










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690
Rev Colom Cienc Pecua vol.25 no.4 Medellín Oct./Dec. 2012
ORIGINAL ARTICLES
Phytoplankton and periphyton in ponds with Nile tilapia (Oreochromis niloticus) and bocachico (Prochilodus magdalenae)¤
Fitoplancton y perifiton en estanques con tilapia nilótica (Oreochromis niloticus) y bocachico (Prochilodus magdalenae)
Fitoplâncton e perifiton em viveiros com tilapia nilotica (Oreochromis niloticus) e bocachico (Prochilodus magdalenae)
José J García Gonzalez1, AE, MSc; Guillermo A Correa Londoño2, IF, PhD; Sandra C Pardo-Carrasco3*, MVZ, PhD.
1Universidad Nacional de Colombia Sede Medellín.
2 Faculty of Agricultural Sciences, Agronomic Engineering Department, Universidad Nacional de Colombia Sede Medellín, Colombia.
3 Faculty of Agricultural Sciences, Animal Production Department, BIOGEM, Universidad Nacional de Colombia Sede Medellín, Colombia.
* Corresponding author: Sandra C Pardo-Carrasco. Facultad de Ciencias Agrarias, Departamento de Producción Animal, BIOGEM, Universidad Nacional de Colombia, Sede Medellín. Calle 59A # 63-20, Bloque 50 oficina 314, Medellín, Colombia. E-mail: scpardoc@unal.edu.co
(Received: 2 april, 2012; accepted: 1 october, 2012)
Summary
Background: phytoplankton is considered the most important community in the aquatic food chain. As the basis of the food chain, periphyton offers possibilities for further optimization of extensive aquaculture ponds, although in-depth research on its effects on phytoplankton is necessary. Objective: a test was conducted to assess periphyton effects on phytoplankton concentration (org/L) and Shannon Weaver algae diversity index (DI). Methods: 18 earthen ponds (90 m2) under extensive polyculture conditions and low stocking density (2 kg/m2 at the end of the culture) were used. Fish density was 2.6 Nile tilapia/m2 and 0.7 bocachico/m2. Treatments consisted in the inclusion or absence of substrate (polyethylene pipes) for periphyton attachment (B1 for substrate presence and B2 for substrate absence) combined with three feeding strategies: no balanced feed offered (A1), 20% crude protein feed (A2), and 25% crude protein feed (A3). The combination of substrate levels and feeding strategies resulted in six treatments. Each treatment was replicated there times using a randomized complete block design for a six-month experimental period. Data were analyzed using a repeated-measure ANOVA in which time appears as a third factor (monthly samplings). Results: an interaction between factor B and time was found (P < 0.05), suggesting that the DI was significantly higher in ponds with substrate (B1) at given times (times 3, 4, 5 and 6). A significant interaction was also found between factors A*B*time (P < 0.05) for the concentration of organisms; resulting in a higher concentration of organisms in treatments that included substrate for periphyton attachment. Conclusion: periphyton had a positive influence on phytoplankton development, which is why stimulating periphyton attachment on an artificial substrate is a good way of improving a pond's primary production.
Key words: Cichlidae, fishculture, polyculture, primary production, Prochilodontidae.
