










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.24 no.4 Medellín Oct./Dec. 2011
Energy requirements for beef cattle: concepts and experimental results in tropical conditions¤
Exigências energéticas para gado de corte: conceitos e resultados experimentais em condições tropicais
Exigencias energéticas para ganado de carne: conceptos y resultados experimentales en condiciones tropicales
Sandra L Posada1, Zoot, PhD; Ricardo R Noguera1, Zoot, PhD; Norberto M Rodríguez2, Bioquim, PhD; Ana L Borges2, MV, PhD; Ricardo Reis2, Zoot, PhD.
1Grupo de Investigación en Ciencias Agrarias-GRICA, Facultad de Ciencias Agrarias, Universidad de Antioquia, AA 1226, Medellín, Colombia 2Departamento de Zootecnia, Escola de Veterinária da Universidade Federal de Minas Gerais, Brasil.
(Recibido: 5 mayo, 2010; aceptado: 16 agosto, 2011)
Summary
Life, interpreted as a combustion process, has been the basic principle guiding the study of energy metabolism, whether from calorimetric or slaughtering techniques. The relationships between metabolizable energy intake, retained energy, and heat production -together with the separation of maintenance and production functions- are the basis for the discussion of the factors affecting energy requirements and utilization efficiency in beef cattle. Energy metabolism is influenced by factors intrinsic and extrinsic to the animal, including age, weight, breed, body composition, sexual condition, production potential, environmental conditions, activity and feeding level, whose effects are discussed in this review. The understanding of these factors represents an opportunity to direct research programs seeking to determine the energy requirements under tropical conditions, in a similar way to the research conducted in developed countries.
Key words: efficiency, energy requirements, maintenance, production, zebu cattle.
Resumo
A vida como um processo de combustão tem sido o princípio que suporta o estudo do metabolismo energético, seja através de técnicas calorimétricas ou de abate comparativo. A relação entre consumo de energia metabolizável, energia retida e produção de calor, juntamente com a separação das funções de mantença e produção, constituem a base para a discussão dos fatores que afetam as exigências de energia e sua eficiência de utilização no gado de corte. O metabolismo energético esta influenciado por fatores intrínsecos e extrínsecos ao animal, entre eles, idade, peso, raça, composição corporal, condição sexual, potencial de produção, condições ambientais, atividade e nível de alimentação, cujos efeitos são discutidos nesta revisão. O conhecimento destes fatores representa uma oportunidade para encaminhar propostas de pesquisa dirigidas à determinação de exigências de energia em condições tropicais, a semelhança do que acontece nos países desenvolvidos.
Palavras chave: eficiência, energia, exigências, gado zebu, mantença, produção.
Resumen
La vida como proceso de combustión ha sido el principio que soporta el estudio del metabolismo energético, bien sea a partir de técnicas calorimétricas o de sacrificio animal. La relación entre consumo de energía metabolizable, energía retenida y producción de calor, conjuntamente con la separación de las funciones de mantenimiento y producción, constituyen la base para la discusión de los factores que afectan las exigencias de energía y su eficiencia de utilización en el ganado de carne. El metabolismo energético está influenciado por factores intrínsecos y extrínsecos al animal, entre ellos, edad, peso, raza, composición corporal, condición sexual, potencial de producción, condiciones ambientales, actividad y nivel de alimentación, cuyos efectos son discutidos en esta revisión. El conocimiento de estos factores representa una oportunidad para encaminar propuestas de investigación dirigidas a la determinación de las exigencias de energía en condiciones tropicales, a ejemplo de lo que sucede en países desarrollados.
Palabras clave: eficiencia, energía, exigencias, ganado cebú, mantenimiento, producción.
¤ To cite this article: Posada SL, R Noguera R, Rodríguez NM, Borges AL, Reis R. Exigências energéticas para gado de corte: conceitos e resultados experimentais em condições tropicais. Rev Colomb Cienc Pecu 2011; 24:623-633
* Corresponding author: Sandra L Posada. Universidad de Antioquia, Facultad de Ciencias Agrarias, Medellín (Colombia), Cra 75 #65-87. E-mail: slposada@gmail.com
Introdução
A energia é o componente que mais limita a produtividade animal, e conjuntamente com a proteína, representam as frações de maior participação no custo de alimentação. Quando a produção de gado de corte se desenvolve em pastejo, os animais alternam períodos de ganho e perda de peso em função da disponibilidade forrageira, consequentemente a melhora das condições nutricionais via suplementação é prioritária para incrementar o desempenho animal. Para a condução deste processo é fundamental o conhecimento das exigências nutricionais, que por serem pouco estudadas em nível latino- americano, faz com que as normas internacionais de alimentação continuem sendo a base para a formulação de rações. O Brasil se destaca como o país que tem conduzido mais pesquisas sobre o tema desde os anos oitenta, fato que deu lugar à elaboração da primeira tabela de exigênciasnutricionais de zebuínos por parte de Valadares et al. (2006), mesmo assim, a geração de informação e o volume de publicações deve aumentar a fim de possuir normas nutricionais mais consolidadas a nível tropical. Esta revisão de literatura tem como objetivo descrever alguns elementos básicos sobre energia (partição, estudo, eficiência de utilização) e apresentar algumas considerações sobre os fatores que afetam as exigências energéticas, mostrando resultados de pesquisa obtidos em condições tropicais.
