










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690
Rev Colom Cienc Pecua vol.27 no.3 Medellín July/Sept. 2014
ORIGINAL ARTICLES
Effects of phytase supplementation on reproductive performance, apparent total tract digestibility of Ca and P and bone characteristics in gestating and lactating sows¶ ¤
Efectos de la suplementación con fitasa sobre el comportamiento reproductivo, la digestibilidad aparente en tracto total de Ca y P y las características óseas en cerdas gestantes y lactantes
Efeitos da suplementação com fitasse sobre o desempenho reprodutivo, digestibilidade aparente no trato digestivo total de Ca e P e características ósseas em porcas gestantes e lactantes
Young Dal Jang1, Anim Sci, PhD; Merlin D Lindemann1, 6*, Anim Sci, PhD; Eric van Heugten2, 6, Anim Sci, PhD; Richard D Jones3, Anim Sci, PhD; Beob Gyun Kim1†, Anim Sci, PhD; Charles V Maxwell4,6, Anim Sci, PhD; J Scott Radcliffe5,6, Anim Sci, PhD.
1 Department of Animal and Food Sciences, University of Kentucky, Lexington KY 40506, USA.
2 North Carolina State University, Raleigh NC 27695, USA.
3 Department of Animal Science, University of Georgia, Athens, GA 30602, USA.
4 Department of Animal Science, University of Arkansas, Fayetteville AR 72701, USA.
5 Department of Animal Sciences, Purdue University, West Lafayette, IN 47907, USA.
6 S-145 Regional Research Committee on Nutrition and Management of Swine for Increased Reproduction Efficiency.
* Corresponding author: Merlin D. Lindemann, Department of Animal and Food Sciences, University of Kentucky, Lexington KY 40506. Tel: 859 257 7524; Fax, 859 323 1027; Email: merlin.lindemann@uky.edu
† Current address: Department of Animal Science and Technology, Konkuk University, Seoul 143-701, South Korea.
(Received: January 21, 2014; accepted: April 16, 2014)
Summary
Background: phytase supplementation to sow diets may improve digestibility of P and Ca and bone integrity of suckling piglets. Objective: two experiments were conducted to investigate the effect of phytase supplementation to gestation and lactation diets on reproductive performance, apparent total tract digestibility (ATTD) of P, Ca, and N in sows and bone characteristics in sows and piglets. Methods: in Experiment 1, a multistate study involving 204 sows was conducted at 3 cooperating research stations. Sows were assigned to 1 of 4 treatments at breeding: 1) negative control [low Ca and available P (aP) diet], 2) Treatment 1 plus 300 FTU of supplemental phytase/kg of diet, 3) Treatment 1 plus 600 FTU of supplemental phytase/kg of diet, and 4) positive control (normal Ca and aP diet). In Experiment 2, a total of 16 sows (Yorkshire × Landrace; average parity, 2.3) were randomly allotted to 1 of 3 treatments based on body weight (BW) and parity at d 34 to 52 of gestation. Treatments were: 1) low Ca and aP diet, 2) low Ca and aP diet with 500 FTU of supplemental phytase/kg of diet, and 3) normal Ca and aP diet. Results: in Experiment 1, the number of total born (p<0.05), live born (p=0.05), litter weight of live born (p<0.01) and weaning pigs (p<0.05) were increased in a quadratic manner by increasing phytase supplementation level up to 300 FTU/kg. There was a linear increase in piglet weaning weight (p<0.05) by increasing supplemental phytase levels. There were no differences in reproductive performance in Experiment 2 (p>0.10). In gestation the ATTD of P was lower for the low Ca and aP without phytase group than the normal Ca and aP (p<0.01) and phytase-supplemented (p<0.05) groups. The ATTD of Ca in the normal Ca and aP group was higher than that in low Ca and aP without phytase group (p<0.05). In lactation, the phytase-supplemented group had higher ATTD of P compared with the other groups (p<0.05) whereas the ATTD of Ca in phytase-supplemented group was higher than that in low Ca and aP without phytase group (p<0.10). When the digestibility data was pooled for both gestation and lactation periods, the ATTD of P in the phytase-supplemented group was the highest among dietary treatments (p<0.05). There were no phytase effects on sow bone ash content at weaning, and piglet bone strength and ash content at birth. However, piglets from the phytase-supplemented sows had higher bone strength (p<0.10) and ash content (p<0.05) at weaning compared with those from non-phytase sows. There were positive correlations at birth between piglet BW and bone strength (p<0.01) and between bone strength and ash content (p<0.001). Additionally, at weaning positive correlations between piglet BW and bone ash content (p<0.05), between piglet BW and bone strength (p<0.001), and between bone strength and ash content (p<0.05) were observed. Conclusion: phytase supplementation from mid-gestation through lactation increased the ATTD of P and Ca both for gestating and lactating sows, improved bone integrity of their progeny, and had potential to increase litter size and performance but did not affect bone characteristics of sows and newborn pigs.
