com a excreção de amônia em tilápia vermelha (Oreochromis sp)
Rubén D Valbuena-Villarreal1*, Biol, esp, MS; Walter Vásquez-Torres2, Biol, MS, PhD.
1 Profesor Universidad Surcolombiana Facultad de Ciencias Exactas y Naturales, Neiva, Colombia.
2 Profesor Instituto de Acuicultura de la Universidad de los Llanos, Sede Barcelona, Km 12 vía Puerto López, Villavicencio, Meta, Colombia.
(Recibido: 8 diciembre, 2010; aceptado: 2 mayo, 2011)
¤ Para citar este artículo: Valvuena-Villareal RD, Vásquez-Torres W. El peso corporal esta inversamente relacionado con la excreción de amonio en tilapia roja (Oreochromis sp).
* Autor para correspondencia: Rubén Darío Valvuena Villareal. Universidad Surcolombiana Facultad de Ciencias Exactas y Naturales, Neiva, Colombia. E-mail: *rubendario@usco.edu.co.
Summary
To meet the increasing demand for fish, aquaculture has expanded through intensive farming that requires large amounts of feed. This can cause environmental and economic difficulties because fish can only retain 20 to 30 % of the ingested nitrogen (N). The remaining fraction is excreted as ammonium and can accumulate in water, thus becoming detrimental to fish performance. This experiment used red tilapia (Oreochromis sp) to determine the effects of temperature, body weight, and dietary crude protein (CP) on ammonium excretion. A factorial AxBxC design was used to compare total ammonium excreted. Factor A is defined as fish body weight (P). Fish weights were: P1= 100 ± 6.2, P2= 249 ± 4.2 y P3= 498 ± 4.7g. Factor B is defined as dietary protein levels (D1, D2 y D3), and factor C as water temperature (T1 and T2). Increased dietary protein levels, as well as water temperature caused significant increases in ammonium excretion (p<0.05). Body weight showed an inverse relationship with ammonia excretion. The 100 g fish at 26 ºC consuming a 25.6% CP diet excreted 468.8 ± 2.2 g N- NH4+ /day/kg, while 498 g fish under the same diet and temperature, excreted 356.3 ± 1.6 mg N- NH4+ /day/kg. This study suggests that dietary factors and environmental variables can alter ammonia excretion in fish, and thus affect of ammonia toxicity in the surrounding water.
Key words: ammonium excretion rate, body weight, diet protein, Oreochromis sp, red tilapia, water temperature.
Resumen
Con el fin de atender la creciente demanda del mercado, la acuicultura se ha expandido mediante la implementación de sistemas de cultivo intensivo que requieren suministro de altas cantidades de alimento. Estas condiciones pueden generar dificultades ambientales y económicas debido a que los peces solamente pueden retener 20 a 30% del nitrógeno (N) ingerido; la proporción restante, que es excretada en la forma de amonio, se puede acumular en al agua llegando a ser perjudicial para los peces. El presente experimento fue realizado con el fin de determinar el efecto de la temperatura, peso corporal y niveles proteicos de las dietas sobre las tasas de excreción de amonio total en tilapia roja (Oreochromis sp). Para la cuantificación del amonio total se aplicó un diseño completamente al azar con arreglo factorial AxBxC, siendo el factor A = peso corporal de los peces (P1= 100 ± 6.2, P2= 249 ± 4.2 y P3= 498 ± 4.7 g), B= dietas experimentales (D1, D2 y D3) y C = temperatura del agua (T1 Y 72). El aumento de los niveles de proteína de la dieta, así como de la temperatura del agua, ocasionaron incrementos significativos en las tasas de excreción de amonio total (p<0.05). El peso corporal presentó una relación inversa respecto a la tasa de excreción de amonio. Peces de 100 g a 26 ºC y una dieta con un nivel proteico del 25.6% excretaron 468.8 ± 2.2 g N-NH4+ al día/kg de peso vivo, en tanto que peces de 498 g bajo la misma dieta y la misma temperatura, excretaron 356.3 ± 1.6 mg N-NH4+ al día/kg de peso vivo.
Palabras clave: peso corporal, proteína dietaría, tasa de excreción de amonio, temperatura del agua, tilapia roja (Oreochromis sp).
Resumo
Para atender à crescente demanda do mercado, a aquicultura tem se expandido através da implementação de sistemas de agricultura intensiva que exigem a entrega de grandes quantidades de alimentos. Essas condições podem gerar dificuldades ambientais e económicas porque o peixe só pode conter 20 a 30% de nitrogênio (N) ingeridos, o resto é excretada na forma de amônia pode acumular-se na água tornando-se prejudicial para os peixes. Este experimento foi conduzido para determinar o efeito da temperatura, peso corporal e níveis de proteína na dieta sobre a taxa de excreção de amônia total em tilápia vermelha (Oreochromis sp.) Para a quantificação de amônia total foi aplicado um delineamento inteiramente casualizado com AxBxC factorial, sendo o factor A = peso corporal dos peixes (P1 = 100 ± 6.2, P2 = 249 ± 4.2 e P3 = 498 ± 4.7 g), B = dietas experimentais (D1, D2 e D3) e C = temperatura da água (T1 e 72). Aumento dos níveis de proteína na dieta, bem como a temperatura da água, resultou em aumentos significativos nas taxas de excreção de amônia total (p<0.05). O peso corporal mostrou uma relação inversa com a taxa de excreção de amônia. Peixes de 100 g mantidos em 26 ºC e uma dieta com teor proteico de 25.6% excretaram 468.8 ± 2.2 g de N-NH4 + por dia / kg de peso corporal, enquanto os peixes de 498 g com a mesma dieta e à mesma temperatura, excretada 356.3 ± 1.6 mg N-NH4 + por dia / kg de peso vivo.
