










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.25 no.3 Medellín July/Sept. 2012
ARTÍCULOS ORIGINALES
Additive genetic group and heterosis effects on growth and corporal composition of crossbred cattle in southern Cesar (Colombia)¤
Efectos aditivos de grupo genético y heterosis sobre crecimiento y composición corporal en bovinos cruzados en el sur del Cesar (Colombia)
Efeitos aditivos do grupo genético e de heterose para crescimento e composição corporal de bovinos mestiços no sul do Cesar (Colômbia)
Carlos A Martínez Niño1,2*, Zoot, MSc; Carlos Manrique Perdomo1, Zoot, MSc, PhD; Mauricio A Elzo2, MV, PhD; Ariel Jiménez Rodríguez1,3, MV, MSc.
* Autor para correspondencia: Carlos A Martínez Niño. Departament of Animal Sciences, Gainesville, FL 32611-0910, USA. Telephone (352) 3927564, e-mail: carlosmn@ufl.edu
1Grupo de Estudio en Mejoramiento y Modelación Animal GEMA, Departamento de Producción Animal, Universidad Nacional de Colombia, Bogotá, Colombia
2Department of Animal Sciences, University of Florida, FL 32611-0910, USA.
3Asociación Colombiana de Criadores de Ganado Cebú ASOCEBU, Bogotá, Colombia
(Recibido: 26 junio, 2011; aceptado: 22 febrero, 2012)
Summary
Objective: given the need to evaluate several cattle breeds for growth and body composition traits in Colombia, the objective of this study was to obtain the best linear unbiased estimators of heterosis and breed group effects for these traits. Methods: live weight was evaluated at birth (PN), and aproximately at 4 months (P4), weaning (PD), first year (PA), and 15 months of age (P15). Body composition traits evaluated were rib eye area (AOL) and back fat thickness (GD) measured at the same ages as live weight, except at birth: AOL4 and GD4, AOLD and GDD, AOLA and GDA, AOL15 and GD15. The evaluated population consisted of 352 offspring from 37 bulls representing 9 breeds (Gray Brahman, Red Brahman, Guzerat, Blanco Orejinegro, Romosinuano, Normando, Braunvieh, Limousin, and Simmental), and Gray Brahman females. The farms were located in southern Cesar, Colombia. Univariate animal models were used. Models included contemporary group, dam parity, age of the animal, expected fraction of breed group in the animal, and proportion of heterozygosis as fixed effects, with direct additive and maternal as random effects. Results: the highest additive genetic values for PN, P4, PD, PA, P15 were found in Guzerat (6.31 kg), Normando (12.67 kg), Normando (8.28 kg), Braunvieh (24.80 kg) and Normando (42.20 kg), respectively. Those for AOL4, AOLD, AOLA and AOL15 in Limousin (20.87 cm2), Limousin (12.60 cm2), Simmental (10.94 cm2) and Limousin (16.09 cm2). And those for GD4, GDD, GDA and GD15 in Limousin (0.27 mm), Guzerat (0.21 mm), Blanco Orejinegro (1.05 mm) and Romosinuano (0.71 mm), respectively. The heterosis values for live weight ranged from -3.22 (PA) to 7.43 kg (P15), for AOL from 0.12 (AOL4) to 4.39 mm2 (AOLA) and for GD from 0.01 (GDD and GD15) to 0.09 mm (GDA). Conclusion: results indicated that Normando was generally superior for live weight and Limousin was superior for AOL among the animals sampled from each breed in the study.
Key words: beef cattle, corporal composition, crossbreeding, growth, heterosis.
Resumen
Objetivo: dada la necesidad de evaluar varias razas bovinas en Colombia para caracteres de crecimiento y composición corporal, el objetivo de este estudio fue obtener mejores estimadores lineales insesgados para efectos de heterosis y grupo racial para tales caracteres. Métodos: las características de crecimiento evaluadas fueron el peso a diferentes edades: nacimiento (PN), y aproximadamente a los 4 meses (P4), destete (PD), año (PA), y 15 meses (P15). Las caracteristicas de composición corporal fueron área de ojo del lomo (AOL) y grasa dorsal (GD) medidas a las mismas edades que el peso, excepto al nacimiento: AOL4 y GD4, AOLD y GDD, AOLA y GDA, AOL15 y GD15. La población estuvo conformada por 352 progenies de 37 toros representando 9 razas (Brahman Gris, Brahman Rojo, Guzerat, Blanco Orejinegro, Romosinuano, Normando, Braunvieh, Limousin y Simmental) y hembras Brahman Gris. Se emplearon modelos animales uni-varidos con los efectos de: grupo contemporáneo, número de partos de la madre, edad del animal, fracción esperada de grupo racial en el animal, y proporción de heterocigosis como efectos fijos y aditivos directo y materno como aleatorios. Resultados: los mayores valores genéticos aditivos para PN, P4, PD, PA y P15 se encontraron en animales de la raza Guzerat (6.31 kg), Normando (12.67 kg), Normando (8.28 kg), Braunvieh (24.80 kg) y Normando (42.20 kg), respectivamente. Para AOL4, AOLD, AOLA y AOL15 se encontraron en animales de la raza Limousin (20.87 cm2), Limousin (12.60 cm2), Simmental (10.94 cm2) y Limousin (16.09 cm2), respectivamente. Y para GD4, GDD, GDA y GD15 se encontraron en animales de la raza Limousin (0.27 mm), Guzerat (0.21 mm), Blanco Orejinegro (1.05 mm) y Romosinuano (0.71 mm). Los valores de heterosis para peso variaron entre -3.22 kg (PA) y 7.43 kg (P15), para AOL entre 0.12 cm2 (AOL4) y 4.39 cm2 (AOLA) y para GD entre 0.01 (GDD y GD15) y 0.09 mm (GDA). Conclusiones: los resultados indicaron que en general Normando fue superior para peso vivo, y Limousin para AOL entre los animales de cada raza en el estudio.
Palabras clave: composición corporal, crecimiento, cruzamiento, ganado de carne, heterosis.