Key words: bone traits, exogenous enzyme, nutrient availability, phosphorus, pig.
Resumen
Antecedentes: la adición de fitasa a la dieta de cerdas puede mejorar la digestibilidad del P y Ca, así como la integridad ósea de los cerdos lactantes. Objetivo: se realizaron dos experimentos para investigar el efecto de la suplementación con fitasa en dietas de gestación y lactancia sobre el rendimiento reproductivo, la digestibilidad aparente de tracto total (ATTD) del P, Ca y N en cerdas, y las características óseas de cerdas y lechones. Métodos: el Experimento 1 involucró 204 cerdas evaluadas en 3 centros de investigación norteamericanos. Las cerdas fueron asignadas a 1 de 4 tratamientos al momento del servicio: 1) Control negativo [dieta baja en Ca y P disponible (aP)], 2) Tratamiento 1 más 300 unidades de fitasa (FTU)/kg de dieta, 3) Tratamiento 1 más 600 FTU/kg de dieta, y 4) Control positivo (dieta con contenido normal de Ca y aP). En el Experimento 2, un total de 16 cerdas (Landrace × Yorkshire, de 2,3 partos en promedio) se asignaron al azar a 1 de 3 tratamientos en el d 34 a 52 de gestación, con base en su peso corporal (BW) y número de partos. Los tratamientos fueron: 1) dieta baja en Ca y aP, 2) dieta baja Ca y aP con 500 FTU de fitasa/kg de dieta, y 3) dieta con normal contenido de Ca y aP. Resultados: en el Experimento 1, el número de nacidos totales (p<0,05), nacidos vivos (p=0,05), peso de la camada de los nacidos vivos (p<0,01), y cerdos destetados (p<0.05) se incrementó cuadráticamente con la adición creciente de fitasa hasta el nivel de 300 FTU/kg. Hubo un aumento lineal en el peso al destete de los lechones (p<0,05) al incrementar el nivel de fitasa. En el Experimento 2 no hubo diferencias en rendimiento reproductivo (p>0,10). En gestación, la ATTD del P fue menor en el grupo de bajo Ca y aP sin fitasa que en el de normal Ca y aP (p<0,01) y que en los grupos suplementados con fitasa (p<0,05). La ATTD del Ca en el grupo de normal Ca y aP fue mayor que en el grupo de bajo Ca y aP sin fitasa (p<0,05). En lactancia, el grupo suplementado con fitasa tuvo una mayor ATTD del P en comparación con los otros grupos (p<0,05), mientras que la ATTD del Ca en el grupo suplementado con fitasa fue mayor que en el de bajo Ca y aP sin fitasa (p<0,10). Cuando los datos de digestibilidad se agruparon para los períodos de gestación y lactancia, la ATTD del P en el grupo con fitasa fue la más alta entre los tratamientos dietarios (p<0,05). No hubo efecto de la fitasa en el contenido de cenizas en hueso de las cerdas al destete ni en la resistencia ósea de los lechones o en las cenizas óseas al nacimiento. Sin embargo, los lechones de las cerdas suplementadas con fitasa tuvieron al destete mayor resistencia ósea (p<0,10) y contenido de cenizas (p<0,05) en comparación con los de las cerdas no suplementadas. Hubo correlaciones positivas al nacimiento entre el peso (BW) del lechón y su resistencia ósea (p<0,01), y entre la resistencia ósea y su contenido de cenizas (p<0,001). Además, se observaron correlaciones positivas al destete entre el BW del lechón y su contenido de ceniza en hueso (p<0,05), entre el BW del lechón y su resistencia ósea (p<0,001), y entre la resistencia y el contenido de cenizas óseas (p<0,05). Conclusión: la suplementación con fitasa desde la mitad de la gestación y durante la lactancia aumentó la ATTD del P y Ca, en cerdas tanto gestantes como lactantes, mejoró la integridad ósea de la progenie, y mostró potencial para aumentar el tamaño y rendimiento de la camada, pero no afectó las características óseas de las cerdas ni de los recién nacidos.
Palabras clave: características óseas, cerdos, disponibilidad de nutrientes, enzima exógena, fósforo.