Palavras-chave: peso corporal, proteína, taxa de excreção de amônia, temperatura da água.
Introducción
La producción mundial de la acuicultura de peces de agua dulce denota un crecimiento acelerado durante los últimos años, pasando de una producción de 24 millones de TM en el 2002 a 31.6 millones en el 2006 que equivalen a un incremento de aproximadamente 32% en un lapso de cinco años (Josupeit, 2009). Con el propósito de atender la creciente demanda de tales productos, la industria acuícola ha incorporado nuevos procesos y sistemas de cultivo, entre ellos la intensificación (Caglan, 2005). Estos sistemas requieren el suministro de grandes cantidades de alimento para proveer a los organismos cultivados proteína y demás nutrientes necesarios para lograr máximo crecimiento en el menor tiempo y costo posibles.
Tomando en cuenta que los peces solamente retienen entre el 20 y el 30% del nitrógeno (N) ingerido con el alimento (Green y Boyd 1995), la proporción restante, que no es depositada para crecimiento del organismo, es eliminada al medio acuático en la forma de desechos nitrogenados que tienden a acumularse en la columna de agua y en los sedimentos, contribuyendo a procesos de eutrofización (Jiménez-Montealegre et al., 2005; Vásquez-Torres, 2007). La intensificación tiende a reducir la calidad de las aguas de cultivo y con frecuencia, expone a los animales al efecto de altas concentraciones de amonio, causando en muchos casos lesiones y/o la aparición de enfermedades que reducen de forma significativa la productividad de dichos sistemas (Hargreaves, 1997; Caglan, 2005).
En teleósteos, el amonio es el principal producto del metabolismo de proteínas correspondiendo a aproximadamente 60- 80% del total del N excretado (Shuenn-Der et al., 2002). Se considera que la cuantificación de las tasas de excreción de amonio (EA) puede ser utilizada como indicador del efecto de factores nutricionales y ambientales en el metabolismo de las proteínas en peces (Kelly y Kohler, 2003). Según Shuenn-Der et al. (2002), la excreción del amonio para cada especie es dependiente de los niveles y calidad de la proteína dietaria, del tamaño corporal y de factores ambientales como temperatura y pH del agua.
Para especies dulceacuícolas de hábito omnívoro como la tilapia roja (Oreochromis sp), son pocas las informaciones disponibles en la literatura científica sobre excreción de nitrógeno en la forma de amonio. Así, el propósito del presente estudio fue investigar el efecto de las variables temperatura del agua, peso corporal y niveles de proteína de la dieta sobre la excreción de amonio en tilapia roja (Oreochromis sp), en un periodo de 24 horas.
Materiales y métodos
Animales experimentales e infraestructura
Los experimentos fueron realizados en las instalaciones de la Estación Piscícola de Piedra Pintada (Huila-Colombia). Un grupo de 300 machos revertidos de tilapia roja de (P1 = 100 ± 6.2, P2 = 249 ± 4.2 y P3 = 498 ± 4.7 g) fueron distribuidos según su peso, en tres tanques circulares de cemento de 2000 L abastecidos con un fl ujo de agua constante (3 L/seg) y aireación permanente. Los peces fueron adaptados a las condiciones experimentales por un periodo de 15 días. Durante los primeros diez días se suministró DRS de 33.6% de proteína bruta (Tabla 1), una vez al día (10 a.m.) a una tasa del 1% del peso vivo, considerada como tasa de mantenimiento y durante los otros cinco las correspondientes dietas experimentales. Para la realización de las mediciones de excreción fueron utilizados nueve tanques circulares en fibra de vidrio con un volumen efectivo de 200 L cada uno y aireación permanente para mantener niveles de oxígeno superiores a 60% de saturación; estos tanques fueron abastecidos con agua reciclada a través de dos biofiltros para eliminar amonio.