Resumo
Objetivo: em virtude da necessidade de avaliar diversas raças de bovinos na Colômbia para características de crescimento e composição corporal, o objetivo deste estudo foi determinar o melhor estimador linear não-viciado para obter os efeitos de heterose e grupo racial para essas características. Métodos: as características de crescimento foram os pesos a diferentes idades: nascimento (PN), y aproximadamente aos quatro meses (P4), desmame (PD), ano (PA) e 15 meses (P15). Características de composição corporal como a área de olho de lombo (AOL) e espessura de gordura subcutânea (GD) foram mensuradas nas mesmas idades, exceto ao peso ao nascimento (AOL4 e GD4; AOLD e GDD; AOLA e GDA; AOL15 e GD15). A população estava conformada por 352 progênies de 37 touros pertencentes a nove raças (Brahman Cinza, Brahman Vermelho, Guzerá, Blanco Orejinegro, Romosinuano, Normando, Braunvieh, Limuosin e Simental) acasalados com fêmeas da raça Brahman Cinza. Foram empregados modelos animais univariados considerando: efeito de grupo de contemporâneos, número de partos da mãe, idade do animal, fração esperada do grupo racial no animal e proporção da heterozigose como efeitos fixos, e como aleatórios o efeito aditivo direto e o efeito materno. Resultados: os maiores valores genéticos aditivos para PN, P4, PD, PA e P15 foram encontrados em animais da raça Guzerá (6.31 kg), Normando (12.67 kg), Normando (8.28 kg), Braunvieh (24.80 kg) e Normando (42.20 kg) respectivamente. Para AOL4, AOLD, AOLA e AOL15 os maiores valores foram encontrados em animais da raça Limousin (20.87 cm2), Limousin (12.60 cm2), Simental (10.94 cm2) e Limousin (16.09 cm2), respectivamente. E para GD4, GDD, GDA e GD15 os maiores valores foram encontrados na raça Limousin (0.27 mm), Guzerá (0.21 mm), Blanco Orejinegro (1.05 mm) e Romosinuano (0.71 mm). Os valores de heterose para peso variaram de -3.22 kg (PA) ate 7.43 kg (P15), para AOL entre 0.12 cm2 (AOL4) e 4.39 cm2 (AOLA) e para GD entre 0.01 mm (GDD e GD15) e 0.09 mm (GDA). Conclusões: os resultados indicam que em geral animais da raça Normando foram superiores para peso vivo quando se compararam com as outras raças, e para a AOL animais da raça Limousin apresentaram os maiores valores.
Palavras chave: composição corporal, crescimento, cruzamento, gado de corte, heterose.
Introducción
En Colombia los cruzamientos son muy comunes en las zonas de trópico bajo (FEDEGAN, 2006), en donde las condiciones ambientales adversas presentan un reto para los animales que se desempeñan en tales ecosistemas. En el país se han empleado razas de diferentes orígenes para establecer programas de cruzamiento. En Colombia existen ocho razas bovinas criollas de la especie Bos taurus (FEDEGAN, 2006; Martínez, 2010); por otro lado, están las razas taurinas especializadas en producción de carne y las razas de la especie Bos indicus que constituyen el 72% de la población vacuna nacional con predominio del Brahman (Martínez, 2010).
La preferencia por un conjunto de razas para establecer un programa de cruzamientos debe obedecer a criterios objetivos. Por esta razón en la actual política de desarrollo del sector ganadero en el país, se identifican los cruzamientos como herramienta de mejora genética y se contempla como prioridad evaluar el desempeño de diferentes razas y sus cruzamientos para tomar decisiones acertadas sobre los recursos genéticos que se han de emplear en los programas de cruza (FEDEGAN, 2006). Han sido pocos los estudios realizados sobre animales mestizos en el país (Vergara et al., 2009); y la mayoría de estos tienen en cuenta pocas razas (Elzo et al., 1998a; Elzo et al., 2001). De allí nace la necesidad de conducir estudios que comparen el desempeño de varias razas en caracteres de importancia económica como el crecimiento y los de composición de canal, medidos vía ultrasonido que están relacionados con la producción de cortes de alto valor comercial (Wilson et al., 1992; Elzo et al., 1998b) y que resultan útiles en programas de selección (Bertrand, 2009). Se ha reportado que el cruzamiento ofrece ventajas como el incremento en el desempeño de los animales en caracteres de crecimiento y de canal (Williams et al., 2010). También se requiere estimar el efecto de heterosis, valor que junto con los efectos de raza sirve para orientar un programa de cruces.
Por lo tanto, el objetivo de la presente investigación fue obtener los mejores estimadores lineales insesgados de los efectos de grupo racial y de heterosis en una población compuesta por animales F1 producto del apareamiento de 37 toros de nueve razas con 352 vacas Brahman gris, en la microrregión del sur del Cesar en Colombia, la cual, por sus condiciones ambientales, está catalogada como estratégica para la producción de carne bovina.
Materiales y métodos
Apareamientos
Se programaron mediante un protocolo de Inseminación Artificial a Tiempo Fijo (IATF) apareamientos aleatorios de 37 toros de nueve razas con 352 vacas y novillas Brahman Gris. Las razas empleadas pertenecían a tres grupos según su origen: Taurinas europeas (TE), Taurinas Criollas (TC) y Cebuínas (C). Las razas del grupo TE fueron: Braunvieh (BV), Limousin (LIM), Normando (NM) y Simmental (SIME). Las razas del grupo TC fueron: Blanco Orejinegro (BON) y Romosinuano (RS). Finalmente, las razas del grupo C fueron: Brahman gris (BG), Brahman rojo (BR) y Guzerat (GZ). Las novillas se escogieron con base a su peso vivo (mayor de 350 kg) y estado general, mientras que las vacas se seleccionaron con base al número de partos (3) y a la sanidad y estado del sistema reproductivo. La IATF se llevó a cabo mediante la colocación de un implante de progesterona (CIDR, Pfizer, NY, USA) y 2 mg de Benzoato de estradiol el día 0. El día 8 se retiró el dispositivo y se aplicó 1 cm3 de prostaglandina F2α (Estrumate, Schering Plough S.A., Kenilworth, NJ, USA). 24 horas después se inyectó 1 mg de benzoato de estradiol. La inseminación artificial se realizó 54 horas después del retiro del implante de progesterona.
Animales y manejo
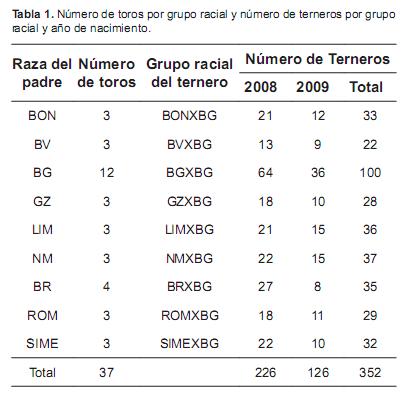
Las crías nacieron en los años 2008 y 2009. La tabla 1 muestra el número de toros por raza y el número de terneros nacidos por grupo racial y año de nacimiento. Los animales fueron criados en dos haciendas ubicadas en el municipio de Aguachica, Departamento del Cesar, Colombia. Esta región presenta una altura sobre el nivel del mar de 50 m, una temperatura media anual de 28 °C y una humedad relativa de 80%. Por estas características, según la clasificación de Holdridge, esta zona corresponde a un bosque muy seco tropical (bms-T), lo cual hace que sea una microrregión deseable para la producción de carne bovina en el país. Se manejó un sistema de pastoreo rotacional con suplementación mineral. Las praderas estaban compuestas por las siguientes especies forrajeras: Brachipará (Brachiaria plantaginea), Guinea (Panicum maximum) y Ángleton (Dichantium aristatum). El suplemento mineral consistió en una sal mineralizada ofertada a granel con un contenido de fósforo del 8% (GANASAL®, Colombia). Los terneros se destetaron a una edad aproximada de 8 meses. Los machos fueron castrados a los 12 meses.