Resumo
Antecedentes: a adição de fitasse na dieta de porcas pode melhorar a digestibilidade do P e Ca, assim como a integridade óssea dos leitões. Objetivo: realizaram-se dois pesquisas para conferir o efeito da suplementação da fitasse em dietas para os períodos de gestação e lactação sobre o desempenho reprodutivo, a digestibilidade aparente no trato digestivo total (ATTD) de P, Ca e N em porcas, e as características ósseas de porcas e leitões. Métodos: o experimento 1 envolveu 204 porcas avaliadas em 3 centros de pesquisa dos Estados Unidos. As porcas foram designadas para um de quatro tratamentos no momento do serviço: 1) controle negativo [dieta baixa em Ca e P disponível (aP)], 2) Tratamento 1 mais de 300 unidades de fitasse (FTU) / kg de ração, 3) Tratamento 2, mais de 600 FTU / kg de ração, e 4) controle positivo (dieta com teor normal de Ca e aP). No Experimento 2, um total de 16 matrizes suínas (Landrace × Yorkshire, de 2,3 nascimentos em média) foram randomizados para 1 de 3 tratamentos nos dias 34-52 de gestação, com base no peso corporal (PC) e número de partos. Os tratamentos foram: 1) dieta pobre em Ca e Pd, 2) dieta pobre em Ca e aP com 500 FTU de fitasse / kg de ração, e 3) dieta com teor normal de Ca e aP. Resultados: no Experimento 1, o número de nascimentos totais (p<0,05), nascidos vivos (p=0,05), peso da leitegada de nascidos vivos (p<0,01) e leitões desmamados (p<0,05) aumentou quadraticamente com a adição crescente de fitasse no nível de 300 FTU / kg. Houve um aumento linear do peso ao desmame de leitões (p<0,05) com os níveis crescentes de fitasse. No experimento 2, não houve diferença no desempenho reprodutivo (p>0,10). Na gravidez, o ATTD de P foi menor no grupo de baixo Ca e aP sem fitasse do que no grupo Ca normal e aP (p<0,01) e nos grupos de fitasse suplementado (p<0,05). O ATTD de Ca no grupo normal de Ca e aP foi maior do que no grupo de baixo Ca e aP sem fitasse (p<0,05). Na lactação, no grupo suplementado com fitasse apresentou um ATTD de P maior em relação aos outros grupos (p<0,05), enquanto o ATTD de Ca do grupo suplementado com fitasse foi maior do que no grupo baixo de Ca e Pd e sem fitasse (p<0,10). Quando os dados de digestibilidade foram agrupados para os períodos de gestação e lactação, a ATTD de P no grupo com fitasse foi maior entre os tratamentos dietéticos (p<0,05). Não houve efeito da fitasse no teor de cinzas de osso de porcas ao desmame ou a força dos ossos ou cinzas ósseas dos leitões ao nascimento. No entanto, os leitões de porcas suplementadas com fitasse tiveram uma maior resistência óssea ao desmame (p<0,10), e cinzas (p<0,05) em comparação com as porcas não suplementadas. Houve correlação positiva entre peso (BW) ao nascimento do leitão e a resistência óssea (p<0,01) e entre a força dos ossos e cinzas (p<0,001). Além disso, foi observada uma correlação positiva entre o desmame dos leitões BW e teor de cinzas ósseas (p<0,05), entre BW dos leitões e a resistência óssea (p<0,001), e entre a resistência óssea e os conteúdos e cinza nos ossos (p<0,05). Conclusão: a suplementação com fitasse na etapa media da gestação e durante a lactação aumentou a ATTD de P e Ca, tanto em porcas gestantes quanto lactantes, melhorou muito a integridade óssea da progênie, e mostrou potencial para aumentar o tamanho e desempenho da leitegada, mas não afetou as características ósseas das porcas nem dos leitões recém nascidos.
Palavras chave: características ósseas, porcos, disponibilidade de nutrientes, enzimas exógenas, fósforo.
Introduction
Phosphorus is one of the most important nutrients, especially in gestation and lactation, to maintain normal fetal development and bone integrity and health for both sows and piglets (Trottier and Johnston, 2001). The P requirement of sows is lower in early gestation but increases in late gestation (Mahan et al., 2009), and then is raised more in lactation to supply an adequate amount of P for milk production (NRC, 2012). However, P exists in the form of phytic acid, commonly called phytate, in cereal grains and plant meals. Because phytate P is poorly available for pigs due to the lack of endogenous phytase activity, adding phytase to swine diets is necessary to release P from phytate, facilitate its absorption (Selle and Ravindran, 2008) and thereby make it available for metabolism.