Los diferentes experimentos se realizaron a dos temperaturas del agua, T1=26 °C y T2=30 ºC. Durante el desarrollo de los experimentos se registraron en el agua los siguientes parámetros de calidad: temperatura 26°C ± 0.4ºC, pH 6.5 ± 0.4, oxígeno disuelto 6.5 ± 0.2 mg/L y amonio <0.001 mg/L. En los ensayos a temperatura del agua de 30ºC se utilizaron calentadores eléctricos dispuestos en el sistema de recirculación. Estos parámetros fueron medidos utilizando una sonda multiparamétrica YSI 556. (YSI Incorporated, Yellow Springs, OH USA). Para cuantificar el amonio disuelto en el agua se empleó el método fotométrico utilizando el Test Amonio Spectroquant® Ref. 1.14752.000, medido en el Spectroquant® NOVA 60 de Merk (Merck KGaA, Darmstadt, Alemania), con una sensibilidad de +0.001 - 5.00 mg/L N-NH4.
Dietas experimentales
Tomando como base la Dieta Referencia Semipurificada propuesta por Vásquez et al. (2002) se formularon tres dietas experimentales con 25, 30 y 35% de PB, respectivamente D1, D2 y D3 (Tabla 1). Tomando en cuenta la metodología propuesta por Vásquez-Torres (2001), los tres niveles se lograron variando la proporción de las fuentes proteicas (caseína y gelatina) manteniendo las proporciones entre sí para garantizar el balance de aminoácidos esenciales en las tres dietas; también se modificaron los porcentajes de las fuentes energéticas (dextrina y aceites) para tratar de conservar el carácter isocalórico en las dietas (aproximadamente 3050 kcal/kg ED); los ajustes finales de masa se lograron modificando los niveles de celulosa y carboximetil-celulosa.
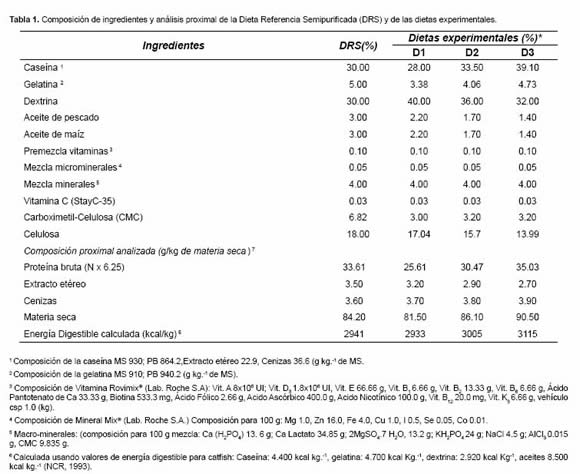
Los análisis de composición proximal fueron realizados en el Laboratorio de Alimentación y Nutrición de Peces (LEANP) del IALL-Unillanos siguiendo normas estándar de la AOAC, 1995. Las raciones fueron fabricadas utilizando ingredientes semipurificados y una micro extrusora de laboratorio (Exteec, Riberão Preto -Brasil)
Procedimiento experimental
Fueron realizados seis experimentos, cada uno con tres tratamientos por triplicado. En cada ensayo los peces fueron colocados en los tanques de 200 L a una densidad de 25 g de pez en número que varió, de acuerdo con su peso para ajustar una biomasa de aproximadamente 5 kg por unidad experimental. El protocolo para todos los experimentos fue el siguiente: Los peces fueron trasladados de las piletas circulares de acondicionamiento a los tanques experimentales en las primeras horas de la mañana y luego a las 10 a.m., se les suministró una ración equivalente al 1% de la biomasa con la dieta experimental en estudio. Después de 15 minutos se aumentó el recambio de agua para renovarla completamente y eliminar heces y alimento no consumido; luego de 5 minutos se suspendió el flujo e inmediatamente se tomó la primera muestra para determinar el contenido de amonio inicial en el agua y después de dos horas se colectó una segunda muestra; el amonio excretado durante ese periodo de tiempo fue la diferencia entre la primera y la segunda muestra.
Este procedimiento de renovación del agua, suspensión del flujo y muestreos al inicio y al final de cada periodo de dos horas, se repitió durante 24 horas. De acuerdo con los resultado obtenidos por David (2010) trabajando con cachama blanca bajo condiciones experimentales similares, dos horas dan un margen suficiente para que los niveles de amonio acumulados en al agua no alcancen valores críticos para los peces. El amonio total excretado en un periodo de 24 horas (mg-NH4+/día/kg de PV) fue el resultado de la sumatoria de los 12 valores obtenidos en las lec turas para cada una de las tres replicas por tratamiento. Esta misma metodología fue aplicada en todos los seis experimentos y fueron realizados en secuencia utilizando los mismos tanques pero diferentes animales, primero a temperatura del agua.
Análisis estadístico
Se aplicó un modelo completamente al azar con arreglo factorial A x B x C (2 x 3 x 3) con tres replicas donde A correspondió a la temperatura (26 y 30 °C), B al peso corporal de los animales (100 ± 6.2, 249 ± 4.2 y 498 ± 4.7 g) y C a las dietas experimentales (D1, D2 y D3). Se efectuaron comparaciones de medias empleando el test de Tukey (p<0.05). Adicionalmente se establecieron ecuaciones de regresión para predecir tanto la producción de amonio en función del peso corporal, de la temperatura del agua y del nivel de proteína de las dietas. El ajuste de dichos modelos se realizó mediante el procedimiento stepwise del prog reg del programa estadístico SAS versión 8.02 para Windows (1999-2001 por SAS Institute Inc, Cary, Nc. USA). Los demás análisis se realizaron mediante el procedimiento GLM de SAS. El nivel de significancia empleado fue del 5%.