Energia
A energia não é considerada nutriente, ela é liberada do alimento uma vez que todos os constituintes orgânicos (carboidratos, proteínas e lipídeos) são susceptíveis à oxidação (Resende et al., 2006). Com base nas pesquisas e conceitos elaborados por cientistas como Leonardo da Vinci (1452-1519), Joseph Priestly (1733-1804) e Antonie-Laurent Lavoisier (1743-1794), firmou-se o conceito de que a vida é um processo de combustão constante (Johnson et al., 2003).
Partição da energia no sistema animal
A energia bruta (EB) corresponde à energia liberada em forma de calor quando uma sustância orgânica é completamente oxidada a dióxido de carbono (CO2) e água. Uma parte da energia consumida (EC) se perde na forma de energia fecal (EF) e a diferença (EB-EF) recebe a denominação de energia digestível (ED). Outra parte da EC vai se perder como energia urinaria (EU) e energia gasosa (EG), resultando a energia metabolizável (EM). A EM pode se recuperar em forma de um produto útil (energia retida, ER), ou pode se perder em forma de calor (PC), o qual é conseqüência de diversas funções que incluem mantença e incremento calórico (IC) (Figura 1). Um aspecto importante da EM é seu valor como unidade de referência para os sistemas baseados no conceito de energia liquida (EL). Assim, a equação EM=ER+PC pode ser escrita como EM = ELr + ELm + IC, onde ELr e ELm são, respectivamente, a energia liquida retida e de mantença (NRC, 1996).
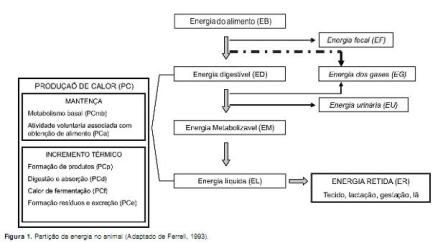
Técnicas para o estudo do metabolismo energético
Calorimetria. Na calorimetria direta, o calor desprendido pelo animal é medido pelo aumento da temperatura de um volume conhecido de água ou por meio de corrente elétrica. Já na calorimetria indireta, que pode ser de circuito aberto ou fechado, o calor é obtido através dos quocientes respiratórios (quantidade de gás carbônico produzido: quantidade de oxigênio consumida), que para carboidratos, proteínas e gordura são 1.0; 0.8 e 0.7, respectivamente (Resende et al., 2006; Rodriguez et al., 2007). Segundo o NRC (1996), a limitação da calorimetria radica em ajustar os dados obtidos em condições de laboratório a situações práticas de alimentação, não obstante, foi a metodologia base para o estabelecimento das exigências energéticas de sistemas como Agricultural Research Council (ARC, 1980), Commonwealth Scientific and Industrial Research Organization (CSIRO, 1990), Agricultural and Food Reasearch Council (AFRC, 1993) e National Research Council (NRC, 1989).
Abate comparativo. Na calorimetria, a ER é determinada por diferença entre o consumo de EM e a produção de calor. Uma técnica também muito utilizada é o abate comparativo, em que se quantifica diretamente a ER, e por diferença obtémse a produção de calor. Em 1959, Garret e seus colaboradores popularizaram o uso desta técnica e, a partir de então, ela foi utilizada nos trabalhos que constituíram a base das recomendações do National Research Council para bovinos de corte (NRC, 1984, 1996). Os procedimentos relacionados com o abate e o processamento matemático da informação são bem descritos por Veloso et al. (2002).
Considerando que o conteúdo de energia corporal determinado por abate comparativo precisa da análise química do organismo, esta técnica tem sido considerada custosa, laboriosa e destrutiva (uma vez que o animal tem que ser sacrificado), mas uma vantagem de seu emprego sobre as técnicas calorimétricas é permitir que as pesquisas sejam realizadas em condições mais parecidas com as de propriedades rurais (NRC, 1996). Numerosas comparações entre a técnica do abate comparativo e da calorimetria apontaram que a estimativa de produção de calor é maior pelo abate comparativo e, portanto, ER é menor (Resende et al., 2006).
Exigências energéticas
Em 1963, Lofgreen e Garrett introduziram o sistema de EL de gado de corte. O sistema separa as exigências de EL para mantença (ELm) daquelas para ganho de peso (ELg) e a soma de ambas representa as exigências líquidas totais de energia dos animais (Lofgreen e Garrett, 1968).
Exigências energéticas para mantença
Definição e importância. A exigência energética de mantença se define como a quantidade de energia que permite lograr um balanço energético zero e que, portanto, não determina ganho nem perda de peso do animal adulto (Ortigues et al., 1993). A manutenção do corpo tem grande importância sobre a economia total do animal; segundo Ferrell e Jenkins (1985), 65 a 70% da energia total necessária para produção de carne é utilizada para suprir a exigência de mantença.
Elementos constitutivos. As funções que compõem as necessidades de mantença do animal podem ser de dois tipos: serviço e associadas com a mantença celular. As funções de serviço são aquelas realizadas por tecidos ou órgãos para beneficio de todo o organismo, representam aproximadamente o 35-50% da produção de calor de metabolismo basal (PCmb) e incluem os trabalhos da circulação e da respiração, de fígado e rins (desintoxicação, mantença da osmolaridade e pH corporal) e as funções nervosas. Os componentes da mantença celular se denominam ciclos substrato, podem representar até o 30-50% de PCmb e abrangem transporte de íons (Na e Ca), reciclagem de proteínas e de lipídeos. Diferenças nos ciclos substrato determinam grande parte da variação nas exigências para mantença. As taxas de reciclagem de proteína são superiores em animais jovens e respondem a baixos níveis de alimentação, enquanto que a reciclagem de triglicerídeos aumenta em resposta a elevados níveis de alimentação. O transporte de íons é maior em animais jovens em relação aos adultos. Processos como manutenção da temperatura corporal e atividade física relacionada com a alimentação também contribuem aos gastos de energia para mantença (Ferrell, 1993).