Resumen
Antecedentes: se considera que la comunidad fitoplanctónica es la más importante en la cadena alimenticia acuática. El perifiton, como base de la cadena alimenticia, ofrece posibilidades que optimizarían el uso de los insumos utilizados en sistemas extensivos, pero es necesario investigar a fondo sus efectos sobre el fitoplancton. Objetivo: evaluar el efecto del perifiton sobre la concentración (org/L) y el índice de diversidad algal Shannon Weaver (DI) del fitoplancton. Métodos: se realizó un ensayo en 18 estanques en tierra, de 90 m2, bajo condiciones de policultivo de baja densidad de siembra (buscando alcanzar máximo 2 Kg/m2 al final del cultivo) con 2,6 tilapias y 0,7 bocachicos/m2. Los tratamientos consistieron en la inclusión o no de sustrato (tubos de polietileno) para perifiton (B1: presencia de sustrato; B2: ausencia de sustrato), bajo tres estrategias alimentarias: sin alimento balanceado, alimento con 20% proteína bruta (PB) y alimento con 25% de PB, denominadas A1, A2 y A3, respectivamente. La combinación de los tres niveles de A con los dos niveles de B resultó en seis tratamientos, cada uno con tres réplicas. Para la asignación de tratamientos a cada unidad se usó un diseño completamente al azar. Los datos fueron analizados usando un ANOVA con medidas repetidas, donde aparece el tiempo como un tercer factor (muestreos mensuales). Resultados: hubo una significativa interacción entre el factor B y el tiempo (P < 0.05), mostrando que en algunos tiempos específicos (3, 4, 5 y 6), el DI fue significativamente más alto en estanques con sustrato (B1). También se encontró interacción entre los factores A*B*tiempo (P < 0.05) para la concentración de organismos, mostrando una mayor concentración para los tratamientos con sustrato para perifiton. Conclusión: el perifiton tuvo una influencia positiva sobre el desarrollo del fitoplancton. Por esta razón, estimular la fijación de perifiton sobre un sustrato artificial es una buena forma para mejorar la productividad primaria del estanque.
Palabras clave: Cichlidae, piscicultura, policultivo, producción primaria, Prochilodontidae.
Resumo
Antecedentes: considera-se que a comunidade fitoplanctônica é a mais importante na cadeia alimentar aquática. O perifíton, como base da cadeia alimentar, oferece possibilidades para melhorar o uso dos insumos correntemente utilizados nos sistemas extensivos, mas precisa se pesquisar bem acerca de seus efeitos sobre o fitoplâncton. Objetivo: avaliar o efeito do perifíton sobre a concentração (org/L) e o índice de diversidade Shannon Weaver (DI) do fitoplâncton. Métodos: realizou-se um teste em 18 viveiros, de 90 m2 sob condições de policultivo de baixa densidade de estoque (procurando alcançar no máximo 2 Kg/m2 no final da safra) com 2,6 tilápias e 0,7 bocachicos por m2 respectivamente. Os tratamentos consistiram na inclusão ou não de substrato (canos de polietileno) para perifíton (B1 presença de substrato, B2 ausência de substrato), sob três estratégias alimentarias: sem alimento balanceado, alimento com 20% de proteína bruta (PB) e alimento com 25% de PB, denominados A1, A2 e A3, respectivamente. A combinação dos três níveis de A, com os dois níveis de B resultou em seis tratamentos, cada um com três replicas. Na designação dos tratamentos para cada unidade experimental usou-se um desenho completamente ao acaso. Os dados foram analisados usando ANOVA com medidas repetidas, onde aparece o tempo como o terceiro fator (amostras mensais). Resultados: houve uma significativa interação entre o fator B e o tempo (P < 0.05), demonstrando que em alguns tempos específicos (3, 4, 5 e 6), o DI foi significativamente maior em viveiros com substrato (B1). Também se encontrou interação entre os fatores A*B*tempo (P < 0.05) para a concentração de organismos, apresentando maior concentração para os tratamentos com substrato para perifíton. Conclusão: o perifíton teve uma influencia positiva sobre o desenvolvimento do fitoplâncton. Por esta razão, estimular a fixação do perifíton sobre um substrato artificial é uma boa maneira para melhorar a produtividade primaria do viveiro.
Palavras-chave: Cichlidae, piscicultura, policultivo, produção primaria, Prochilodontidae.
Introduction
Besides being a food source, phytoplankton also plays a series of important roles in aquatic ecosystems. As a primary producer, phytoplankton is responsible for CO2 fixation, meaning that it is a primary energy source for low trophic level organisms (Azim et al., 2005). Considering these phytoplankton characteristics, the bocachico and tilapia species, both low trophic level fish, were chosen for this study due to their anatomical characteristics.
Nile tilapia are herbivorous filter feeders. Their gill rakers have a wide contact surface for filtering plankton components. Likewise, they can use their mouth to browse or scrub those areas where other autotrophic and heterotrophic organisms attach. Bocachicos, in turn, are a benthic, detritivorous-iliophagous species with lips especially adapted for browsing (Gneri and Angelescu, 1951).