Resultados
Como se observa en la tabla 2, el peso corporal, la temperatura y el nivel de proteína de la dieta presentaron interacción sobre la excreción de amonio en tilapia roja (p<0.05).
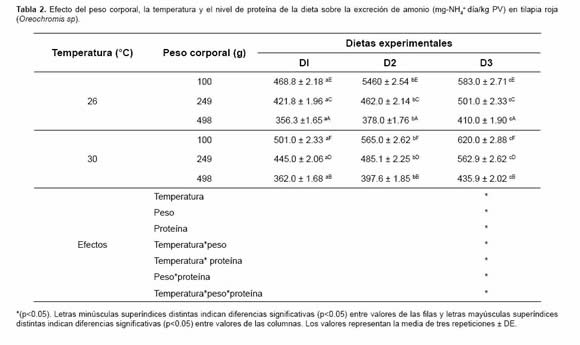
Los efectos simples de las tres variables las tasas de EA en tilapia roja (Oreochromis sp) en analizadas fueron significativamente diferentes diferentes niveles de proteína en la dieta y pesos (p<0.05). Igualmente la interacción entre pares de factores así como de los tres factores simultáneamente, también fue signifi cativa indicando que sus efectos sobre la excreción de amonio no son aislados y son interdependientes.
Efecto del nivel de proteína sobre la tasa de excreción de amonio
El efecto del aumento de los niveles de proteína en la dieta sobre las tasas de EA se presenta en la figura 1. Se observaron incrementos lineales signifi cativos (p<0.05) en las tasas de EA cuando se aumentaron los niveles de proteína en diferentes pesos corporales para cada una de las temperaturas del agua estudiadas.
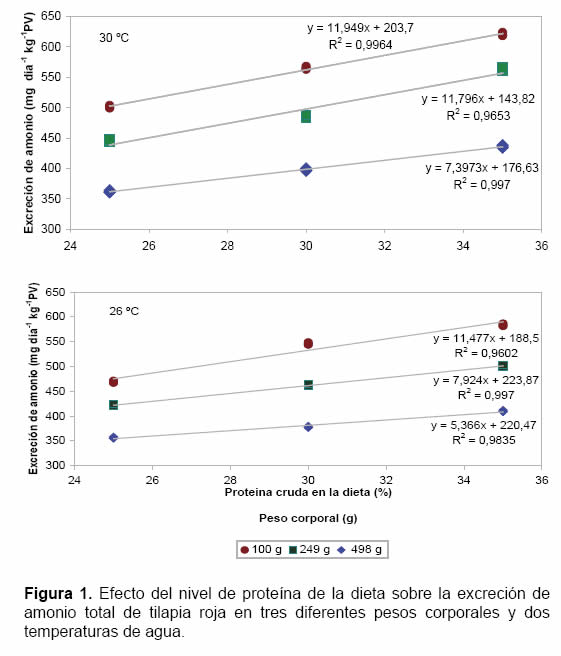
Efecto de la temperatura sobre la tasa de excreción de amonio en tilapia roja
Aumentos en la temperatura del agua ocasionaron incrementos signifi cativos (p<0.05) en las tasas de EA en tilapia roja (Oreochromis sp) en diferentes niveles de proteína en la dieta y pesos
corporales (Tabla 2 y Figura 2).
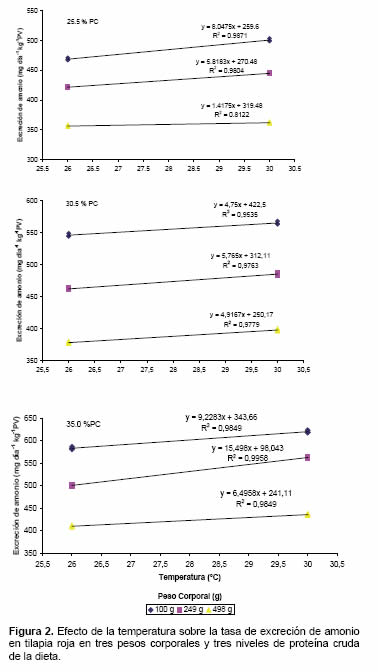
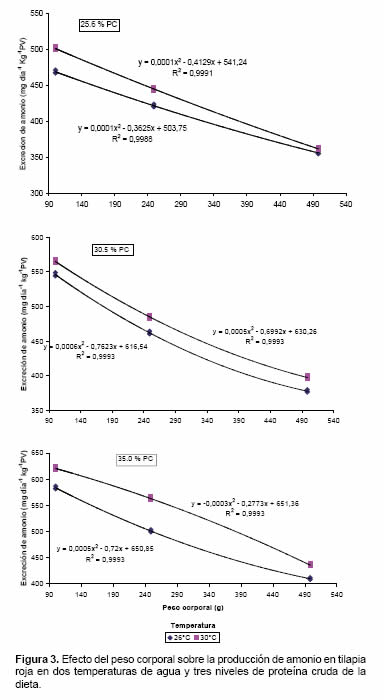
Efecto del peso corporal sobre la tasa de excreción de amonio en tilapia roja
Se observó que los peces de menor peso corporal excretaron mayor cantidad de amonio por unidad de peso que los animales mayores (comportamiento cuadrático decreciente). Sin embargo dichas tasas de EA también dependieron de la temperatura del agua y del nivel de proteína de la dieta (Figura 3).