Caracteres de crecimiento
Se consideraron cinco caracteres de crecimiento: peso al nacer (PN, kg) y 4 pesos tomados aproximadamente a los 4 meses (PA4, kg), 8 meses (destete; PD, kg), 12 meses (PA, kg) y 15 meses (P15, kg). Los pesajes fueron tomados y digitados por personal de la Asociación Colombiana de Criadores de Ganado Cebú (ASOCEBU, Colombia), a excepción del peso al nacer, el cual fue tomado por el personal de cada hacienda.
Caracteres de canal medidos vía ultrasonido
Se consideraron ocho caracteres de canal medidos mediante ultrasonido en tiempo real: 4 de área del músculo Longissimus dorsi (AOL) y 4 de espesor de grasa dorsal (GD) medidos entre la doceava y treceava costilla. Las cuatro mediciones de AOL y de GD se hicieron aproximadamente a los 4 (AOL4, cm2; GD4, mm), 7 (AOLD, cm2; GDD, mm), 12 (AOLA, cm2; GD12, mm) y 15 meses (AOL15, cm2; GD15, mm). Las medidas de ultrasonido fueron tomadas por un técnico entrenado de la Asociación Colombiana de Criadores de Ganado Cebú (ASOCEBU, Colombia) usando un equipo Aquila modelo Esaote de Pie Medical (Pie Medical Equipment B.V., Maastricht, Limburg, TheNetherlands) con sonda ASP 18 y almohadilla de acople. Se utilizó el programa Echo Image Viewer (Pie Medical Equipment B.V., Maastricht, Limburg, The Netherlands) para obtener las mediciones de área y longitud requeridas.
Registros
Los registros genealógicos de los terneros, así como el sexo, fecha de nacimiento y raza de los padres, fueron colectados por el personal de las fincas en el programa GANADERO® (USATI®, Bolívar, Colombia). Los pedigrís de los reproductores fueron suministrados por cada una de las asociaciones ganaderas participantes. Los pesajes y medidas de ultrasonido fueron colectados desde Octubre de 2008 hasta Mayo de 2010.
Análisis genéticos
Se realizaron análisis univariados mediante procedimientos de modelos mixtos con la finalidad de obtener los mejores estimadores lineales insesgados (MELI) de funciones estimables de los efectos fijos (Henderson, 1988). Para alcanzar tal objetivo se hizo necesario en un primer paso explorar la estimabilidad de los parámetros en diferentes escenarios, esto es, verificar que los diferentes efectos fijos o combinaciones lineales de los mismos que sean de interés se pudiesen estimar de manera insesgada.
Tales escenarios se definieron de acuerdo a la manera de definir los grupos raciales:
1. Individualmente, es decir, 9 grupos (criterio C1).
2. Brahman blanco y Brahman rojo en un solo grupo y los demás grupos de manera individual (criterio C2).
3. Cebuinos en un grupo y las demás razas de manera individual (criterio de agrupamiento C3).
Para cada uno de los agrupamientos anteriores el efecto aditivo directo del grupo racial se modeló mediante regresión, esto es, con la fracción esperada de cada grupo (según se defina en C1, C2, y C3) en el animal como regresor.
Un segundo criterio que se exploró fue la forma de modelar el efecto de heterosis directa:
1. Mediante la heterocigocidad directa individual (HI).
2. Fraccionando la heterocigocidad directa en heterocigocidad directa taurina-cebuina (HTC) y cebuina-cebuina (HCC). No hubo cruzamientos entre razas taurinas en esta población multirracial, por lo cual no se estimó heterosis taurina-taurina (HTT).
Este primer paso se hizo necesario, ya que se ha reportado que puede ser muy difícil conseguir estructuras de las bases de datos que permitan la estimación de todos los efectos aditivos y no aditivos directamente de la información colectada a nivel de campo (Bertrand, 2009). Se debe tener presente que en este estudio sólo se tuvieron animales F1 y Brahman puros, por lo cual existió dependencia lineal entre las columnas de las matrices de diseño de los efectos fijos. Bajo el criterio C1 la columna de heterosis tuvo unos en las filas correspondientes a animales F1 y ceros en las filas que representan animales BG. Las columnas de grupos raciales diferentes a Brahman Gris tuvieron un 0.5 en las filas correspondientes a animales F1 y ceros en las filas asociadas a animales puros. Por lo tanto, la suma de las columnas de los grupos raciales diferentes al Brahman multiplicada por dos, será igual a la columna de heterosis. Por lo anterior, se tuvo que fijar en cero una de las soluciones de los grupos raciales. Se eligió forzar a cero la solución de BG para C1, Brahman (Gris y Rojo) para C2 y Cebuinos para C3. Con estas restricciones sobre las soluciones de los modelos mixtos establecidos, se procedió a evaluar su estimabilidad bajo los escenarios planteados.
La HI se calculó mediante la siguiente expresión (Cerón-Muñoz, 2008):
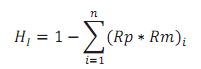
Donde n es el número de razas involucradas y Rp y Rm las fracciones esperadas de cada una en el padre y la madre respectivamente.
La HCC se calculó como sigue:
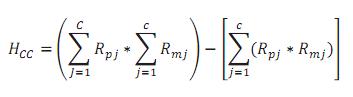
Siendo c el número de razas cebuinas involucradas, y Rpj y Rmj las fracciones esperadas de la j-ésima raza cebuina en el padre y la madre, respectivamente.
Finalmente para la HTC, la expresión empleada para su cálculo fue:
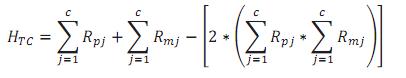
En donde cada uno de los miembros de la expresión es definido de la misma manera que lo está en el cálculo de HCC.
Para los caracteres pre-destete se empleó el siguiente modelo univariado:
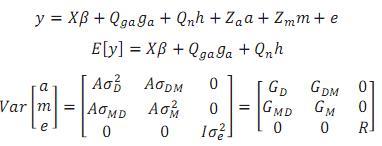
Donde:
y = Vector de registros.