Several phytase studies which have been conducted for gestating and lactating sows reported phytase supplementation clearly improved P availability in the lactation diet with less consistent effects in gestation (Kemme et al., 1997b; Sulabo et al., 2004; Nyachoti et al., 2006) and did not affect reproductive performance (Nyachoti et al., 2006). Additionally, phytase supplementation may increase milk P content in lactation (Grela et al., 2010), and high P content and phytase supplementation in the diet for nursery pigs increased available P (aP), resulting in increased bone ash content (Jones et al., 2010; Brãna et al., 2006). However, it has not been demonstrated yet that phytase supplementation to sow diets has beneficial effects on bone characteristics of their progeny.
Therefore, the objective of this study, conducted at several research stations, was to evaluate the effect of dietary phytase supplementation to gestation and lactation diets on general reproductive performance, apparent total tract digestibility (ATTD) of P and Ca of sows, and bone characteristics of their progeny.
Materials and methods
Experiment 1 and 2. Research at individual stations followed the guidelines stated in the Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching (FASS, 1999) and was conducted under protocols approved by the respective Institutional Animal Care and Use Committees.
Animals and housing condition
Experiment 1. A regional study involving 204 sows was conducted at 3 experiment stations (University of Georgia, University of Arkansas, and North Carolina State University) to evaluate phytase effects on general reproductive performance. Sows were allotted to 1 of 4 treatments at breeding. The number of sows, genetics, and facility characteristics of the stations participating in this study are presented in Table 1.
During gestation, sows were fed 1.82 kg/d during the months of March to November, and 2.27 kg/d during the months of December to February for those stations that determined that sows needed greater feed intake during that time of year. Feed was provided on an ad libitum basis during lactation. Animals were on standard deworming and vaccination schedules particular to each station. Newborn pigs were processed according to standard procedures at each station.
Experiment 2. A total of 16 sows (Yorkshire × Landrace; average parity, 2.31) were used and randomly allotted within body weight (BW) and parity groupings to the dietary treatments at d 34 to 52 of gestation after verification of pregnancy to evaluate the effects of phytase on ATTD of P and Ca as well as bone characteristics of the offspring. All sows were in Parity 1 and 2 except for one advanced parity sow on each dietary treatment (Parity 7, 7 and 8 in Treatment 1, 2 and 3, respectively). During gestation, pregnant sows were housed individually in gestation stalls (0.57 × 2.13 m2). At d 110 of gestation, all sows were moved to farrowing crates (1.52 × 2.13 m2) in an environmentally-controlled farrowing room. The crates had a plastic coated woven wire floor area with heat lamps for piglets and were equipped with a drinking nipple and feed trough for sows. Sows were fed 1.9 kg/d of a common gestation diet before being assigned to dietary treatments. Once allotted to treatments, the sows were fed 1.9 kg/d of the experimental diets for gestation until farrowing. Farrowing sows were provided 3.2 kg of the experimental diets for the first 3 days until all feed was consumed in a day. The feed allowance was then increased by 0.9 kg every 3rd day until daily feed intake reached at least 6.4 kg.
Experimental diets and treatments
Experiment 1. Corn-soybean meal (SBM) basal diets were used in both gestation and lactation, and individual stations could use either regular or dehulled SBM with appropriate adjustments. The diets were formulated to meet or exceed NRC (1998) daily requirement estimates for all nutrients (Table 2). The same source of phytase (NatuphosTM 1200; BASF Corp., Mount Olive, NJ, USA) was used at each participating station. Two supplemental levels of phytase were evaluated (300 and 600 FTU/kg of diet) in comparison with negative and positive control diets throughout gestation and lactation. The four dietary treatments were: 1) negative control (a low Ca and P diet with 0.1% less total Ca and P than recommended by NRC, 1998), 2) Treatment 1 plus 300 FTU of supplemental phytase/kg of diet, 3) Treatment 1 plus 600 FTU of supplemental phytase/kg of diet, and 4) a positive control containing recommended Ca and P levels by NRC (1998). Treatments 1, 2 and 3 contained a minimum of 0.65% Ca and 0.24% aP in gestation and 0.65% Ca and 0.22% aP in lactation. Treatment 4 contained a minimum of 0.75% Ca and 0.34% aP in gestation and 0.75% Ca and 0.32% aP in lactation. Limestone and dicalcium phosphate were adjusted to meet the assigned Ca and P levels in each treatment diet.