Discusión
Durante el proceso metabólico ocurrido en los peces, la degradación de las proteínas y ácidos nucleicos, endógenos o dietéticos, llevan a la formación y excreción de diferentes compuestos
nitrogenados siendo el amonio el principal producto (Kaushik et al., 1991); por lo tanto, las mediciones y/o cuantificaciones del amonio han sido usadas como indicadoras de los efectos de varios factores ambientales y nutricionales en el metabolismo de la proteína y pueden dar una visión sobre el balance del nitrógeno en los peces (Perera et al., 1995).
Efecto de nivel de proteína en la dieta sobre la tasa de excreción de amonio
Buttle et al. (1995), describen la existencia de una relación determinante entre tasas de excreción de amonio y proteína de la dieta en teleósteos; el nitrógeno amoniacal proviene de la remoción
del grupo amino, (-NH2), de los aminoácidos del pool que inician su catabolismo. Un exceso de proteína digestible y/o un desbalance del perfil de aminoácidos de la proteína presente en el alimento,
generan un aumento del pool de aminoácidos disponibles y estimulan un mayor catabolismo y por tanto, una mayor excreción de amonio.
Tanto en crustáceos como en teleósteos de agua dulce y marinos se ha demostrado una relación directa entre nivel de ingestión de N (correspondiente a la proteína dietaria) y la excreción de N-amonio (Kaushik et al., 1991; Engin y Carter, 2001), observándose incrementos en las tasas de excreción de nitrógeno total en la medida en que se aumenta la ingestión de nitrógeno (N) (Chakraborty y Chakraborty, 1998; Ballestrazzi et al., 1994). Engin y Carter (2001) reportan que la mayoría del nitrógeno excretado por los peces es derivado de la desaminación de los aminoácidos que ingresan al organismo con la proteína de la dieta.
Los incrementos en las tasas de EA con el incremento en los niveles de proteína dietaria observados en Oreochromis sp del presente experimento fueron semejantes a los descritos para otras especies. Caulton (1978) señala que en Tilapia rendalli (Boulenger) la tasa de excreción de amonio es dependiente de la cantidad de proteína en el alimento; la misma relación es descrita por Chakraborty et al. (1992) para carpa común. Desempeño similar de relación directa entre tasas de EA y niveles de proteína son reportados por Chakraborty y Chakraborty (1998) en alevinos Labeo rohita y por García et al. (1999) y Engin y Carter (2001) en Anguilla anguilla; coincidiendo con lo anterior Shuenn-Der et al. (2002) observaron una relación de tipo lineal entre el nivel de proteína en la dieta y la cantidad de amonio postprandial excretado por Bidyanus bydianus durante 24 horas Buttle et al. (1995), quienes trabajaron con Clarias gariepinus encontraron que el patrón de excreción de amonio presentaba una relación directa con el contenido de proteína de la dieta.
Otras especies reportadas con el mismo patrón de excreción de amonio frente al aumento de proteína son: Oncorhynchus mykiss (Cheng et al., 2004), Ctenopharyngodon idella (Carter y Brafield, 1992), Anguilla Anguilla (Engin y Carter, 2001) Sciaenops ocellatus (Webb y Gatlin, 2003), silver perch (Shuenn-Der et al., 2002), Sparus aurata (Martínez, 2002) y Chanos chanos (Sumagaysay-Chavoso, 2003). El incremento en la producción de amonio como resultado del aumento en los niveles de proteína en la dieta presentado por tilapia roja en la presente investigación, al igual que para las demás especies reportadas por diferentes autores, está relacionado con el incremento en el catabolismo de las proteínas utilizadas como fuente de energía.
De otro lado, Dosdat et al. (1996) manifiestan que el consumo de proteína, tanto en calidad como en cantidad, es el principal factor que afecta la producción de desechos nitrogenados. El conocimiento de las tasas EA en función del nivel de la proteína, para el caso de tilapia y la mayoría de los peces de cultivo, puede ser considerado un factor determinante para desarrollar estrategias que permitan hacer de la acuicultura una actividad rentable y sostenible desde el punto de vista económico, social y ambiental; la utilización de dietas con niveles de proteína por encima de los requerimientos de la especie, cualquiera que esta sea, puede hacer que los cultivos sean más costosos y causen un mayor impacto ambiental. Los resultados obtenidos en la presente investigación confirman este razonamiento; tal es el caso de los peces de 100 g alimentados con un nivel de proteína de 35% a 30 ºC que excretaron aproximadamente un 20% más de amonio comparado con los peces alimentados con la dieta de 25.6% de proteína, nivel que está más próximo al requerimiento de proteína para esta especie (Bahnasawy, 2009; Zimmermanns y Fitzsimmons, 2004).