Azim (2002), Dodds (2003), and Bender et al. (2004) pointed out that the functions of autotrophic periphyton are similar to those of phytoplankton. These similarities include the catching of organic detritus, removal of nutrients from the water column, and helping to control oxygen and pH concentrations in the surrounding water. Furthermore, periphyton assimilate nutrients and subsequently transform them into periphytic biomass that can be later consumed by culture organisms. The biomass generated by phytoplankton and periphyton has the advantage of being attached to a submerged substrate, facilitating consumption by other organisms.
Having a submerged substrate is important because it could increase economic competitiveness in low load production systems like the one used in this study. It is also vital to find environmentally sustainable complements and feeding alternatives.
Likewise, it is important to reduce the nitrogen waste generated by the remains of unconsumed feed. In fact, in traditional systems, only 15% to 30% of protein is converted into fish meat; the rest is disposed in water, thus contributing to pollution (Gross et al., 2000). Periphyton is therefore an economically and environmentally feasible option, given the ability to recirculate nutrients in the water column and the capacity of supplementing feeding rations with natural food.
Considering the above, it was expected that the improvement in physical and chemical conditions of water surrounding periphytic communities would have an effect on the number of organisms and taxa which make up the plankton. Therefore, this experiment was conducted to determine the effects of a substrate (polyethylene pipes) for periphyton attachment as well as the feeding strategy on the organisms concentration (org/L) and on their diversity index over time.
Materials and methods
Ethical considerations
This research was approved by the Animal Experimentation Ethics Committee of the Fish Research Center (CINPIC), University of Córdoba, Colombia (CINPIC 001 from February 16, 2009).
Type of study
The experiment was conducted in 18 rectangular earthen ponds, each measuring 90 m2, with a depth of 1 m. Experimental units belong to Piscícola Meléndez, located in Cereté municipality (Córdoba province, Colombia). A total of six treatments with three replicates were used. Each treatment was a combination of two factor levels. Factors were feeding strategy (factor A) and substrate (factor B). Each factor had different levels: for factor A, the levels were 0% (indicating that no fish feed was supplemented), 20% (a commercial ration with 20% crude protein or CP), and 25% (a commercial ration with 25% crude protein or CP). Factor B had two levels: presence or absence of substrate, such as additional surfaces to which periphytic communities could attach.
Ponds were previously dried and cleaned. Soil drying and handling took approximately three days. During this time, soil samples were analyzed for pH and nutrient content. Depending on the soil pH levels, liming was applied. If necessary, the pH was rectified using calcium carbonate. The soil was disinfected using quicklime at a ratio of 50 g/m2.
The ponds were populated with Nile tilapia fingerlings of the chitralada variety. These fingerlings were acquired from a company located in the area, which guaranteed homogeneity and a high rate of sexual reversion. Bocachico fingerlings were supplied by the Center for Fish Research at Córdoba University (CINPIC) and stocked at a rate of 243 tilapias to 63 bocachicos, for a density of 2.6 tilapias/m2 and 0.7 bocachicos/m2 (total density: 3.3 fish/m2).
In this study, the additional areas were plastic polyethylene pipes with a diameter of 6 cm and a length of 1.2 m. Pipes were deployed at a rate of 3.3 pipes/m2. Pipes provided an additional attachment surface of 67% for each pond. The pipes were first suspended in a vertical position inside the ponds and then filled with water.
Twenty days after flooding, periphyton attachment on the substrate was verified and fish were stocked. Initially, fish were fed with an amount of fish feed equal to 12% of the culture's biomass. This amount was progressively reduced based on average weight until the culture's biomass reached 1.1%. Culture time was eight months. The analyses of planktonic organisms and the biometries for adjusting feeding rations were conducted on a monthly basis beginning at month two. Besides substrate and feeding strategy, time was also included as the experimental units were measured successively over time.
Throughout the experimental period, the water's physical and chemical parameters were analyzed. Oxygen was determined using a digital oximeter (YSI 550A, USA), pH was determined using a digital pH meter (YSI 100, USA) and the Secchi disk visibility. All measurements occurred between 08:00 to 09:00 hours and 17:00 to 18:00 hours.