Experiment 2. A corn-SBM-based diet was formulated to meet or exceed NRC (1998) nutrient requirement estimates (Table 3). Treatments were: 1) low Ca and aP diet, 2) low Ca and aP diet with phytase (500 FTU/kg diet), and 3) normal Ca and aP diet. When phytase was used (Natuphos® 600G; BASF Corp), it was supplemented at the expense of corn. The contents of Ca and aP were adjusted with dicalcium phosphate and limestone, while maintaining essentially equal lysine and ME with changes in corn, SBM and choice white grease. The calculated levels of Ca and aP in the diets were 0.64% and 0.24% for the low Ca and aP groups, and 0.75% and 0.35% for the normal Ca and aP group, both in gestation and lactation. All diets contained 0.25% chromic oxide (Cr2O3) as an indigestible marker that was premixed with corn starch for determination of ATTD of Ca, P and N.
Data and sample collection
Experiment 1. Data collected included sow BW at d 110 of gestation, farrowing (within 24 h postpartum), and weaning. The number of pigs at birth (total and alive), and weaning were recorded as well as BW at birth, and weaning. Overall lactation daily feed intake was recorded. After weaning, days to estrus postweaning was recorded.
Experiment 2. The objective of this experiment was primarily to evaluate the effect of phytase supplementation in the gestation and lactation diets on the ATTD of Ca and P of sows and bone characteristics of their progeny. All sows were weighed at d 110 of gestation, farrowing and weaning, and piglet BW was also recorded at birth and weaning. Overall lactation daily feed intake and the numbers of pigs at birth (total and alive), and weaning were recorded.
The fecal collection periods were scheduled in gestation (d 60, 70, 80 and 90) and lactation (d 6 and 16). Fecal collection was conducted in the morning at each collection day and the collected samples were stored at -20 °C until processing. The fecal samples were thawed, dried in a forced-air drying oven at 55 °C for 1 week, and ground through a 1 mm screen using a Wiley Laboratory Mill (Model 3, Arthur H. Thomas Co., PA, USA) for laboratory analysis. All ground feces were put in a single bag for each sow and collection day, and then stored at 4 °C until analysis.
To investigate bone characteristics, two piglets/ litter were sacrificed at birth before suckling and at weaning. At weaning, all sows were moved to the abattoir and killed to collect bone samples. The 3rd and 4th metacarpals of each foot of the sows were collected to analyze bone ash contents and the femurs from piglets were collected to measure bone strength and ash content. Collected bone samples were autoclaved at 120 °C for 3 minutes and soft tissues were removed, and then bones were stored frozen in plastic bags.
Chemical analysis
In Experiment 2, P was assessed by a gravimetric method (modification of method 968.08; AOAC, 1990). Calcium was assessed by flame atomic absorption spectrophotometry (AAS; Thermoelemental, SOLAAR M5, Thermo Electron Corp., Verona, WI, USA) according to a modification of the AOAC (1995) procedure (method 927.02). Nitrogen was measured using Dumas methodology in an automatic N analyzer (model FP-2000, LECO Corp., Saint Joseph, MI, USA). Chromium concentration was assessed by a modification of the method described by Williams et al. (1962) with flame AAS.
To assess bone strength, bones were thawed and broken using an Instron machine (Model TM 1123; Instron Corp., Canton, MA, USA). To assess bone ash, after obtaining bone strength, bones were dried overnight at 105 °C. Bones were ashed at 600 °C in a muffle furnace overnight and total ash percentage was measured.
Calculations and statistical analysis
Experiment 1. Post-study evaluation revealed a parity imbalance across treatments. Thus, results from sows greater than actual Parity 6 at study initiation were removed from the data analysis. This reduced the data set from 411 litters from 204 sows to 365 litters from 182 sows. The data were subjected to ANOVA using the GLM procedure of SAS (SAS inst. Inc., Cary, NC, USA) as a completely randomized design, with the sow or litter serving as the experimental unit. The model included terms for station, treatment, parity on study (or actual parity in a preliminary analysis), and all possible interactions. Linear and quadratic contrast coefficients were used to evaluate treatment effects. Because of unbalanced data observation by station and treatment, least squares means are presented and are separated by using the PDIFF option of SAS.
Experiment 2. The ATTD of P, Ca, and N were calculated by the indicator method, using Cr contents in the diets and fecal samples. The formula to calculate ATTD was:
Apparent total tract digestibility (%) = [1 − (Crdiet/Crdigesta) × (Nutrdigesta/Nutrdiet)] × 100
where, Crdiet is the Cr concentration in the diet, Crdigesta is the Cr concentration in the feces, Nutrdigesta is a nutrient content in the feces, and Nutrdiet is a nutrient content in the diet.