Efecto de la temperatura sobre la tasa de excreción de amonio en tilapia roja
Zakes et al. (2007) y Person-Le et al. (2004) afirman que generalmente la temperatura tiene un gran efecto sobre la tasa metabólica de los peces y por lo tanto ha sido relacionada con la EA de dichos organismos. Esto es explicado por Fu-Guang et al., (2009), quienes describen cómo, a elevadas temperaturas del agua para los procesos metabólicos normales del pez se requiere mayor cantidad de energía, la cual es obtenida por oxidación de los esqueletos de carbono que resultan de procesos de desaminación de aminoácidos dietarios; de esta manera, a mayor demanda de energía, mayor la tasa de desaminación y consecuentemente mayor la tasa de excreción de amonio.
Como se observa en la tabla 2, los efectos simples del factor temperatura fueron estadísticamente diferentes (p<0.05) entre los dos tratamientos, tales diferencias en las tasas de EA fueron del orden del 10%, considerando los efectos combinados con cada una de las otras dos variables en estudio (nivel de proteína y peso corporal) este porcentaje de diferencias fue semejante entre tratamientos.
De manera general, estos resultados concuerdan con los descritos para otras especies (Person- Le et al., 2004). Relaciones similares entre tasas de excreción y temperatura han sido reportadas para Epinephelus areolatus (Leung et al., 1999), Perca fluviatilis (Zakes et al., 2003) y Miichthys miiuy (Zheng et al., 2008).
Efecto del peso corporal sobre la tasa de excreción en tilapia roja
De acuerdo con Zhang et al. (2004) las tasas de EA son dependientes del tamaño de los peces, siendo proporcionalmente menor cuando mayor sea el peso de los animales. El decrecimiento de la tasa de EA con el incremento de peso del cuerpo refleja una tasa metabólica más baja en animales grandes (Zakes et al., 2003). En el presente experimento tal desempeño pudo ser verificado encontrándose diferencias de aproximadamente 10% en las tasas de excreción de amonio entre los animales de 100 y 249 g y de 30% entre los menores (100 g) y los mayores (498 g) (Figura 3). Adicionalmente, se observó que tales diferencias se mantuvieron al comparar el efecto de la temperatura y de los niveles de proteína en la dieta. La mayor tasa de excreción registrada en este experimento (620.0 ± 2.88 mg-NH4+ día/kg PV fue observada en los peces de 100 g alimentados con la dieta del 35% de PB y temperatura de 30 ºC y la menor, 356.3 ± 1.65 mg4+ día/kg PV en los peces de 498 g alimentados con la dieta de menor contenido de proteína y a una temperatura de 26 ºC.
El efecto del peso corporal sobre las tasas de EA tanto en tilapia como en las demás especies puede ser explicado por cambios fisiológicos como el tipo de metabolismo que ocurre durante la ontogénesis de los organismos; además estos pueden estar relacionados con el desarrollo de la masa muscular y los cambios en el coeficiente superficie/volumen de los órganos respiratorios (Post y Lee, 1996).
Con respecto a los reportes de la literatura, las tendencias de excreción de amonio en función de las variables analizadas en este experimento, son semejantes a las descritas para las especies Epinephelus areolatus (Leung, 1999), Sparus aurata (Martínez, 2002) y Perca fluviatilis, (Zakes et al., 2003), entre otras. Los resultados de la presente investigación permiten confirmar que en la tilapia roja, al igual que en otras especies de peces, las tasas de EA dependen significativamente de la temperatura del agua, del peso corporal de los animales y de los niveles de proteína dietaria, siendo este último parámetro el que tiene mayor influencia. Resultados obtenidos en investigaciones con otras especies han mostrado que además de los factores en este trabajo considerados, otros como tasa de alimentación, número y periodicidad de las raciones también afectan la EA. Para propósitos de comparación entre los seis experimentos realizados en esta investigación, a los peces solamente se les suministró una ración al día a una tasa fija de 1% de la biomasa. Zdzislaw-Zake et al. (2006) observaron que en Tinca tinca la EA se cuadriplicaba al aumentar de 0.2 a 0.8% de la biomasa el tamaño de la ración e igualmente, encontraron diferencias significativas al pasar de una ración a tres por día.
De otro lado, los resultados obtenidos y el mismo conocimiento de los patrones de comportamiento de las tasas de excreción frente a las variables estudiadas, pueden se utilizadas como herramientas valiosas para optimizar el uso de la proteína en las raciones que se utilizan para alimentar a la Tilapia roja, especialmente en cultivos intensivos en jaulas y en estanques, procurando no exceder el nivel de 32%, tal como lo recomiendan Zimmermann y Fitzsimmons (2004). De esta manera se logrará reducir costos de producción y minimizar el impacto de la actividad sobre el recurso agua.