For plankton characterization, 3 liters of water were collected from the first 50 cm of each pond. In order to collect an integrated sample reflecting plankton distribution in the water column, samples were taken from different areas of the experimental units. Then, 50 ml of an aqueous formaldehyde solution at 10% and 5 ml of Lugol's iodine were added to the sample and allowed to settle for 24 hours. Once the plankton settled, the sample was reduced to 100 ml in another 100 ml cylindrical container for an additional 72 hours. Finally, the supernatant was removed, reducing the sample to 50 ml. A sub-sample of such reduction was transferred to a Neubauer chamber to determine plankton concentration. Then, the genera found were characterized following the methodology proposed by Azim et al. (2001).
Statistical analysis
The covariance structures that Littel et al. (1998) suggested as the most appropriate were adjusted for each of the response variables. Structures included compound symmetry, first-order regressive, and unstructured. Finally, the ANOVA corresponding to the best structure, as shown by the Bayesian information criterion (BIC), was evaluated. Simple and main effects were analyzed using Tukey's test at 5%.
An HJ-biplot representation (Galindo, 1986) was used to analyze which plankton genera were more numerous based on the influence of the factors in question. A routine created with MATLAB® was used for this purpose.
Results
The ANOVA for the concentration of phytoplanktonic organisms (org/L) showed a significant second-order interaction between factors A, B, and time (P=0.042). As a result, analysis of the simple effects of A*B interaction at specific times showed that treatments including substrates for periphyton (B1) had a significant and positive influence on the amount of plankton in the experimental units. The average concentration of organisms found in the experimental units with substrate was 110,918 org/L, whereas the units without substrate (B2) had an average concentration of 32,832 org/L (Table 1)(Table 2).
Regarding the diversity index, no significant interaction was found between the feeding strategy and any other factor. Likewise, the main effect of this index was not significant (P=0.684). Moreover, the significance of the B*Time interaction (P=0.012) for the diversity index indicated that the differences between the levels of factor B were not consistent over time (Table 3). Upon analyzing factor B's simple effects at times 3, 4, and 6 (Table 3), it could be observed that this index was statistically different when the treatments that included substrates for periphyton attachment were compared with the treatments without substrate.
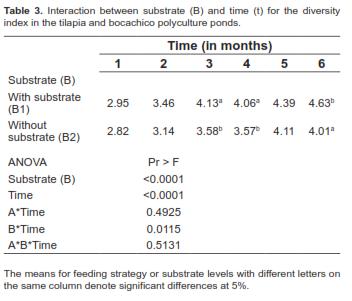
Phytoplanktonic organisms have an increasing tendency to concentrate over time. For most comparisons, the difference between one given time period and the other is significant. There is an exception, however, when times 2 (August) and 3 (September) are compared (P=0.734). In these cases, the observed concentration of planktonic organisms fluctuated between 51,000 and 53,000 org/L.
The analysis of the simple effects of the A*B interaction at specific times showed no significant difference for plankton concentration (org/L) between treatments with substrates, nor between the treatments without substrate. Conversely, there were significant differences between the group of treatments with substrate and the group of treatments without substrate (Figure 1). The analyzed water samples showed a total of 38 genera of phyto- and zooplankton. For a list of these genera, see table 4.
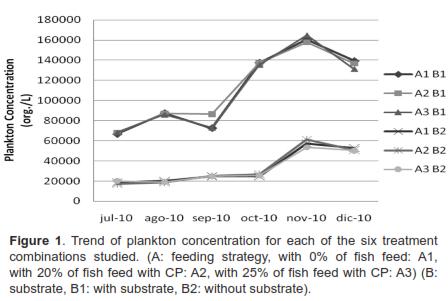
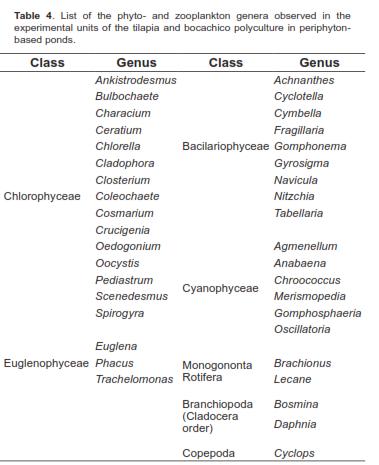
The Chlorophyceae class was predominant during the study, as 39% of the genera observed belonged to it. Following this class were Bacilariophyceae (24%), Cyanophyceae (16%), and Euglenophyceae (8%). Cladocerans and rotifers were less predominant, with a presence of 5%, and copepods, representing only 3% of the total genera found (Table 3). An HJ-Biplot representation was obtained in order to observe association trends between genera and the studied factors, namely feeding strategy (A) and substrates for periphyton attachment (B) (Figure 2). Similar to the results of the ANOVA for the plankton concentration (org/ml), the HJ-Biplot showed that the experimental units with substrates for periphyton attachment had richer microorganism populations. Likewise, a clear separation could be seen between the ponds without substrates for periphyton attachment (represented by triangles) and the ponds that did have these substrates (represented by circles).