β =Vector desconocido de efectos fijos de grupo contemporáneo (subclase año de nacimiento* hacienda*época de nacimiento*sexo), número de partos de la madre y coeficientes de regresión lineal y cuadrática de la edad del animal.
ga = Vector desconocido de efectos fijos aditivos directos de grupo racial.
h = Escalar desconocido de efectos no aditivos fijos, heterosis en un locus.
a = Vector aleatorio desconocido con los efectos genéticos aditivos directos de cada animal.
m = Vector aleatorio desconocido de efectos genéticos aditivos maternos.
e = Vector aleatorio de residuos.
X, Qga, Qn, Za y Zm son matrices de incidencia que respectivamente relacionan:
1. El vector de efectos no genéticos fijos al vector de registros a través de unos para las variables de clasificación y edad del animal y edad del animal al cuadrado para los efectos de la edad.
2. El vector de efectos aditivos directos fijos de grupo racial al vector de registros mediante la fracción esperada de cada grupo en el individuo.
3. El vector que relaciona el efecto de heterosis al vector de registros mediante la probabilidad de alelos de diferentes razas en un locus.
4. el vector de efectos aleatorios directos aditivos al vector de registros a través de unos.
5. el vector de efectos aleatorios maternos aditivos al vector de registros mediante unos.
A = matriz de parentescos aditivos,
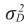 = varianza genética para efectos aditivos directos,
= varianza genética para efectos aditivos directos,
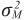 = varianza genética para efectos aditivos maternos,
= varianza genética para efectos aditivos maternos,
σDM= covarianza entre efectos aditivos directos y efectos aditivos maternos,
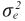 = varianza residual.
= varianza residual.
Para los caracteres pos-destete, se asumió que el efecto materno no era importante, por ello, para estos caracteres el modelo animal empleado tuvo los mismos efectos a excepción del efecto materno, esto es:
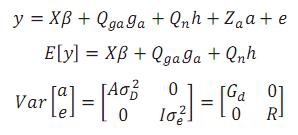
Los estimadores de máxima verosimilitud restringida (MVR) de los componentes de varianza y los MELI de los efectos fijos se obtuvieron mediante el programa MTDFREML (Boldman et al., 1995), el cual emplea un método libre de derivadas para encontrar el máximo de la función de verosimilitud.
Para realizar inferencias sobre los parámetros de modelos lineales mixtos, los grados de libertad debieron aproximarse (Littelet al., 2006). En este caso, los grados de libertad requeridos para realizar la prueba t sobre los efectos fijos de heterosis y de grupo racial (se probó si estos fueron diferentes de cero) se aproximaron mediante el método general de Satterthwaite, empleando el procedimiento MIXED de SAS (SAS, 2008), en el cual los grados de libertad (v) se calculan como sigue:
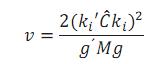
Donde ki= función estimable de los efectos fijos,
Ĉ = inversa generalizada de la matriz de coeficientes de la parte fija del modelo,
g = gradiente (vector de primeras derivadas parciales) de ki'Ĉki con respecto a θ (el vector de parámetros desconocidos en la matriz de covarianzas del vector de registros (V)) evaluado en su estimador MVR 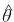 .
.
M = matriz de covarianzas asintóticas de , la cual se obtiene de la matriz de segundas derivadas de las ecuaciones de verosimilitud.
Para realizar esta estimación, se utilizaron los componentes de Covarianza obtenidos de MTDFREML. Así, los dos modelos fueron exactamente iguales. Para el caso del programa en SAS, la matriz de parentesco se construyó a través del procedimiento INBREED. Se utilizó el paquete SAS para la estimación de los grados de libertad requeridos para la prueba t, porque MTDFREML no tiene implementada una rutina para estimarlos.
Resultados
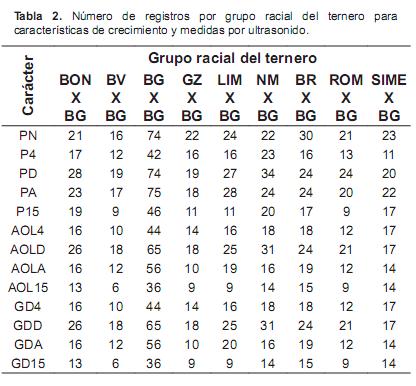
Como se aprecia en la tabla 2, el número de datos por grupo racial varió entre 6 (GD15 en el grupo BVXBG) y 75 (PA para el grupo BGXBG).El número total de datos para los caracteres estudiados varió entre 125 (AOL15 y GD15) y 269 (PD). El número total de datos, los valores medios, las desviaciones estándar y los coeficientes de variación para los 13 caracteres estudiados se muestran en la tabla 3. Los caracteres que presentaron una mayor variación fenotípica, a juzgar por el coeficiente de variación, fueron las mediciones de espesor de grasa dorsal.
Las edades medias para las variables P4, PD, P12 y P15 fueron: 120.2, 221.6, 346.7 y 447.1 d, respectivamente. La edad promedio para las variables AOL4 y GD4 fue 120.31 d; para AOLD y GDD fue 232.97 d; AOLA y GDA 331.81 d, y para AOL15 y GD15 fue 445.98 d. Estos promedios se calcularon con los datos que finalmente se utilizaron en los análisis, es por esto, que si bien los pesajes y las mediciones de ultrasonido se tomaron en las mismas ocasiones, las medias de edad difieren.
Estimabilidad
Bajo el criterio C1 se encontró que la heterosis no pudo ser estimada de manera insesgada, por lo tanto, se hizo necesario fijar la solución de dos grupos raciales a cero. La heterosis individual fue estimable bajo el criterio de definición de grupos raciales C2, caso en el cual, los efectos de los demás grupos se expresan como desvíos de los animales Brahman (grupo conformado por BGXBG y BRXBG). Al fraccionar HI como HCC y HTC, no fue posible estimar estos dos componentes de la heterosis de manera insesgada bajo los agrupamientos C2 ni C3. Estos efectos fueron estimables sólo cuando el criterio de agrupamiento fue: TC, TE y C, pero dicho agrupamiento se descartó pues no fue acorde con los objetivos del trabajo. Por lo tanto, el set de funciones estimables K'ga, las cuales describieron el efecto de los grupos raciales y para las cuales se obtuvieron los MELI fueron de la forma:
ki'ga = [0 0 ...1 0 ... 0 - 1] ga,i = 1, 2, 3, ..., 7
En donde ki' representa la i-ésima fila de la matriz K' o equivalentemente la i-ésima función estimable para el i-ésimo grupo racial (definido según el criterio C2), el escalar uno está en la i-ésima posición y el escalar -1 está en la posición asociada al grupo Brahman (posición 8).