Agradecimientos
A CENTRACAFE por su valiosa colaboración con el aporte de las instalaciones de la Estación Piscícola Piedra Pintada centro en donde se realizó la presente investigación. Igualmente al MADR por el apoyo financiero a través del proyecto 018/05 IICA/UNILLANOS.
Referencias
1. AOAC International (US). Official methods of analysis of AOAC International. 2 vols. 16th edition. Arlington: Association of Analytical Communities; 1995.
[ Links ]2. Bahnasawy MH. Effect of dietary protein levels on growth performance and body composition of mono sex nile tilapia, Oreochromis niloticus L reared in fertilized tanks. Pakistan J Nutr 2009; 5:674-678.
[ Links ]3. Ballestrazzi R, Lanari D, Mion A. The effect of dietary protein level and source on growth, body composition, total ammonia and reactive phosphate excretion of growing sea bass (Dicentrarchus labrax). Aquaculture 1994; 127:197-206.
[ Links ]4. Buttle LG, Uglow RF, Cowx IG. Effect of dietary protein on the nitrogen excretion and growth of the African catfish, Clarias gariepinus. Aquat Living Resour 1995; 8:407-414.
[ Links ]5. Caglan AKB. Investigation of acute toxicity of fluthrin on tilapia fry (Oreochromis niloticus L. 1758).Environ Toxicol Pharmacol 2005; 20:279-282.
[ Links ]6. Carter CG, Brafield AE. The bioenergetics of grass carp, Ctenopharyngodon idella (Val.): the influence of body weight, ration and dietary composition on nitrogenous excretion. J Fish Biol 1992; 41:533-543.
[ Links ]7. Caulton MS. The importance of habitat temperature for growth in the tropical cichlid, Tilapia rendalli. J Fish Biol 1978; 13:99-112.
[ Links ]8. Chakraborty SC, Chakraborty S. Effect of dietary protein level on excretion of ammonia in Indian major carp, Labeo rohita, fingerlings. Aquacult Nutr 1998; 4:47-51.
[ Links ]9. Chakraborty SC, Ross LG, Ross B. The effect of dietary protein level and ration level on excretion of ammonia in common carp, Cyprinus carpio. Compar Biochem Physiol. Part A: Physiology 1992; 103:801-808.
[ Links ]10. Cheng ZJ, Hardy RW, Huige NJ. Apparent digestibility coefficients of nutrients in brewer's and rendered animal by-products for rainbow trout (Oncorhynchus mykiss) (Walbaum). Aquacult Res 2004; 35:1-9.
[ Links ]11. David C. Cuantificación de los niveles de excreción de nitrógeno amoniacal en función del nivel de proteína en la dieta y la masa corporal, en cachama blanca (Piaractus brachypomus) (Cuvier 1818) bajo condiciones de laboratorio. Tesis de Maestría en Acuicultura. Universidad de los Llanos/ IALL, Villavicencio, Colombia.
[ Links ]12. Dosdat A, Servais F, Metáiller R, Huelvan C, Desbruyeres E. Comparison of nitrogenous losses in five teleost fish species. Aquaculture 1996; 141:107-127.
[ Links ]13. Engin C, Carter G. Ammonia and urea excretion rates of juvenile Australian short-finned eel Anguilla australis australis as influenced by dietary protein level. Aquaculture 2001; 194:123-136.
[ Links ]14. Fu-Guang L, Shuenn-Der Y, Hon-Cheng C. Effect of temperature, stocking density and fish size on the ammonia excretion in palmetto bass (Morone saxatilis_M. chrysops). Aquacult Res 2009; 40:450-455.
[ Links ]15. García-Gallego M, Suárez MD, Hidalgo MC, Sanz A, De la Higuera M. Nitrogen excretion by the eels: effect of the amount and source of dietary energy. Archiv Zoot 1999; 48:21-32.
[ Links ]16. Green BW, Boyd CE. Chemical budgets for organically fertilized ponds in the dry tropics. J World Aquacult Soc 1995; 26:284-296.
[ Links ]17. Hargreaves JA. A simulation model of ammonia dynamics in commercial catfish ponds in the southeastern United States. Aquacult Engineer 1997; 16:27-43.
[ Links ]18. Jimenez-Montealegre R, Avnimelech Y, Verreth J, Verdegem MCJ. Nitrogen budget and fl uxes in Colossoma macropomum ponds. Aquacult Res 2005; 36:8-15.
[ Links ]19. Josupeit H. Informe sobre Mercado de tilapia. 2009. FAO GLOBEFISH
[ Links ]20. Kaushik SJ, Cowey CB. Dietary factors affecting nitrogen excretion by fish. In: Cowey CB, Cho CY, editors. Nutritional Strategies and Aquaculture Waste. Proceeding of the first International symposium on nutritional strategies in management of aquaculture wastes. Canadá: University of Guelph; 1991. p.3-19.