Of equal importance is the observation that the protein levels making up the feeding strategy had no effect on the appearance of any particular taxon, including plankton concentration or abundance. This was evidenced by the fact that protein levels for each feeding strategy (0%, 20% and 25%) represented by three colors (white, gray and black, respectively) did not separate from each other in the treatments with or without substrates for periphyton attachment. Although these substrates generally allow for a richer population of microorganisms, in this study the species from Trachelomonas, Characium, and Tabellaria genera benefitted from the absence of substrates for periphyton attachment. Eventually, the species from the Spyrogira, Closterium, and Scenedesmus genera could also benefit from the absence of substrates.
Overall, water quality remained unaffected during the culture process, except for the values related to dissolved oxygen, transparency, and temperature. The analysis of the simple effects of factor A on dissolved oxygen at specific times revealed significant differences only from time 5 onwards (Table 5). These differences were always in favor of the ponds without feeding (A3). Factor B, in turn, had no significant effect on dissolved oxygen while significantly affecting transparency (P=0.009), as this variable had lower values in the treatments with substrates for periphyton attachment (Table 2).
Similarly, a significant interaction was found between feeding strategy (A) and time (t) for the visibility variable (Table 6). Another significant difference (P=0.038) of 3.7 cm was detected in the photic zone when comparing feeding strategies A2 (13.74 cm) and A1 (10.08 cm) at specific time 6.
Discussion
As a food source, plankton plays a key role in productive ponds, provided that these units have extensive or semi-intensive production systems and that the environmental conditions allow for the development of these plankton communities. As a component of these associations, phytoplankton also plays a key role as a primary producer and as the base of the trophic chain, supporting the other trophic levels.
Similarly, plankton is a good indicator of the production conditions of a pond, as these organisms act as a direct or indirect source of food for fish and other culture organisms (Azim et al., 2001). Although phytoplankton is notably predominant in terms of energy attachment and feeding in traditional systems, it is more dynamic and efficient in systems with additional areas for attachment. For some culture organisms, periphyton is a source of food that can be consumed directly by browsing or scrubbing the areas where it is attached. Otherwise, it would not be possible to use periphyton as we do. Many of the species of interest for aquaculture lack structures (i.e. gill rakers) for taking plankton directly from the water column, therefore, two or more trophic levels would have to be involved before this phytoplankton could be converted to harvestable biomass.
Aquacultural production systems with additional areas have the appropriate conditions for adequate periphyton colonization, but the development of this periphyton involves positive and negative interactions between phytoplankton, autotrophic and heterotrophic periphyton, heterotrophic microbial communities from the bottom of the pond, and of course, the fish species in the culture (Milstein, 2005). In this study in particular, plankton counts decreased gradually over time for treatments without substrates for periphyton. Also, the plankton population in these treatments was 30% of the total amount of plankton found in the treatments with substrates for periphyton. This difference could be explained by competition between the periphyton attached to the substrates and the plankton in the water column. This is favored by the fact that water quality is strongly influenced not only by the manner in which production ponds are handled, but also by stocking density and by the quantity and quality of nutrients (Milstein, 1993).