Fatores que afetam as exigências energéticas de mantença
Tamanho metabólico. A velocidade metabólica (produção de calor por unidade de tempo) em animais de diferentes tamanhos é proporcional à superfície corporal. A área de superfície de dois corpos de forma e densidade similares é proporcional a ¾ de seu peso, valor correspondente à unidade de tamanho metabólico (UTM) (Kleiber, 1972). Isto se traduz em que a produção de calor por unidade de peso vivo diminui com a idade, razão pela qual os atuais sistemas de alimentação estimam o gasto energético em função do peso metabólico. Lofgreen e Garrett (1968) utilizando bovinos de corte, machos castrados e novilhas de raças britânicas, estimaram as exigências líquidas de mantença em 77 kcal/kg de peso corporal vazio metabólico (PCV0,75)/dia, referente à produção de calor em jejum (PCmb). Esse valor foi adotado pelo NRC (1984, 1996) e refere-se a animais confinados, sem estresse e com atividade mínima.
Proporções de órgãos. Mudanças nas exigências de mantença podem ser explicadas por variações nas proporções de órgãos do corpo. Mesmo as massas conjuntas de tecido nervoso, coração, rins, aparelho digestivo e fígado representem menos de 10% da massa corporal, recebem aproximadamente 55% do rendimento cardíaco e gastam o 50-60% de PCmb (Ferrell, 1993). Smith e Baldwin (1974) citados por Berndt et al. (2002), além dos órgãos mencionados, incluem à glândula mamaria como um tecido de grande atividade metabólica nos animais.
Os pesos do fígado, aparelho digestivo e, em menor quantia, dos rins e do coração, variam em resposta ao nível nutritivo e à idade (Ferrell e Jenkins, 1985). Segundo Almeida et al. (2001), quando é fornecido nível normal de nutrição a animais anteriormente submetidos a estresse nutricional, espera-se menor exigência de energia de mantença, por certo período de tempo, pela redução da massa dos órgãos internos. Em relação com a idade, um animal que atinge a maturidade apresenta declínio na proporção dos órgãos viscerais, especialmente o fígado e o trato digestivo, resultando em uma redução na exigência de energia para mantença. Em decorrência da menor exigência de energia para mantença, proporcionalmente ao peso corporal, mais energia pode ser gasta para o crescimento da carcaça, particularmente a deposição de gordura (Putrino et al., 2006).
Proporções de tecidos.A proteína do corpo, especialmente em órgãos viscerais, é metabolicamente muito mais ativa que o tecido adiposo e pode responder por diferenças nas exigências de mantença por unidade de massa corporal entre diferentes estádios de desenvolvimento (Silva et al., 2002b). Garrett (1980) observou que diferenças na intensidade de reciclagem protéica são responsáveis pelas maiores exigências de energia para mantença em raças bovinas que atingem a maturidade com pesos mais elevados, em machos inteiros em relação a castrados e destes em relação às fêmeas.
Condição sexual. Os machos inteiros têm maior exigência para mantença que os castrados e as fêmeas, porém seus ganhos têm menor conteúdo energético. Segundo ARC (1980), CSIRO (1990) e NRC (1996), as exigências de energia líquida para mantença (ELm) de machos castrados e novilhas são semelhantes, enquanto as de touros são 15% maiores do que as anteriores.
Grupo genético. Segundo o NRC (1996), os animais zebuínos apresentam exigências de energia para mantença 10% menores ao valor de referencia de 77 kcal/kg PCV0.75/dia, correspondente aos taurinos, enquanto as raças mestiças têm exigências intermediárias, ou seja, 5% a mais em relação aos zebuínos. Isto é explicado pelo menor potencial genético para produção das raças zebuínas, conseqüência de sua adaptação a condições ambientais menos favoráveis (Solis et al., 1988; Siqueira et al., 2007). O sistema australiano (CSIRO, 1990) recomenda um desconto maior das exigências energéticas de mantença para zebuínos, de 20%, em relação aos taurinos.
Para animais de quatro raças zebuínas (Gir, Tabapuã, Guzerá e Nelore), Paulino et al. (1999a) obtiveram valor médio de ELm de 60.4 kcal/kg0.75/ dia. Por outro lado, Boin (1995) encontrou um valor de 69.8 kcal/kg PCV0.75/dia para zebuínos não castrados. Para animais mestiços ½ Marchigiana x Nelore e ½ Limousin x Nelore, Pires et al. (1993a) estimaram valores de 67.92 e 68.03 kcal/kg PCV0.75/ dia, respectivamente.
Outros trabalhos não têm constatado diferenças entre grupos genéticos para as exigências de ELm. Tedeschi et al. (2002), em estudo com bovinos Nelore, acharam uma exigência de 77 kcal/kg0.75/ dia. Freitas et al. (2006b) obtiveram um valor médio de 79 kcal /kg0.75/dia para grupos genéticos Nelore, F1 Nelore x Aberdeen Angus, F1 Nelore x Pardo Suíço e F1 Nelore x Simental. Ferrell e Jenkins (1998b), estimaram um valor 77.2 kcal/kg0.75/dia e não detectaram diferenças entre novilhos Bos taurus (Angus e Hereford) e Bos indicus (Boran, Brahman e Tuli).