[ Links ]21. Kelly AM, Kohler CC. Effects of Yucca shidigera extract on growth, nitrogen retention, ammonia excretion, and toxicity in channel catfish Ictalurus punctatus and hybrid tilapia Oreochromis mossambicus X O. niloticus. Journal of the World Aquacult Soc 2003; 34:156-161.
[ Links ]22. Leung KM, Chu JCV, Wu RSS. Nitrogen budgets for the areolated grouper Epinephelus areolatus cultured under laboratory conditions and open-sea cages. Marine Ecol Progr series 1999; 186: 271-281.
[ Links ]23. Martínez LR. Efecto de la dieta y otros factores sobre la excreción de amonio y el aprovechamiento del nitrógeno por la dorada Sparus aurata y su incidencia en los cultivos de esta especie. Tesis Doctorado 2002. (20/08/2009) .URL: http://www. tdx.cat/TDX-0731102-160857.
[ Links ]24. Perera WMK, Carter CG, Houlihan DF. Feed consumption, growth and growth efficiency of rainbow trout, Oncorhynchus mykiss Walbaum fed diets containing bacterial single cell protein. Brit J Nutr 1995; 73:591-603.
[ Links ]25. Person-Le R, Mahe JK, Le Bayon N, Le Delliou H. Effects of temperature on growth and metabolism in a Mediterranean population of European sea bass, Dicentrarchus labrax. Aquaculture 2004; 237:269-280.
[ Links ]26. Post JR, Lee JA. Metabolic ontogeny of teleost fishes. Canadian J Fisher Aquatic Sci 1996; 53:910-923.
[ Links ]27. Shuenn-Der Y, Chyng-Hwa,L, Fu-Guang L. Effects of dietary protein level on growth performance, carcass composition and ammonia excretion in juvenile silver perch (Bidyanus bidyanus). Aquaculture 2002; 213:363-372.
[ Links ]28. Sumagaysay-Chavoso. Nitrogen and phosphorous digestibility and excretion of different-sized of groups milkfish (Chanos chanos Forsskal) fed formulate and natural food-based diets. Aquacult Res 2003; 34: 407-418.
[ Links ]29. Vásquez-Torres W. Exigências de proteína, gordura e carboidratos em dietas para crescimento de juvenis de Pirapitinga, Piaractus brachypomus (CUVIER 1818). Tese de doutorado em Ciências Biológicas, área de concentraçãoBiologia de Água Doce e Pesca Interior. Convênio Universidade do Amazonas/Instituto Nacional de Pesquisas da Amazônia; Manaus (Brasil) 2001.p. 89.
[ Links ]30. Vásquez-Torres W. Las dietas como factor de impacto sobre la calidad del agua en sistemas de cultivo intensivo de peces. Rev Electr Ingen Prod Acuíc año II 2007; 1.
[ Links ]31. Vásquez-Torres W, Pereira-Filho M, Arias-Castellanos JA. Estudos para composição de uma dieta referência semipurificada para avaliação de exigências nutricionais em juvenis de pirapitinga, Piaractus brachypomus (Cuvier, 1818). Rev Brasil Zoot 2002; 31:283-292.
[ Links ]32. Webb KA, Gatlin DM. Effects of dietary protein level and form on production characteristics and ammonia excretion of red drum Sciaenops ocellatus. Aquaculture 2003; 225:17-26.
[ Links ]33. Zakes Z, Demska-Zakes K, Kata K. Rates of oxygen consumption and ammonia excretion of juvenile Eurasian perch Perca fluviatilis L. Aquacult Internat 2003; 11:277-288.
[ Links ]34. Zakes Z, Szczepkowski M, Demska-Sakes K, Jesiolowski M. Oxygen consumption and ammonia excretion by juvenile pike, Esox lucius l. Archiv Polish Fisher 2007; 15:79-92.
[ Links ]35. Zdzislaw-Zake S, Krystyna Demska-Zake S, Przemyslaw J, Konrad S. The effect of feeding on oxygen consumption and ammonia excretion of juvenile tench Tinca tinca (L.) reared in a water recirculating system. Aquacult Internat 2006; 14:127-140.
[ Links ]36. Zhang ZZ, Goodwin AE, Pfeijer TJ, Thomforde H. Effects of temperature and size on ammonia excretion by fasted golden shiners. North Am J Aquacult 2004; 66:15-19.
[ Links ]37. Zheng Z, Jin C, Li M, Bai P, Dong S. Effects of temperature and salinity on oxygen consumption and ammonia excretion of juvenile miiuy croaker, Miichthys miiuy (Basilewsky).Aquacult Internat 2008; 16:581-589.
[ Links ]38. Zimmermanns S, Fitzsimmons K. Tilapicultura Intensiva, In: Possebon-Cyryno JE Criusculo-Urbinati E, Machado-Fracalossi D, Castagnolli N Editores. Tópicos especiais in piscicultura de água doce tropical Intensiva. Sociedade Brasileira de Aqüicultura e Biologia Aquática. São Paulo: TecArt; 2004. p.239-266.
[ Links ]