The ATTD for each nutrient was calculated for each collection day and was subsequently pooled for a single gestation and lactation values per sow and were also pooled to evaluate the overall treatment effects.
Because of different weaning ages across litters, the piglet BW at weaning was normalized by the following equation:
Weaning weight = BW of born alive + [(BW at weaning – BW of born alive)/weaning age] × 21
Statistical data-analyses were performed on the reproductive performance, ATTD of P, Ca and N, and bone characteristics. The experimental data were analyzed by ANOVA as a completely randomized design with the GLM procedure of SAS. Nonorthogonal contrasts were conducted to compare the 3 dietary treatments. The individual sow or litter was considered as the experimental unit for the reproductive performance, ATTD values, and bone ash of sows, and the individual piglet was considered as the experimental unit for bone strength and bone ash content of the piglets. For the ATTD values in gestation and lactation the model included the collection day, diet, and diet × collection day interaction. Following the establishment of no diet × collection day interactions (p>0.10), the data were pooled within period (gestation and lactation) and the model included the period, diet, and diet × period interaction for overall ATTD values. For reproductive performance and bone characteristics, the diet effect was the only term used in the statistical model. In the bone measurements for piglets, BW for each sampling time was considered as a covariate. Least squares mean separations were accomplished by using the PDIFF option of SAS. Correlation coefficient analysis was performed with Proc Corr of SAS to determine correlations among piglet BW, bone strength and ash content at birth and weaning. An alpha level of 0.05 was used in Experiment 1 for declaration of significance but, because of limited experimental units in Experiment 2, an alpha level of 0.10 was used for declaration of significance.
Results
Experiment 1
The results of sow and litter responses by station in the cooperative research studies are presented in Table 4. Not all stations were able to collect data for all response measurements. As expected, there were several individual station effects on litter size and litter performance.
The results of phytase supplementation on sow and litter performance are presented in Table 5. There were many main effects of test parity (sows could remain on the study for as many as 3 parities), but these were normal responses in performance or BW associated with advancing parity and, because there were no parity × diet interactions, these results are not reported.
There were no significant dietary differences in sow weight measures. However, quadratic responses were detected in the number of total (p<0.05) and live born (p=0.05), and litter weight of live born (p<0.01) and weaning pigs (p<0.05) with the highest values in the 300 FTU/kg phytase supplementation group. A linear increase was observed in piglet weaning weight with increasing phytase supplementation levels (p<0.05).
Experiment 2
All sows and piglets were in good health and condition during the experimental period. Two sows were removed from the gestation data set because one sow was not pregnant and the other sow had only 2 pigs at farrowing which was considered an abnormal litter size. Additionally, a sow from each of Treatment 2 and 3 was absent from the lactation data set because of sudden death at d 100 and 112 of gestation, respectively.
In the results of sow and litter performance, phytase supplementation to gestation and lactation diets had no influences on any response measures of litter size, lactation daily feed intake, and BW of sows and piglets during lactation (p>0.10; Table 6). While, this was not unexpected because there were limited observations and piglets were slaughtered at birth and weaning, which further hinders examination of those measures, the means are provided for informational purposes when examining results that are presented in Tables 7 and 8.
In gestation, the ATTD of P in the low Ca and aP without phytase group was lower than that in the normal Ca and aP (p<0.01) and phytase-supplemented (p<0.05) groups while the phytase-supplemented group had similar ATTD of P compared to the normal Ca and aP group (p=0.37; Table 7). For the ATTD of Ca, the value in the normal Ca and aP group was similar to that in the phytase-supplemented group (p=0.31) but higher than that in the low Ca and aP without phytase group (p<0.05). In lactation, sows fed the low Ca and aP diet with phytase had higher ATTD of P compared with those fed the normal Ca and aP (p<0.05) or the low Ca and aP without phytase diet (p<0.05) whereas the ATTD of Ca in the phytasesupplemented group was only higher than that in the low Ca and aP without phytase group (p<0.10). There were no differences for ATTD of N in gestation and lactation among the dietary treatments (p>0.19).
When the digestibility data were averaged for both gestation and lactation periods (the mean value), the ATTD of P in the phytase-supplemented group was the highest (p<0.05) and that in the low Ca and aP without phytase group was the lowest (p<0.05) among dietary treatments. For the ATTD of Ca, the value in the low Ca and aP without phytase group was lower than those in the normal Ca and aP (p<0.05) and phytase-supplemented (p<0.05) groups, and the phytase-supplemented group did not differ from the ATTD of Ca for the normal Ca and aP group (p=0.71). Evaluating the period effect of gestation and lactation, the ATTD of all nutrients measured were enhanced in lactation compared with those in gestation (p<0.01). An interaction between treatment and period was observed for the ATTD of P (p<0.05) wherein the response to added phytase was greater in lactation than in gestation.