As discrepâncias nas exigências de ELm entre grupos genéticos podem ser explicadas por diferenças no tamanho de seus órgãos internos, que são maiores nos taurinos o F1 europeu-zebu do que nos zebuínos (Jorge et al., 1999). Somando-se a isso, Fontes et al. (2005) argumentaram que a maior exigência de energia de mantença para os taurinos e mestiços pode ser devida ao maior conteúdo de proteína corporal (massa muscular), resultando em maior reciclagem de proteína. Finalmente, Perón et al. (1993) relataram que os zebuínos, em relação aos taurinos, apresentam depósito de gordura periférica mais pronunciado, em detrimento ao depósito de gordura interna, razão pela qual suas exigências de energia para mantença são inferiores.
Exigências energéticas para produção
Definição. A energia líquida necessária para o crescimento e ganho de peso dos animais (ELg) corresponde ao valor calórico ou energia bruta dos tecidos, que é uma função da proporção de gordura e proteína depositadas no corpo (Garrett et al., 1959). O valor calórico da gordura é de 9.4 kcal/g e o da proteína, 5.6 Kcal/g (Backes et al., 2002). Dessa maneira, o entendimento do metabolismo energético do crescimento pode ser avaliado de duas maneiras: a) considerando a energia retida no tecido como um conceito uniforme, definindo a eficiência desse processo como kg e, b) separando as principais entidades químicas envolvidas no crescimento, a energia retida na forma de proteína e de gordura, com uma eficiência de deposição Kptn e Kgord para proteína e gordura, respectivamente (Resende et al., 2006).
Fatores que afetam a composição corporal e as exigências de ELg
Idade e peso. Segundo McDonald et al. (1995), o peso do animal, que tem uma relação direta com a idade, é o principal determinante da composição corporal. Após o nascimento, os bovinos iniciam uma fase de crescimento que pode ser representada por uma curva do tipo sigmóide. No inicio da vida à puberdade, os ganhos de peso consistem principalmente de água, proteína e minerais necessários para o crescimento dos ossos e dos músculos, conseqüentemente a curva possui inclinação crescente; a partir deste estádio, o crescimento muscular se reduz e por tanto a curva apresenta uma fase de desaceleração, caracterizada pelo aumento na taxa de deposição de gordura e diminuição na taxa de deposição de proteína. Assim, com a elevação do peso do animal, os conteúdos corporais totais de proteína e gordura no corpo elevam-se, os teores de proteína e água diminuem e os teores de gordura e energia aumentam (Freitas et al., 2000).
Freitas et al. (2006b) utilizando machos não- castrados verificaram elevação de 67.8% na concentração de gordura (g/kg PCV), com elevação de 250 para 550 kg no peso vivo, ao passo que a concentração de proteína corporal foi reduzida em 10.9%. Na sua pesquisa, as curvas de deposição de proteína e gordura foram próximas até 300 kg de PCV e seu distanciamento se relacionou com o final da fase de puberdade e início da maturidade do animal. A maior intensidade de crescimento muscular, em relação ao tecido adiposo, antes da maturidade depende da ação dos hormônios do crescimento (somatotropina), juntamente com os hormônios testosterona e tiroxina. De acordo com Owens et al. (1995), em bovinos em crescimento, o ganho de proteína declina a zero quando os animais atingem uma porcentagem de gordura no corpo vazio de aproximadamente 36%. O comportamento na deposição de gordura e de proteína com o avanço da idade é acompanhado pelo aumento nas exigências de energia e redução nas necessidades de proteína por kg de ganho (Carvalho et al., 2003).
Maturidade fisiológica. O verdadeiro determinante da composição dos ganhos é o peso relativo ao peso à maturidade do grupo genético do qual o animal procede, e não o peso corporal absoluto (Ferreira et al., 1999). Se comparados animais de maturidade fisiológica precoce em relação aos de maturidade tardia, observam-se maiores conteúdos corporais de gordura e menores de proteína nos primeiros. Considerando que o estádio de maturidade fisiológica é importante nos estudos de composição química corporal, as exigências energéticas devem ser corrigidas pelo fator de maturidade fisiológica. Para isto deve ser levado em consideração o peso standard de referencia (PER) (peso vivo de total desenvolvimento esquelético), que segundo NRC (1996) e CSIRO (1990) se obtém quando a porcentagem de gordura no PCV atinge 28 e 25%, respectivamente.
Condição sexual. As principais diferenças do sexo sobre a composição dos ganhos são observadas quanto ao tecido adiposo (Ferreira et al., 1999). Considerando animais pertencentes à mesma raça e com PCV similar, fêmeas possuem maior quantidade corporal de gordura que machos castrados, e estes, mais que os inteiros. A menor deposição de gordura no corpo, seguida por maior concentração protéica do ganho de peso em animais inteiros, é explicada pela secreção dos hormônios androgênicos, substâncias esteróides com ação anabólica (Guiroy et al., 2002). Segundo o NRC (1996), a um mesmo peso corporal, fêmeas e animais inteiros têm exigências energéticas líquidas para ganho de peso 18% superiores e inferiores às de novilhos, respectivamente.
Nível de alimentação. O nível de consumo de energia pode modificar a partição no uso da energia para síntese de proteína ou lipídeos (Backes et al., 2002). Dietas mais energéticas resultam em carcaças com maior teor de gordura (Jones et al., 1985). Em pesquisa conduzida por Ferrell e Jenkins (1998b), novilhos da raça Angus submetidos à alimentação restrita apresentaram menores teores de gordura na carcaça (22%) que aqueles sob alimentação ad libitum (30.7%). Esses autores verificaram que, para os mesmos níveis de ingestão alimentar, os teores de proteína na carcaça reduziram de 13.6 para 13%.