The two factors mentioned above play a key role in the development of periphytic communities and in the increase of algae biomass (Cuker, 1983; Fairchild, 1985). This is due to the fact that although periphyton attached to substrates recycles nutrients, the substrates not only serve as the basis for increasing periphytic biomass, but also improve physical and chemical conditions of the surrounding water column. Thus, nutrients that increase the amount of plankton, especially its autotrophic component (phytoplankton) are made available. Similarly, thanks to conceptual models such as the ones proposed by Azim et al. (2003), it is known that the main sources of variability in periphyton-based systems, either with fertilization or without it (as in this experiment), are the increase in periphytic mass and its effect on turbidity. In the case of this study, turbidity was higher in the ponds with substrates (as shown by the Secchi disk test) (Table 2), and the difference between turbidity in ponds with substrate and those without it was statistically significant (P=0.009). Due to this, the photic zone for periphytic strata was reduced, causing photosynthetic and renovation processes to either stop or slow down. This logically caused detachment of the periphyton layers, which in turn raised water turbidity and at the same time generated nutrients for the plankton. All of this ultimately restricted the photic zone to 10 or 11 cm in treatments with substrates.
Similarly, it is necessary to take into account the percentages of protein in food rations, as the latter provide nitrogen waste when they are not provided or balanced enough (Jiménez-Montealegre et al., 2002). Nitrogen waste is responsible for algal blooms that reduce the photic zone. In fact, a reduction of 3.7 cm was observed in the photic zone with the feeding strategy whose formula had 20% crude protein. This, however, is not conclusive and further research is required. As for the oxygen dissolved in the water, it is important to notice that the dynamics observed were closely related to the phenomena described above. Treatments with no fish feed administered had higher amounts of dissolved oxygen than the ones that received fish feed. This was obviously caused by the fishes' higher metabolic rates for food assimilation, the remains of uneaten food, fish faeces, and the nitrification and denitrification processes used by the bacteria in the water column to transform this waste into compounds that phytoplankton can assimilate, e.g nitrate (NO3) (Turker et al., 2003). As a result, there was an increase in phytoplankton biomass.
In contrast, factor A, feeding strategy, showed no significant effect on the Shannon-Weaver diversity index. This trend was supported by the HJ-biplot representation (Figure 2), which showed no specific plankton genera grouping patterns in terms of the different constitutive levels of the aforementioned factor (A1, A2, and A3). It is worth noting that under the conditions of this experiment, temperature was also affected in the experimental units with substrates, which showed lower temperature values. This somewhat explains the gradual loss of the photic zone. Although plankton concentration was higher in ponds with substrates, this progressive growth began to decrease as a result of the loss of the photic zone. In fact, the concentration showed a decrease in the samples taken in the sixth month.
Regarding water transparency, browsing by fish in the culture is an important factor that must be taken into account. Although it is not the main focus of this study, there is background research such as that of Havens (1991) and Tátrai et al. (1997) which places a great deal of importance on the turbulence caused by fish, particularly by Nile tilapia, one of the two species chosen for this study's polyculture. Another focus of interest in those studies was the effects of such turbulence on the sedimentation of the total solids and on the resuspension of organic matter. These two factors are highly correlated with fish biomass, total suspended solids, food supply, and Secchi disk visibility.
Additionally, since bocachico was the other species in the polyculture and is a species with completely benthophagous habits, they were likely to generate even more bio-turbulence, and therefore higher rates of resuspension (Yossa et al., 2009). According to Breukelaar et al. (1994) and Scheffer et al. (1998) in the studies they conducted on carps, resuspension rates (expressed in resuspended kg/day) were five times greater than the amount of fish biomass. This is why further and more specific research is required. For instance, different stocking densities for the polyculture's species (Nile tilapia and bocachico) could be assessed whilst quantifying the possible effects on the levels of solids in suspension and organic matter resuspension. It cannot be ruled out that this could also be another reason for the low photic zones observed throughout the study.
Regarding plankton characterization, it was observed that out of the 38 genera found, six had a tendency to appear in the treatments without substrates (Figure 2). These genera belong to the autotrophic component of plankton (Trachelomonas, Characium, Tabellaria, Spyrogira, Closterium and Scenedesmus). This is consistent with the studies conducted by Azim et al. (2001), in which genera in plankton and periphytic communities were identified in a carp culture. The researchers found genera exclusive to periphyton and other genera exclusive to plankton. Regarding this, our study shows that the six genera previously mentioned, except for the algae from the Closterium genus, belong to the plankton-only group.