There were no differences in sow bone ash content at weaning, and piglet bone strength and ash content at birth (p>0.12; Table 8). Comparing piglet bone characteristics at weaning between phytase-supplemented and nonphytase groups, phytase supplementation of the sow diet increased piglet bone ash content (47.6 vs. 45.1%, p<0.05) and bone strength (125.1 vs. 90.8 kg, p<0.10). Differences remained after piglet BW was considered as a covariate (47.5 vs. 45.4%, p<0.05 for bone ash content; 118.7 vs 98.5 kg, p<0.10 for bone strength).
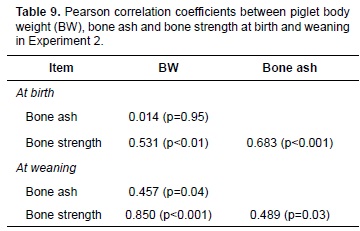
In the results of the correlation analysis, there were positive correlations at birth between piglet BW and bone strength (r=0.53; p<0.01) and between bone strength and ash content (r=0.68; p<0.001, Table 9). Additionally, at weaning there were positive correlations between piglet BW and bone ash content (r=0.46; p<0.05), between piglet BW and bone strength (r=0.85; p<0.001), and between bone strength and ash content (r=0.49; p<0.05).
Discussion
Aaron and Hays (2001) stated that progress in sow nutrition and management research is impeded by large variation among sows in the economically important reproductive traits. With normal variation, the number of replications needed to detect a 10% difference in litter size at birth with an 80% chance of detecting that difference and a 10% probability level is 112 sows per treatment. Although none of the 3 participating stations could dedicate that many sows to the experiment individually, about 90 litters per treatment are collectively reported herein, and those numbers were adequate to detect differences in several responses.
The relative lack of differences in sow BW and litter size among all dietary treatments in both studies mean that the reduced Ca and P contents in gestation and lactation diets had no detrimental effect on reproductive performance, which agrees with Nyachoti et al. (2006) who reported that low levels of Ca and P in the gestation and lactation diets with phytase had no negative effects on sow BW and piglet growth during the nursing period. In Experiment 1 there was a quadratic effect of phytase supplementation on litter size. The lower level of phytase supplementation inexplicably increased litter size and weight at birth, and litter weight at weaning while those responses were diminished when sows were fed the highest level of phytase. Additionally, piglet weaning weight was linearly increased by increasing level of phytase supplementation. Grela et al. (2010) reported higher numbers of live born and weaned pigs with phytase supplementation (500 FTU/kg with low P content) and higher litter weight at birth and weaning in the high P or phytase supplementation groups in comparison with a low P group. In agreement with these recent data, the results of the present study suggest that phytase supplementation to low P diets could improve litter growth. However, there is limited information to explain the reason why quadratic responses appeared in litter size and performance by phytase supplementation levels. Nevertheless, the number of pigs at weaning did not differ from the normal Ca and aP diet. Lyberg et al. (2007) reported higher numbers of piglets born from sows fed low P diets supplemented with phytase compared with sows fed a high P diet but it was not maintained until weaning because of a negative effect on piglet mortality. In contrast, Nyachoti et al. (2006) reported sow and litter performance was not affected by phytase supplementation. Therefore, further study is needed to clarify the phytase effect on litter size and performance and possible modes of action (e.g. increase of P content in colostrum and milk) in addition to simply supplying additional P.
In Experiment 2, as with Experiment 1, there were no significant differences on reproductive performance including BW of sows and piglets and litter size at birth and weaning. As stated, this was not unexpected because there were limited observations and piglets were slaughtered at birth and weaning. But the primary objectives of Experiment 2 were not normal production measures but ATTD of Ca and P and offspring bone characteristics.