Efectos de grupo racial
En la tabla 4 se presentan los MELI de los efectos de grupo racial junto con su error de estimación para cada uno de los caracteres estudiados. Los mayores valores aditivos para PN, P4, PD, PA y P15 los presentaron las razas GZ (6.31 kg), NM (12.67 kg), NM (8.28 kg), BV (24.80 kg) y NM (42.20 kg), respectivamente; para AOL4, AOLD, AOLA y AOL15: LIM (20.87 cm2), LIM (12.60 cm2), SIME (10.94 cm2) y LIM (16.09 cm2), respectivamente y para GD4, GDD, GDA y GD15: LIM (0.27 mm), GZ (0.21 mm), BON (1.05 mm) y RS (0.71 mm).
Efectos de heterosis
Los valores de heterosis para los caracteres de crecimiento variaron entre -3.22 (PA) y 7.43 kg (PD); para AOL el rango fue 0.12 (AOL4) a 4.39 cm2 (AOLA) y para GD de 0.01 (GDD; GD15) a 0.09 mm (GDA). El efecto de heterosis solamente fue significativo (p<0.05) para las variables PD, AOLA y AOL15 (Tabla 4). Como se esperaba, debido al reducido tamaño de la población y al número relativamente alto de grupos raciales, los errores de estimación fueron grandes para la mayoría de los parámetros.
Discusión
Estimabilidad
Elzo y Famula (1985), reportaron que puede existir confusión entre efectos genéticos fijos debido a subclases vacías (tipos de apareamientos no realizados), lo cual concuerda con lo encontrado en el presente estudio, ya que sólo se tenían apareamientos para obtener progenie F1 y animales BG puros. Tales resultados también fueron descritos por Rodríguez-Almeida et al. (1997), quienes reportaron que se requieren diferentes tipos de cruzamientos para poder separar los efectos de grupo racial.
En un estudio conducido por Sánchez et al. (2008) se hizo necesario incluir información previa, debido a que no existieron suficientes grupos contemporáneos con una composición adecuada de animales puros y cruzados para que los efectos de heterosis y aditivos de grupo racial se pudieran estimar.
En este estudio, se tuvo representación de todos los grupos raciales en todos los grupos contemporáneos, pero solamente un tipo de cruzamientos (p.e., F1). Esto creó confusión entre efectos raciales aditivos y de heterosis en cruzamientos específicos, lo cual impidió que estos dos tipos de efectos se pudieran estimar de forma insesgada en el agrupamiento racial C1. En estudios que utilizaron modelos similares, pero con más tipos de apareamientos por pares de razas y mayor número datos, fue posible estimar los efectos aditivos directos fijos de cada una de las razas consideradas como desviaciones del grupo Cebú (Arboleda et al., 2008; Vergara et al., 2009). En el estudio de Arboleda et al. (2008) fue posible estimar particiones de la heterosis (HTT y HTC) al igual que en la investigación conducida por Barreto (2005).
Efectos de grupo racial
No se encontraron reportes en la literatura de trabajos que involucraran la totalidad de las razas estudiadas en la presente investigación. Por ello, se comparan resultados de esta investigación con trabajos que incluyeron una o más de las razas utilizadas aquí. Para PN, los resultados difirieron de los encontrados por Williams et al. (2010) quienes estimaron efecto aditivos de la raza SIME (8.89 kg) superiores a los de BG (8.21 kg) y de LIM (6.08 kg), al expresar tales efectos como desviaciones de la raza Angus. Rodríguez-Almeida et al. (1997) trabajaron con dos conjuntos de datos en una región templada. Con el primero también encontraron efectos mayores para el SIME que para el LIM mientras que con el segundo hallaron un mayor efecto de LIM seguido de SIME y BV, hecho que difiere con los hallazgos del presente estudio. Resultados divergentes también fueron reportados por Thrift (1997), Elzo et al. (1998a) y Riley et al. (2007) quienes encontraron menores efectos de PN para la raza RS que para BG. Estos últimos trabajos fueron realizados en condiciones subtropicales. Para P4 no se encontraron referencias bibliográficas.
Para PD, nuestros resultados son diferentes a los descritos por Vergara et al. (2009), quienes hallaron efectos negativos para las razas BON y RS en el trópico bajo colombiano en una región clasificada como bosque húmedo tropical. Este estudio expresó los efectos aditivos de raza como desvíos del Cebú. Similares resultados fueron descritos por Elzo et al. (1998a) y Riley et al. (2007) en condiciones subtropicales.
Aquí se encontró que el efecto de LIM fue inferior al de Brahman (p<0.05) y el de SIME fue numéricamente inferior al de Brahman, resultados diferentes a los descritos por Williams et al. (2010), quienes encontraron un mayor efecto para SIME y para LIM que para BG. Otro estudio que involucró animales de razas taurinas y que se realizó en zona templada, reportó un mayor efecto para SIME que para LIM. Sin embargo, en este estudio el BV tuvo un menor efecto que el LIM (Rodríguez-Almeida et al., 1997), resultado que divergió con los del presente estudio.
En el caso de la variable PA los resultados encontrados son distintos a los descritos por Arboleda et al. (2008), quienes encontraron efectos negativos tanto para la raza BON (-0.11 kg) como para la raza RS (-0.05 kg). No se encontraron reportes para P15. Sin embargo, estudios realizados a una edad similar (18 meses) mostraron efectos negativos para las razas BON (-0.02 kg) y RS (-0.12 Kg) (Arboleda et al., 2008) a diferencia de los valores positivos encontrados en este estudio.
Para el carácter AOL, sin especificar una edad particular, se encontraron reportes de un mayor efecto de la raza LIM respecto a las razas SIME y BG (Williams et al., 2010), al igual que en el presente estudio y el de Ríos-Utrera et al. (2006) para AOL ajustada a los 432.5 días (14.4 meses) en una región templada (Nebraska, USA). Ellos encontraron un mayor valor para la raza LIM que para las razas BV y SIME. Horne (2004) reportó que los menores valores medios de AOL al momento del sacrificio fueron para animales BG puros y F1 producto de la cruza de animales BG y RS en una región subtropical. En este estudio se compararon animales puros y F1 producto de un apareamiento dialélico de tres razas: Angus, RS y BG. Sin embargo, Horne (2004) no separó los efectos aditivos raciales y los no aditivos de heterosis utilizando regresión como se hizo aquí, sino que trató cada raza pura o cruzamiento como un efecto de clasificación y comparó las medias de cada grupo vía pruebas de hipótesis múltiples. Por ello, las comparaciones de Horne (2000) involucraron efectos combinados aditivos y no aditivos. En el caso de la variable GD, Williams et al. (2010) hallaron menores valores para las razas SIME y LIM que para BG, resultados que coinciden con los reportados por Ríos-Utrera et al. (2006) y con los reportados en esta investigación para GDD. No se encontraron reportes de literatura para AOL4, GD4, AOLD, GDD, AOLA, ni para GDA.