Similar to the findings of a study conducted by Azim et al. (2001A), in which the rate of Clorophyceae in plankton was 35% of the genera found, our results show a predominance of 39% of the genera belonging to this class. Nevertheless, it is worth noting that algae from genera that frequently appear in periphytic communities were also found in the plankton. These algae were Achnantes, Fragillaria, Nitzschia, Ceratium, Chlorella, and filamentous algae such as Oedogonium, Bulbochaete, Coleochaete, and Anabaena, consistent with the findings of Ibarra et al. (2009) and Azim et al. (2005). This could be the result of the detachment of chunks from the periphyton matrix due to a fish's browsing and the resuspension it causes. This is obviously true only for the treatments with submerged substrates for periphyton attachment.
The concentration of planktonic organisms is higher in treatments with substrates for the attachment of periphyton. However, we must bear in mind that due to the second degree interaction among Food Strategy, Substrate, and Time on plankton concentration, this is an additive effect of the three factors, so it must be considered as a whole. Therefore, it is not possible to attribute this fact to one single factor, but to the three factors given above as a whole. Likewise, the study of protein percentages in feeding strategies is very important, since, as evidenced by this study, they can affect the amount of dissolved oxygen in water. This is probably due to the fact that more nitrogen (a major component of protein) implies more oxygen intake for the creation of nitrogen compounds that can be assimilated by aquatic ecosystems (McGraw et al., 2001) and, in consequence, by plankton communities.
Finally, it is also worth noting that, although no effect of the feeding strategy was detected on the diversity of plankton genera, the substrate factor was found to have an effect on such diversity over time. The diversity index was higher when substrate was included; suggesting that further studies could consider using substrates to stimulate specific planktonic genera, with greater benefits for the production system.
Acknowledgements
This study was fully sponsored by the MADR-UNALMED-UNICORDOBA Project, code 234-2008U62189-6087.
References
1. Azim ME, Wahab MA, Van Dam AA, Beveridge MCM, Verdegem MCJ. The potential of periphyton-based culture of two Indian major carps, rohu Labeo rohita (Hamilton) and gonia Labeo gonius (Linnaeus). Aquac Res 2001; 32:209-216. [ Links ]
2. Azim ME, Wahab MA, Van Dam AA, Beveridge MCM, Milstein A, Verdegem MCJ. Optimization of fertilization rate for maximizing periphyton production on artificial substrates and the implications for periphyton-based aquaculture. Aquac Res 2001A; 32:749-760. [ Links ]
3. Azim ME, Wahab MA, Verdegem MCJ, Van Dam AA, Van Rooij JM, Beveridge MCM. The effects of artificial substrates on freshwater pond productivity and water quality and the implications for periphyton-based aquaculture. Aquat Living Resour 2002; 15:231-241. [ Links ]
4. Azim ME, Milstein A, Wahab MA, Verdegam MCJ. Periphyton–water quality relationships in fertilized fishponds with artificial substrates. Aquaculture 2003; 228:169-187. [ Links ]
5. Azim ME, Beveridge MCM, Van Dam AA, Verdegem MCJ. Periphyton and aquatic production: an introduction. In: Azim ME, Verdegem MCJ, van Dam AA, Beveridge MCM, editors. Periphyton Ecology, Exploitation and Management. Wallingford: CABI Publishing; 2005. p.1-13. [ Links ]
6. Bender J, Lee R, Sheppard M, Brinkley K, Philips P, Yeboah Y, Wah RC. A waste effluent treatment system based on microbial mats for black sea bass Centropristis striata recycledwater mariculture. Aquac Eng 2004; 31:73-82. [ Links ]