Taxa de ganho de peso. Maiores conteúdos de gordura no ganho se obtêm conforme aumenta o ganho de peso. Freitas et al. (2006b) trabalhando com bovinos Nelore puros e mestiços, assim como Berndt et al. (2002) com animais Santa Gertrudis, mostraram aumento quadrático na energia retida com o aumento do ganho de peso. Vários autores (Ferrell e Jenkins, 1998a,b; Owens et al., 1995) relataram que as taxas de acréscimo de proteína e de energia são maximizadas a uma determinada taxa de ganho de PCV, a partir da qual se mantêm constantes, com pouca ou nenhuma deposição adicional sob taxas de crescimento mais rápidas.
Grupo genético. O grupo genético apresenta, de acordo com Garrett (1980), maior influência sobre a composição corporal que o nível nutricional. Backes et al. (2005) trabalhando com animais mestiços leiteiros (½ sangue Holandês x Gir e ½ sangue Holandês x Guzerá) e zebu notaram que estes últimos apresentaram maior conteúdo de gordura por kg de ganho que os mestiços. Infere- se então que animais zebuínos possuem maturidade fisiológica mais precoce.
Eficiência de utilização da energia metabolizável (k)
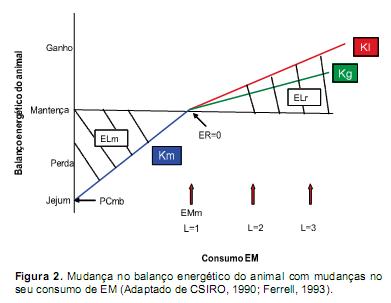
A eficiência de utilização da EM (k) é definida como o aumento na retenção de energia por unidade de aumento da ingestão de EM. A EM é utilizada pelos tecidos com uma eficiência inferior a 1 como resultado da produção de calor. A eficiência para mantença corresponde a km = ELm/EMm, em quanto a eficiência para produção pode ser expressa como kp = ELr/EM-EMm (Resende et al., 2006). Na figura 2 as linhas sólidas (pendentes da regressão linear) ilustram as eficiências para as diversas funções. Mostra-se que a EM é utilizada com maior eficiência para mantença (km), e com menor eficiência para crescimento (kg) e lactação (kl). Na mesma figura, as necessidades de EM para mantença (EMm) se apresentam como o consumo de EM no qual a ER=0. Neste ponto, a EMm é equivalente à produção de calor durante o jejum (PCmb) mais o IC, ou seja EM=PC (CSIRO, 1990).
O conhecimento da eficiência de utilização da energia metabolizável (k) da dieta para os diferentes processos biológicos é necessário para a determinação das exigências de energia metabolizável (a partir das exigências líquidas) e até mesmo de nutrientes digestíveis totais (NDT), o que tem maior valor prático, uma vez que a maioria das tabelas tropicais de composição química de alimentos fornece o valor energético dos alimentos em termos de NDT (Silva et al., 2002a).
O NRC (1984) apresentou valores de km entre 57.6 e 68.6% e, para kg de 29 até 47.3%, para dietas com diferentes proporções volumoso : concentrado, cujos teores de EM variaram de 2.0 a 3.2 Mcal/kg MS. O ARC (1980) desenvolveu equações lineares para o cálculo de k a partir da metabolizabilidade (relação EM/EB) da dieta, a qual não é constante, mas diminui conforme o nível de alimentação aumenta.
Fatores que afetam k
Nutrientes disponíveis. Os nutrientes não se substituem uns com outros de forma proporcional a seus calores de combustão, mas sim na medida em que proporcionam energia livre às células do organismo. Estabelecendo-se em 100% o valor relativo de eficiência da glicose, os valores correspondentes ao acetato, propionato, butirato, estearato e proteínas serão de 85, 87, 91, 95 e 76- 79%, respectivamente. Isto supõe que a EMm pode variar até em 20-25% dependendo dos nutrientes disponíveis para o animal (Baldwin et al., 1980).
O que diz respeito a kg, a grande variação nas estimativas de kptn e kgord advém, igualmente das diferenças na composição das dietas e, consequentemente, nos produtos finais da digestão (Garrett, 1980). kgord decresce com o aumento da proporção de ácido acético porque a síntese de ácidos graxos requer equivalentes reduzidos produzidos no ciclo das pentoses fosfatadas, sendo que a fermentação propiônica disponibilizara mais glicose para a ocorrência desta via metabólica (Véras et al., 2001a). Para a síntese de ácidos graxos tem-se observado eficiências de 30-60% quando se usa acetato, de 60-80% quando se usa glicose e de 90% ou superiores quando se emprega gordura dietética (Baldwin et al., 1980).
Proporção volumoso: concentrado. A adição de concentrado a dietas volumosas aumenta parcialmente km e kg em virtude da depressão na produção de metano, redução da ruminação e diminuição do incremento calórico (Véras et al., 2001a). Silva et al. (2002a) trabalhando com novilhos Nelore inteiros obtiveram estimativas de kg de 0.30; 0.35; 0.38 e 0.43, respectivamente, para as concentrações de EM da dieta de 2.65; 2.84; 2.92 e 2.90 Mcal/kg MS, correspondentes aos tratamentos 20, 40, 60 e 80% de concentrado. Não foi detectada diferencia em km, que foi 0.63.