Para el caso de aquellos estudios realizados en zonas templadas, las diferencias encontradas pueden atribuirse a las condiciones ambientales y de manejo, ya que los genotipos pueden presentar desempeños distintos en diferentes condiciones de entorno (Falconer y Mackay, 1996). En primer lugar, en tales zonas las condiciones medioambientales varían de manera notable a través del año a diferencia del trópico, en donde no se presentan estaciones. Además, los forrajes en zonas templadas tienden a tener un mayor valor nutricional que en zonas tropicales (Minson, 1990). En segundo lugar, el manejo también puede causar las diferencias en los resultados. En algunos de los estudios discutidos, las dietas de los animales se balancearon en cada estación con el fin de satisfacer los requerimientos nutricionales (Ríos-Utrera et al., 2006; Rodríguez-Almeida et al., 1997).
Si se consideran los estudios en los cuales las condiciones ambientales y el manejo fueron similares, las diferencias podrían deberse en parte a diferencias genéticas entre los grupos de animales de cada raza utilizados en cada estudio.
Los cambios de valores de efectos aditivos para ciertos grupos raciales de una edad a otra, pueden deberse a diferencias en las curvas de crecimiento de cada raza, pues es conocido que el componente genético tiene efecto sobre la trayectoria de crecimiento (Thornley y France, 2004; Agudelo- Goméz et al., 2007). Por ello, grupos genéticos diferentes pueden describir trayectorias de crecimiento distintas, lo cual genera que el ordenamiento de dichos grupos cambie en diferentes etapas del desarrollo. Esto puede ocurrir en caracteres diferentes al peso vivo como se evidenció en este trabajo para GD (Tabla 4).
Si se mira el desempeño de los grupos raciales a través de todas las edades, y se considera que para el peso al nacer no son deseables valores aditivos muy altos debido a la dificultad de parto que se asocia con los mismos, el grupo NOR presenta los mayores efectos genéticos a través de la etapa de crecimiento estudiada. Solamente para P12 no fue el grupo con el mayor valor, pero en este caso su valor fue muy cercano al de BV (el grupo con el mayor valor). Para AOL se observa que la raza LIM presentó los mayores valores a través de la etapa de desarrollo. Para este grupo todos los valores fueron significativos (p<0.05).
Para GD no se observó una tendencia clara en el comportamiento de los grupos genéticos. Debe considerarse que el espesor de grasa dorsal es un indicador de las reservas corporales del animal y que esta característica es influenciada por múltiples factores de origen tanto genético como no genético (Owenset al., 1995). Por esta razón, ante cualquier cambio del medio que implique movilización o acumulo de reservas, el espesor de grasa dorsal es uno de los primeros caracteres en presentar variación (Van Soest, 1994) y es posible que cada genotipo responda de manera diferente a tales variaciones (Cerón-Muñoz et al., 2004)
Efectos de heterosis
La falta de significancia de los efectos genéticos no aditivos fijos para la mayoría de caracteres fue debida en gran parte a los errores de estimación grandes, los cuales se presentaron debido al reducido tamaño de la población (propiedad de consistencia de los estimadores). Para PN los efectos no aditivos fueron levemente negativos, hecho favorable, ya que pesos altos al nacer se asocian con dificultad de parto. Mayores valores de heterosis para PN fueron reportados por Gregory et al. (1991) en una zona templada (1.4 kg para machos y 1.8 kg para hembras). Williams et al. (2010) encontraron valores desde 0.63 hasta 2.43 kg al fraccionar la heterosis en británica x británica, británica x Cebú, continental x continental y continental x Cebú. Los valores reportados por Riley et al. (2007) variaron entre 1.4 (heterosis RS x Angus) y 3.7 kg (Heterosis BG x Angus), la heterosis RS x Angus fue de 1.4 kg. En ninguno de estos estudios se reportaron valores negativos de heterosis directa para el peso al nacimiento. Sin embargo, Rodríguez-Almeida et al. (1997) hallaron valores desde -5 hasta 2.9 kg. Aumentos en el peso al nacimiento debidos a efectos intralocus interraciales en una población multirracial BG-Angus también fueron descritos por Elzo y Wakeman (1998).
Los valores de heterosis para PD fueron inferiores a los descritos por Vergara et al. (2009) (20.08 kg), Gregory et al. (1991) (20.4 kg para machos y 18 kg para hembras) y algunos de los reportados por Williams et al. (2010) (quienes encontraron valores entre 3.47 para heterosis continental x continental y 25.93 kg heterosis continental x Cebú) y Riley et al. (2007) (14.67 hasta 27.8 kg). El rango de valores encontrados por Rodríguez-Almeida et al. (1997) fue 3.5 a 14.5 kg. El valor de heterosis para PA fue negativo. Este resultado concuerda con el valor de -0.0247 kg para HTT y es inferior a los valores para HI (0.06 kg) y HCT (0.09 kg) reportados por Arboleda et al. (2008). Valores de heterosis muy superiores (28.8 kg para macho y 26 kg para hembras) fueron hallados por Gregory et al. (1991). Valores negativos de heterosis fueron descritos por Arboleda et al. (2008) para peso a los 18 y 24 meses (HTT y heterosis materna) y por Barreto (2005) para peso a los 390 días (-5.48 kg para heterosis entre alelos de origen cebuíno y taurino adaptado al trópico y -0.79 kg para la heterosis debida a la interacción de alelos de razas de origen británico y continental). Cabe destacar que estos dos estudios fueron realizados en zonas de trópico bajo.
Para una edad cercana a los 15 meses (18 meses), Arboleda et al. (2008) reportaron valores bajos, los cuales variaron entre -0.13 (para HTT) y 0.20 kg (para HI), estos valores fueron menores a los aquí descritos para P15.