7. Breukelaar AW, Lammens EHRR, Breteler JGPK, Tátrai I. Effects of benthivorous bream (Abramis brama) and carp (Cyprinus carpio) on sediment resuspension and concentrations of nutrients and chlorophyll-a. Freshwater Biol 1994; 32:113-121. [ Links ]
8. Cuker BE. Grazing and nutrient interactions in controlling the activity and composition of the epilithic algal community of an artic lake. Limnol Oceanogr 1983; 28:133-141. [ Links ]
9. Dodds WK. The role of periphyton in phosphorus retention in shallow freshwater aquatic systems. J Phycol 2003; 39:840-849. [ Links ]
10. Espinal GC, Martínez CH, González RF. 2005. La cadena de la piscicultura en Colombia: una mirada global de su estructura y dinámica, 1991-2005 Documento de Trabajo No. 106. [ Links ]
11. Fairchild GW, Lowe RL, Richardson WB. Algal periphyton growth on nutrient-diffusing substrates: An in in situ bioassay. Ecology 1985; 66:465-472. [ Links ]
12. Galindo MP. Una alternativa de representación simultánea: HJ-biplot. Questió 1986; 10:13-23. [ Links ]
13. Gneri FS, Angelescu V. La nutrición de los peces iliófagos en relación con el metabolismo general del ambiente acuático. Rev Inst Nal Inv Cienc Nat 1951; 2:1-44. [ Links ]
15. Gross A, Boyd CE, Wood CW. Nitrogen transformations and balance in channel catfish ponds. Aquac Eng 2000; 24:1-14. [ Links ]
16. Havens KE. Fish-induced sediment resuspension: effects on phytoplankton biomass and community structure in a shallow hypereutrophic lake. J Plankton Res 1991; 13:1163-1176. [ Links ]
17. Ibarra CR, Tavera E, Novelo E. Diversity and structure of periphyton and metaphyton diatom communities in a tropical wetland in Mexico. Rev Mex Biodivers 2009; 80:763-769. [ Links ]
18. Jiménez-Montealegre R. Nitrogen transformation and fluxes in fish ponds: a modeling approach. PhD thesis. The Netherlands: Wageningen University; 2001. [ Links ]
19. Jiménez R, Verdegem MCJ, Van Dam A, Verreth JAJ. Conceptualization and validation of a dynamic model for the simulation of nitrogen transformations and fluxes in fish ponds. Ecol Model 2002; 147:123-152. [ Links ]
20. Littell RC, Milliken GA, Stroup WW, Wolfinger RD. 1996. SAS® System for Mixed Models. SAS Institute. Cary (North Carolina, USA), 633 p. [ Links ]
21. Littell RC, Henry PR and Ammerman CB. Statistical Analysis of repeated measures using SAS procedures. J Anim Sci 1998; 76: 1216-1231. [ Links ]
22. McGraw W, Teichert DR, Rouse DB, Boyd CE. Higher minimum dissolved oxygen concentrations increase penaeid shrimp yields in earthen ponds. Aquaculture 2001; 199:311-321. [ Links ]
23. Milstein A. Water quality and freshwater fish culture intensification: the Israeli example. Aquac Fish Manage 1993; 24:715-724. [ Links ]
24. Milstein A. Effect of periphyton on water quality. In: Azim E, Verdegem M, Van Dam A, Beveridge M, editors. Periphyton: Ecology, Explotation and Management. Oxfordshire: CABI Publishing; 2005. p. 179-190. [ Links ]
25. Scheffer M. Ecology of Shallow Lakes. New York: Chapman and Hall; 1998. [ Links ]
26. Tátrai I, Oláh J, Paulovits G, Mátýas K, Kawiecka BJ, Józsa V, Pekár F. Biomass dependent interactions in pond ecosystems: responses of lower trophic levels to fish manipulation. Hydrobiologia 1997; 345:117-129. [ Links ]
27. Turker H, Eversole AG, Brune D. Comparative Nile tilapia and silver carp filtration rates of Partitioned Aquaculture System phytoplankton. Aquaculture 2003; 220:449-457. [ Links ]
28. Yossa MI, Hernández G, Vasquez W. Efecto del ''coporo'', Prochilodus mariae (Caharaciformes: Prochilontidae), sobre la calidad del agua en sistema de policultivo. Actual Biol 2009; 31:199. [ Links ]
Notas al pie
¤ To cite this article: García JJ, Correa GA, Pardo-Carrasco SC. Phytoplankton and periphyton in ponds with Nile tilapia (Oreochromis niloticus) and bocachico (Prochilodus magdalenae). Rev Colomb Cienc Pecu 2012; 25:603-614.