Densidade energética e metabolizabilidade. Segundo ARC (1980), os valores de km e kg podem variar por causa da concentração de EM e metabolizabilidade da dieta, com kg apresentando maior variação do que km. Araújo et al. (1998) advertiram que constantes aumentos no consumo de energia diária podem resultar em pequenos aumentos na retenção de energia, toda vez que a energia da dieta decresce quando o nível de consumo de matéria seca aumenta. Esta relação inversa entre densidade energética e consumo de matéria seca se explica pela aceleração da taxa de passagem, que gera diminuição da digestão do amido e de carboidratos da parede celular e, portanto da metabolizabilidade (Véras et al., 2001a). Freitas et al. (2006a) indicaram que para o mesmo consumo de EM, dietas contendo maiores valores de metabolizabilidade proporcionam mais alta eficiência de utilização da EM para mantença e ganho que aquelas com menores valores.
Tecido depositado. A energia retida pelo animal no crescimento tecidual é menor do que o excesso de energia acima da mantença, porque existe um custo energético de crescimento. Essa diferença é devida á ineficiência acumulativa de todas as reações bioquímicas envolvidas no crescimento de um tecido, mais o IC do alimento adicional (Resende et al., 2006). Baldwin et al. (1980) citaram que a eficiência teórica máxima de crescimento dos ruminantes é de 70-80%; porém, os valores para kg são de 30-60%, existindo uma notável variação entre animais.
Diferentes percentuais de proteína e gordura no total de energia retida correspondem a diferentes eficiências de utilização da energia (Freitas et al., 2006a). A eficiência teórica da deposição de proteína, que poderia ser de 75-84%, pode se reduzir por conta da sua reciclagem (Baldwin et al., 1980). Garrett (1980) indicou que a eficiência liquida de deposição de proteína nos ruminantes flutua entre 10-40%. Quando a energia é estocada na forma de gordura, menor quantidade de calor é gerada, obtendo uma eficiência que oscila entre 60-80%. A eficiência aparentemente elevada da deposição de gordura pode ser resultado de taxas relativamente baixas de reciclagem e uma eficiência relativamente alta de síntese a partir dos nutrientes disponíveis (Ferrell, 1993). Considerando a curva normal de desenvolvimento de bovinos, Freitas et al. (2006a) afirmaram que durante a fase final de crescimento ocorre maior kg, fato associado à maior deposição de gordura no ganho com o acréscimo no PCV (maior grau de maturidade fisiológica).
Crescimento compensatório. Diversos experimentos demonstram que, quando o alimento volta a ser abundante após um período de restrição alimentar, as taxas de crescimento dos animais tornam-se mais aceleradas e excedem aquelas dos animais bem alimentados durante o mesmo período. Segundo Almeida et al. (2001), o ganho compensatório implica em redução das necessidades de ELm e incremento na utilização da energia metabolizável, usada acima da mantença, resultando em maior disponibilidade de ELg.
Consideração final
A eficiência produtiva e econômica dos sistemas de produção de gado de corte no trópico é altamente dependente de medidas racionais de manejo alimentar dos animais. Através das Universidades e Centros de Pesquisa deve se fomentar a construção de uma agenda de investigação que objetive a determinação das exigências nutricionais com a participação de raças zebuínas e autóctones, que além de possuir vantagens adaptativas ás condições ambientais imperantes, representam uma porcentagem importante do inventário bovino. Os resultados obtidos poderão ser adotados por profissionais e produtores do setor agropecuário para melhorar o balanceamento de dietas e o desempenho animal.
Agradecimientos
Los autores agradecen a la Fundación Universitaria San Martín por el respaldo económico para la divulgación de esta información y para el desarrollo del trabajo de investigación asociado con el tema presentado.
Referências
1. Agricultural Research Council - ARC. The Nutrient Requirements of Ruminant Livestock. Farnham Royal: Commonwealth Agricultural Bureaux; 1980. 351p. [ Links ]
2. Agricultural and Food Research Council - AFRC. Energy and Protein Requirements of Ruminants. Wallingford: CAB International; 1993. 159 p. [ Links ]
3. Almeida MI, Fontes CA, Almeida F, Valadares Filho SC, Campos OF. Conteúdo Corporal e Exigências Líquidas de Energia e Proteína de Novilhos Mestiços Holandês-Gir em Ganho Compensatório. Rev Bras Zootec 2001; 30:205-214. [ Links ]
4. Araújo GGL, Silva JFC, Valadares Filho SC, Signoretti RD, Turco SHN. Eficiência de utilização da energia metabolizável, para mantença e ganho de peso e exigências de energia metabolizável e nutrientes digestíveis totais de bezerros alimentados com dietas contendo diferentes níveis de volumoso. Rev Bras Zootec 1998; 27:1031-1036. [ Links ]