Resultados inferiores de heterosis para AOL15 (heterosis media de 2.79 cm2, con valores mínimo y máximo de 2.29 y 3.11 cm2) fueron reportados por Ríos-Utrera et al. (2006). Este mismo trabajo reportó un valor medio de heterosis para GD de 0.02 mm y un rango de -0.01 a 0.03 mm, valores similares a los resultados de la presente investigación. Los valores de heterosis descritos por Williams et al. (2010) variaron entre 2.40 (heterosis británica x británica) y 6.57 cm2 (heterosis británica x Cebú). Para GD la heterosis varió entre -0.02 (heterosis británica x continental) y 0.16 mm (heterosis continental x Cebú).Williams et al. (2010) no especificaron la edad para la cual reportaron los valores de heterosis, los cuales fueron mayores a los hallados en este estudio para AOL4 y AOLD, similares para GDD y GD15, e inferiores a los encontrados para GD4 y GDA. No se encontraron reportes para AOL4, AOLD, AOLA, GD4, GDD y GDA.
Los efectos de heterosis indican que los efectos genéticos no aditivos tendrían importancia en PD y AOL a edades posteriores al destete.
En general, los mayores valores de heterosis encontrados en zonas templadas pueden obedecer a las mejores condiciones que se presentan en tales regiones en cuanto a manejo zootécnico y medio ambiente (calidad de las pasturas, menor incidencia de parásitos, etc.). Esto podría permitir que los efectos genéticos no aditivos se manifiesten en mayor medida. Otros factores que podrían haber contribuido a las diferencias entre valores de heterosis aquí y en otros estudios serían las razas utilizadas y la expresión utilizada para estimar heterosis.
En cuanto a las razas incluidas, es de resaltar que las interacciones entre razas continentales y cebuínas dependerán de las razas particulares de cada grupo que se consideren. Muchas de las razas taurinas reportadas en la literatura citada no se encuentran en el país, o se encuentran en un reducido número, y por lo tanto no se incluyeron en la presente investigación. Resultaría de interés la inclusión de tales razas en estudios futuros similares al presente, pues dada su ausencia o reciente introducción en el país, las recomendaciones para su uso en programas de cruzamiento deben obedecer a criterios objetivos.
Por otro lado, en ciertos trabajos se incluyeron solamente cruzamientos entre razas de origen Europeo, que para ciertos caracteres son similares o superiores a los encontrados en la presente investigación (Rodríguez-Almeida et al., 1997; Ríos-utrera et al., 2006). Sin embargo, en las condiciones de trópico bajo resulta necesario realizar cruzamientos entre genotipos adaptados a estas condiciones (razas nativas y cebuínas) y aquellos de alta producción, para que mediante la complementariedad entre los dos grupos se logre una progenie que resulte productiva y resistente (Barreto, 2005).
Para diferentes caracteres, los mayores valores de heterosis se dieron para efectos de interacción entre alelos de origen cebuíno y de origen taurino (continental o británico), mientras los menores valores fueron aquellos para interacciones del mismo grupo (p.e., continental x continental) (Williams et al., 2010; Arboleda et al., 2007). En el estudio de Arboleda et al. (2008) para P12 se encontraron efectos negativos de HTT y positivos de HTC, pero al ser mayores estos últimos, el efecto de HI (HTT + HTC) fue positivo. Esto muestra la necesidad de hacer una diferenciación entre los tipos de heterosis con el fin de encontrar las más favorables interacciones intralocus. Sin embargo, una de las grandes limitantes sería la estimabilidad de tales parámetros y el requerimiento de bases de datos de tamaño y con suficiente número de cruzamientos diferentes de dos o más razas representadas por animal.
Nuestros resultados sugieren que para los caracteres de crecimiento la raza NOR presentó los mayores efectos genéticos aditivos en condiciones de pastoreo en la región del sur del Cesar. Desde el punto de vista de la proyección hacia al producto final (p.e., carne despostada),los animales de la raza LIM serían una buena opción a considerar, debido al buen comportamiento que demostró para AOL. El uso de esta raza en cruzamientos produciría animales con un mayor rendimiento en cortes minoristas dada la alta correlación tanto genética como fenotípica de estos dos caracteres (Wilson et al., 1992).
Resultados de este estudio proveen información objetiva que ayudará al desarrollo e implementación de programas de cruzamientos en esta región del país. La elección de la o las razas a emplear deberá basarse en los objetivos productivos de cada explotación (venta de animales destetos, de un año, etc.) y el sistema de mercado (p.e., pago por composición de canal).
Los valores de heterosis sugieren que programas de cruzamiento similares a los aquí estudiados aumentarían la productividad fenotípica de la población bovina cruzada para PD, AOLA y AOL15. Sin embargo, los resultados de esta investigación deben ser validados con conjuntos de datos mayores y con mayor cantidad de cruzamientos, pues los errores de estimación debidos al bajo tamaño de la población pueden haber causado la falta de significancia de los efectos de heterosis en los demás caracteres. También, sería útil realizar estudios similares en una mayor cantidad de regiones.
Se requerirá de un trabajo organizado entre asociaciones ganaderas, productores y la comunidad académica, para obtener las estructuras de bases de datos que permitan modelar efectos genéticos no aditivos en mayor detalle. Esto permitirá obtener estimaciones de efectos raciales y de heterosis en combinaciones de razas específicas para los caracteres de peso y de canal medidos vía ultrasonido en poblaciones multirraciales en el trópico bajo colombiano.
Agradecimientos
Los autores expresan su gratitud con la Asociación Colombiana de Criadores de Ganado Cebú ASOCEBU por el apoyo económico, logístico y humano que permitió la obtención de los datos para la realización de la presente investigación.
Referencias
1. Agudelo-Gómez DA, Cerón-Muñoz MF, Restrepo LF. Modelación de funciones de crecimiento aplicadas a la producción animal. Rev Col Cien Pec 2007; 20:157-173. [ Links ]
2. Arboleda-Zapata EM, Cerón-Muñoz MF, Cotes-Torres JM. Livest Res Rural Dev 20 # 209 2008. Heterocigosis individual y materna en poblaciones bovinas multirraciales de diferentes sistemas para producción de carne en el trópico bajo Colombiano. [Fecha de acceso: Mayo 18 de 2011] URL:http://www.lrrd.org/lrrd20/12/arbl20209.htm [ Links ]
3. Barreto G. Estimação de efeitos genéticos aditivos diretos e maternos e nao aditivos e, predição do desempenho de pesos, perímetro escrotal e musculosidade em uma população de bovinos de corte compostos (Bos taurus x Bos indicus). Tese de PhD, Universidade- de São Paulo, Pirassununga, 2005. [ Links ]
4. Bertrand JK. Using actual and ultrasound carcass information in beef genetic evaluation programs. Rev Bras Zoot 2009; 38:58- 63. [ Links ]
5. Boldman KGA, Kriese LD, Van Vleck LP, Kachman SD. A manual for use of MTDFREML.A set of programs to obtain estimates of variances and covariances. Washington, D.C: ARS, USDA; 1995. [ Links ]
6. Cerón-Muñoz MF, Tonhati H, Costa CN, Maldonado-Estrada J, Rojas-Sarmiento D. Genotype X environment interaction for age at first calving in Brazilian and Colombian Holsteins. J Dairy Sci 2004; 87:2455-2458. [ Links ]
7. Cerón-Muñoz MF. Cruzamiento. En: Elzo MA, Cerón-Muñoz MF, editores. Modelación aplicada a las ciencias animales: Genética Cuantitativa. 1ª ed. Medellin: L. Vieco e hijas Ltda; 2009. p.105-140. [ Links ]
8. Elzo MA, Famula TR. Multibreed sire evaluation procedures within a country. J Anim Sci 1985; 60:942-952. [ Links ]
9. Elzo MA, Manrique C, Ossa G, Acosta O. Additive and non additive genetic variability for growth traits in the Turipaná Romosinuano-Zebu multibreed herd. J Anim Sci 1998a; 76:1539-1549. [ Links ]
10. Elzo MA, West RL, Johnson DD, Wakeman DL. Genetic variation and prediction of additive and non additve genetic effects for six carcass traits in an Angus-Brahman multibreed herd. J Anim Sci 1998b; 76:1810-1823. [ Links ]
11. Elzo MA,Wakeman DL. Covariance components and prediction for additive and non additive preweaning growth genetic effects in an Angus-Brahman multibreed herd. J Anim Sci 1998; 76:1290-1302. [ Links ]
12. Elzo MA, Martínez G, Gonzáles F, Huertas H. Variabilidad y predicciones genéticas aditivas, no aditivas y totales para la producción de ganado de carne en el rebaño multirracial Sanmartinero-Cebú de La Libertad. Rev Corpoica 2001; 3:51- 64. [ Links ]
13. Falconer DS, Mackay TFC. Introduction to quantitative genetics. 4thed. England: Longman limited; 1996. [ Links ]
14. Federación Nacional de Ganaderos FEDEGAN (Colombia). Plan estratégico de la ganadería colombiana 2019. Bogotá D.C: San Martín Obregón y Cía; 2006. [ Links ]
15. Gregory KE, Cundiff LV, Koch RM. Breed effects and heterosis. In advanced generations of composite populations for growth traits in both sexes of beef cattle. J Anim Sci 1991; 69:3202-3112. [ Links ]
16. Hausman GJ, Dodson MV, Ajuwon K, Azain M, Barnes KM, Guan LL, Jiang Z, Poulos SP, Sainz RD, Smith S, Spurlock M, Novakofski J, Fernyhough ME, Bergen WG. The biology and regulation of preadipocytes and adipocytes in meat animals. J Anim Sci 2009; 87:1218-1246. [ Links ]
17. Henderson CR. Theoretical basis and computational methods for a number of different animal models. Proceedings of the animal model workshop. J Dairy Sci 1988; 71 (Suppl 2):1-16. [ Links ]
18. Horne WJ. The effect of Romosinuano breeding on carcass characteristics, shear force, retail shelf life, and oxidative rancidity susceptibility of beef steaks. MS thesis. Texas Tech Univ, Texas, 2004. [ Links ]
19. Littell RC, Milliken GA, Stroup WW, Wolfinger RD, Schabenberger O. SAS® for mixed models. 2nd ed. Cary (NC): SAS Institute Inc.; 2006. [ Links ]
20. Martínez G. Plan nacional de acción para la conservación, mejoramiento y utilización sostenible de los recursos genéticos animales de Colombia. 1ª ed. Bogotá: Ministerio de Agricultura y Desarrollo Rural, república de Colombia– Organización de las Naciones Unidas para la Agricultura y la Alimentación, FAOColombia; 2010. [ Links ]
21. Minson DJ. Forage in ruminant nutrition. 1st ed. San Diego (CA): Academic Press , Inc.; 1990. [ Links ]
22. Owens FN, Gill DR, Secrist DS, Coleman SW. Review of some aspects of growth and development of feedlot cattle. J Anim Sci 1995; 73:3152-3172. [ Links ]
23. Riley DG, Chase CC, Coleman SW, Olson TA. Evaluation of birth and weaning traits of Romosinuano calves as purebreds and crosses with Brahman and Angus. J Anim Sci 2007; 85:289-298. [ Links ]
24. Ríos-Utrera A, Cundiff LV, Gregory KE, Koch RM, Dikeman ME, Koohmaraie M, Van Vleck LD. Effects of age, weight, and fact slaughter end points on estimates of breed and retained heterosis effects for carcass traits. J Anim Sci 2006; 84:63-87. [ Links ]
25. Rodríguez-Almeida FA, Van Vleck LD, Gregory KE. Estimation of direct and maternal breed effects for predictions of expected progeny differences for birth and weaning in three multibreed populations. J Anim Sci 1997; 75:1203-1212. [ Links ]
26. Sánchez JP, Misztal I, Aguilar I, Bertrand JK. Genetic evaluation of growth in a multibreed beef cattle population using random regression-linear spline models. J Anim Sci 2008; 86:267-277. [ Links ]
27. SAS Inst. Inc. (USA). SAS/STAT User's guide: Statistics. Cary, NC: The institute; 2008. [ Links ]
28. Thrift FA. Reproductive performance of cows mated to and preweaning performance of calves sired by Brahman vs alternative subtropically adapted breeds. J Anim Sci 1997; 75:2597-2603. [ Links ]
29. Thornley JHM, France J. Mathematical models in agriculture. Quantitative methods for Plant, Animal and Ecological sciences. 2nd ed. UK: Cromwell Press Trowbridge; 2007. [ Links ]
30. Van Soest PJ. Nutritional ecology of the ruminant. 2nd ed. Ithaca (NY): Comstock publishing associates; 1994. [ Links ]
31. Vergara OD, Ceron-Muñoz MF, Arboleda EM, Orozco Y, Ossa GA. Direct genetic, maternal genetic, and heterozygocity effects on weaning weight in a Colombian multibreed beef cattle population. J Anim Sci 2009; 87:516-521. [ Links ]
32. Williams JL, Aguilar I, Rekaya R, Bertrand JK. Estimation of breed and heterosis effects for growth and carcass traits in cattle using published crossbreeding studies. J Anim Sci 2010; 88:460-466. [ Links ]
33. Wilson DE. Application of ultrasound for genetic improvement. J Anim Sci 1992; 70:973-983. [ Links ]
Notas
¤ Para citar este artículo: Martínez CA, Manrique C, Elzo M, Jiménez A. Efectos aditivos de grupo genético y heterosis sobre caracteres de crecimiento y composición corporal en bovinos cruzados en el sur del Cesar, Colombia. Rev Colomb Cienc Pecu 2012; 25:377-390.