5. Backes AA, Sanchez LM, Gonçalves MB, Pires CC. Composição Corporal e Exigências Líquidas de Energia e Proteína para Ganho de Peso de Novilhos Santa Gertrudis. Rev Bras Zootec 2002; 31:2307-2313. [ Links ]
6. Backes A, Paulino MF, Alves DD, Rennó LN, Valadares Filho SC, Lana RP. Composição Corporal e Exigências Energéticas e Protéicas de Bovinos Mestiços Leiteiros e Zebu, Castrados, em Regime de Recria e Engorda. Rev Bras Zootec 2005; 34:257- 267. [ Links ]
7. Baldwin RL, Smith NE, Taylor J, Sharp M. Manipulating metabolic parameters to improve growth rate and milk secretion. J Anim Sci 1980; 51:1416-1428. [ Links ]
8. Berndt A, Henrique W, Leme PR, Alleoni GF, Lanna DPD. Exigências Líquidas de Proteína e Energia para Crescimento de Tourinhos Santa Gertrudis em Dietas de Alto Teor de Concentrado. Rev Bras Zootec 2002; 31:2098-2104. [ Links ]
9. Boin C. Alguns dados sobre exigências de energia e proteína de zebuínos. Simpósio internacional sobre exigências nutricionais de ruminantes. Viçosa, MG: Universidade Federal de Viçosa; 1995. p.457-465. [ Links ]
10. Carvalho PA, Bonnecarrère LM, Pires CC, Viégas J, Velho JP, Paris W. Composição Corporal e Exigências Líquidas de Proteína e Energia para Ganho de Peso de Bezerros Machos de Origem Leiteira do Nascimento aos 110 Dias de Idade. Rev Bras Zootec 2003; 32:1484-1491. [ Links ]
11. Ferrell CL. Metabolismo de la Energía. En: Church DC, editor. El Rumiante: Fisiología Digestiva y Nutrición. Zaragoza: ACRIBIA; 1993. p.283-303. [ Links ]
12. Commonwealth Scientific and Industrial Research Organization - CSIRO. Feeding Standards for Australian Livestock, Ruminants. Victoria: CSIRO Publications; 1990. 266 p. [ Links ]
13. Ferreira M, Valadares Filho SC, Silva JF, Paulino MF, Valadares R. Composição corporal e exigências líquidas de proteína e energia para ganho de peso de bovinos F1 Simental x Nelore. Rev Bras Zootec 1999; 28:352-360. [ Links ]
14. Ferrell CL. Jenkins TG. Body Composition and Energy Utilization by Steers of Diverse Genotypes Fed a High- Concentrate Diet During the Finishing Period: I. Angus, Belgian Blue, Hereford, and Piedmontese Sires. J Anim Sci 1998a; 76:637-646. [ Links ]
15. Ferrell CL, Jenkins TG. Body Composition and Energy Utilization by Steers of Diverse Genotypes Fed a High- Concentrate Diet During the Finishing Period: II. Angus, Boran, Brahman, Hereford, and Tuli Sires. J Anim Sci 1998b; 76:647- 657. [ Links ]
16. Ferrell CL, Jenkins TG. Cow type and the nutritional environment: nutritional aspects. J Anim Sci 1985; 61:725-741. [ Links ]
17. Fontes CA, Oliveira RC, Erbesdobler E, Queiroz D. Uso do Abate Comparativo na Determinação da Exigência de Energia de Mantença de Gado de Corte Pastejando Capim-Elefante: Descrição da Metodologia e dos Resultados. Rev Bras Zootec 2005; 34:1721-1729. [ Links ]
18. Freitas JÁ, Fontes CA, Soares JE, Jorge AM, Estrada LH. Composição corporal e exigências de energia para mantença de bovinos (zebuínos e mestiços) e bubalinos não castrados, em confinamento. Arq Ciên Vet Zool 2000; 3:19-29. [ Links ]
19. Freitas JA, Queiroz AC, Dutra AR, Vieira RAM, Lana RP, Leonel FP, Henrique DS, Lima AV, Souza JC. Eficiência de utilização da energia metabolizável em bovinos Nelore puros e cruzados submetidos a quatro níveis de concentrado na ração. Rev Bras Zootec 2006a; 35:894-901. [ Links ]
20. Freitas JA, Queiroz AC, Dutra AR, Vieira RAM, Lana RP, Leonel FP, Henrique DS, Lima AV, Souza JC. Composição corporal e exigências de energia de mantença em bovinos Nelore, puros mestiços, em confinamento. Rev Bras Zootec 2006b; 35:878-885. [ Links ]
21. Garrett WN, Meyer JH, Lofgreen GP. The comparative energy requirements of sheep and cattle for maintenance and gain. J Anim Sci 1959; 18:528-547. [ Links ]
22. Garrett WN. Factors influencing energetic efficiency of beef production. J Anim Sci 1980; 51:1434-1440. [ Links ]
23. Guiroy PJ, Tedeschi LO, Fox DG, Hutcheson JP. The effects of implant strategy on finished body weight of beef cattle. J Anim Sci 2002; 80:1791-1800. [ Links ]
24. Johnson DE, Ferrell CL, Jenkins TC. The history of energetic efficiency research: Where have we been and where are we going?. J Anim Sci 2003; 81 Suppl 1:E27-E38. [ Links ]
25. Jones SDM, Rompala RE, Jeremiah LE. Growth and composition of the empty body in steers of different maturity types fed concentrate or forage diets. J Anim Sci 1985; 60:427- 433. [ Links ]
26. Jorge AM, Fontes CA, Paulino MF, Júnior PG. Tamanho relativo dos órgãos internos de zebuínos sob alimentação restrita e ad libitum. Rev Bras Zootec 1999; 28:374-380. [ Links ]
27. Kleiber M. Bioenergética animal. El fuego de la vida. Zaragoza: ACRIBIA; 1972. 428 p. [ Links ]
28. Lofgreen GP, Garrett WN. A system for expressing net energy requirements and feed values for growing and finishing beef cattle. J Anim Sci 1968; 27:793-806. [ Links ]
29. McDonald P, Edwards RA, Greenhalgh JFD, Morgan CA. Animal Nutrition. 5 ed. Singapore: Longman; 1995. 607p. [ Links ]
30. National Research Council - NRC. Nutrient Requirements of Beef Cattle. 6 ed. Washington, D.C: National Academy Press; 1984. [ Links ]
31. National Research Council - NRC. Nutrient Requirements of Dairy Cattle. 6 ed. Washington, D.C: National Academy Press; 1989. [ Links ]
32. National Research Council - NRC. Nutrient requirements of beef cattle. 7 ed. Washington, D.C: National Academy Press; 1996. [ Links ]
33. Ortigues I, Petit M, Agabriel J, Vermorel M. Maintenance Requirements in Metabolizable Energy of Adult, Nonpregnant, Nonlactating Charolais Cows J Anim Sci 1993; 71:1947-1956. [ Links ]
34. Owens FN, Gill DR, Secrist DS, Coleman SW. Review of Some Aspects of Growth and Development of Feedlot Cattle. J Anim Sci 1995; 73:3152-3172. [ Links ]
35. Paulino MF, Fontes CA, Jorge A, Pereira JC, Júnior P. Exigências de energia para mantença de bovinos zebuínos não- castrados em confinamento. Rev Bras Zootec 1999a; 28:621- 626. [ Links ]
36. Perón AJ, Fontes CAA, Lana RP, Silva DJ, Queiroz AC, Paulino M. Tamanho de órgãos internos e distribuição da gordura corporal em novilhos de cinco grupos genéticos, submetidos a alimentação restrita e ad libitum. Revista da Sociedade Brasileira de Zootecnia 1993; 22:813-819. [ Links ]
37. Pires CC, Fontes CA, Galvão JG, Queiroz AC, Pereira J, Paulino MF. Exigências nutricionais de bovinos de corte em acabamento. I-Composição corporal e exigências de proteína para ganho de peso. Rev Bras Zootec 1993a; 22:110-120. [ Links ]
38. Putrino SM, Leme PR, Silva S, Alleoni GF, Lanna DPD, Grossklaus C. Exigências líquidas de proteína e energia para ganho de peso de novilhos Nelore alimentados com dietas contendo grão de milho úmido e gordura protegida. Rev Bras Zootec 2006; 35:301-308. [ Links ]
39. Resende KT, Teixeira IA, Fernandes MH. Metabolismo de energia. Em: Berchielli TT, Pires AV, Oliveira SG, editores. Nutrição de Ruminantes. São Paulo: FUNEP; 2006. 583 p. [ Links ]
40. Rodríguez NM, Campos WE, Lachica ML, Borges I, Gonçalves LC, Borges ALCC,, Saliba EOS. A calorimetry system for metabolism trials. Arq Bras Med Vet Zootec 2007; 59:495-500. [ Links ]
41. Silva F, Valadares Filho SC, Ítavo LC, Veloso CM, Paulino MF, Cecon PR, Moraes EBK, Paulino PVR. Eficiência de Utilização da Energia Metabolizável para Mantença e Ganho de Peso e Exigências de Energia Metabolizável e de Nutrientes Digestíveis Totais de Bovinos Nelore Não-Castrados. Rev Bras Zootec 2002a; Suppl 31:514-521. [ Links ]
42. Silva F, Valadares Filho SC, Ítavo LCV, Veloso CM, Valadares RFD, Cecon PR, Paulino PVR, Moraes EBK. Composição Corporal e Requisitos Energéticos e Protéicos de Bovinos Nelore, Não-Castrados, Alimentados com Rações Contendo Diferentes Níveis de Concentrado e Proteína. Rev Bras Zootec 2002b; Suppl 31:503-513. [ Links ]
43. Siqueira JG, Fontes CAA, Pereira AL, Lombardi CT, Sant´Ana NF, Vieira RAM. Exigência de energia de mantença e composição corporal e do ganho de vacas de corte adultas de três grupos genéticos confinadas. Rev Bras Zootec 2007; Suppl 36:2159-2167. [ Links ]
44. Solis JC, Byers FM, Schelling GT, Long CR, Greene LW. Maintenance requirements and energetic efficiency of cows of different breed types. J Anim Sci 1988;66:764-773. [ Links ]
45. Tedeschi LO, Boin C, Fox DG, Leme PR, Alleoni GF, Lanna DPD. Energy requirement for maintenance and growth of Nellore bulls and steers fed high-forage diets. J Anim Sci 2002; 80:1671-1682. [ Links ]
46. Valadares Filho SC, Paulino PVR, Magalhães KA. Exigências nutricionais de zebuínos e tabelas de composição de alimentos. 1 ed. Universidade Federal de Viçosa; 2006. 142 p. [ Links ]
47. Veloso CM, Valadares Filho SC, Júnior AG, Silva FF, Paulino MF, Valadares RFD, Cecon PR, Paulino PVR. Composição Corporal e Exigências Energéticas e Protéicas de Bovinos F1 Limousin x Nelore, Não-Castrados, Alimentados com Rações Contendo Diferentes Níveis de Concentrado. Rev Bras Zootec 2002; 31:1273-1285. [ Links ]
48. Véras AS, Valadares Filho SC, Silva JFC, Paulino MF, Cecon PR, Ferreira MA, Valadares RFD, Moraes EHBK. Eficiência de Utilização da Energia Metabolizável para Mantença e Ganho de Peso e Exigências de Energia Metabolizável e de Nutrientes Digestíveis Totais de Bovinos Nelore, Não-Castrados. Rev Bras Zootec 2001a; 30:904-910. [ Links ]