In Experiment 2 phytase supplementation improved the ATTD of P both for gestating and lactating sows. These results agree with many previous studies that report phytase supplementation increases P digestibility and availability in gestating and lactating sows (Kemme et al., 1997a, b; Baidoo et al., 2003; Jongbloed et al., 2004; Nyachoti et al., 2006; Männer and Simon, 2006; Hanczakowska et al., 2009; Grela et al., 2011). The low Ca and aP diet without phytase reduced the ATTD of Ca and P in gestation and phytase supplementation to this diet then improved ATTD to levels similar to that in the normal Ca and aP group, which agrees with previous studies (Jongbloed et al., 2004; Hanczakowska et al., 2009). Additionally, in lactation the ATTD of Ca was increased by phytase supplementation compared with the non-phytase group. The possible mode of action whereby Ca digestibility improves by supplementing phytase could be that when phytase hydrolyzes phytates, there is limitation of Ca-phytate complex formation in the small intestine that blocks intestinal Ca absorption (Selle et al., 2009). In addition, already phytate-bound minerals may be liberated and become available for absorption from the intestine (Kemme et al., 1999). Jongbloed et al. (2004) observed an improvement of Ca digestibility in gestation and lactation by phytase supplementation and Grela et al. (2011) also demonstrated the same effect in lactating sows. Nyachoti et al. (2006) reported Ca digestibility in gestation was slightly improved when sows were fed phytase-supplemented diets, but more so in lactation. Therefore, supplementing phytase to sow diets appears to improve the ATTD of Ca as well as that of P.
The interaction between treatment and period observed for the ATTD of P was due to a difference in magnitude of the improvements in P digestibility of gestating and lactating sows by phytase supplementation and not to a difference in the direction of the response. In gestation, phytase supplementation improved the ATTD of P to levels similar to the normal Ca and P group whereas a more pronounced effect of phytase was observed in lactation. In the ATTD of Ca, the mean value was improved by phytase supplementation similar to that in the normal Ca and P group which, as with the ATTD of P, was associated with a slight improvement in gestation and greater improvement in lactation. Across studies, the phytase effect on P digestibility in gestation is conflicting. Several studies reported that gestating sows had an increased P digestibility when fed phytase in the low P diets (Jongbloed et al., 2004; Männer and Simon, 2006; Hanczakowska et al., 2009) whereas Nyachoti et al. (2006) reported phytase supplementation with 500 FTU/kg in a corn-SBM based diet with low P level showed a positive effect on P digestibility in lactation but not in gestation. Kemme et al. (1997b) also reported that even though mean gestation P digestibility was increased by supplementing phytase at 500 FTU/kg of diet, no improvement in P digestibility from phytase addition was observed in the mid-gestation period (d 60 of gestation). While phytase supplementation levels may have varied among the reported studies, it should be noted that P levels of the low P diets used in the studies which showed no effect of phytase on P digestibility for gestating sows (0.48% total P; Kemme et al., 1997b, Nyachoti et al., 2006) were relatively higher than those in the other studies which reported the improvement of P digestibility in gestating sows from phytase supplementation (0.36 to 0.39% total P; Jongbloed et al., 2004; Männer and Simon, 2006; Hanczakowska et al., 2009). This indicates that the inconsistent effect of dietary phytase supplementation in gestating sows that is reported might be attributed to different levels of aP for sows from the diet. Regarding the reproductive period effect, higher digestibilities of P, Ca, and N were detected in lactation compared to gestation. This observation is in full agreement with previous research (Kemme et al., 1997b; Sulabo et al., 2004; Jongbloed et al., 2004). Kemme et al. (1997b) demonstrated lactating sows had higher apparent digestibilities of Ca and P than gestating sows. Similarly, Sulabo et al. (2004) reported that the ATTD of P was higher in lactation than in gestation by as much as 14.7%. Theoretically, P released from the diet containing phytase could provide sufficient amounts of aP for gestating sows close to or above their P requirement (Kemme et al., 1997b). However, lactating sows demand more nutrients to produce milk, resulting in increased Ca and P requirements (NRC, 2012). In this study it could be assumed that lactating sows consumed more aP and phytate P than gestating sows because of ad libitum feed allowance and approximately 2 times higher proportion of SBM in the lactation diet than in the gestation diet. Therefore, more pronounced effect of phytase in lactating sows than gestating sows may be likely due to both higher consumption of available nutrients (i.e. P) as well as substrates for phytase in lactation than in gestation.
In the results of bone characteristics, no phytase effects on bone strength and ash content of piglets at birth were observed, even though the ATTD of P was increased in the gestation period. Mahan and Vallet (1997) suggested that the contents of Ca and P in fetus and milk were not affected by inadequate levels of these elements in sow diets. Additionally, the bone ash content of newborn pigs was not influenced by dietary Ca and P levels under or above the requirements for gestating sows when assessed by Mahan and Fetter (1982). Therefore, in the current study the lack of an improvement in bone integrity of newborn pigs in spite of increased P absorption in gestating sows is not surprising. At weaning, while no differences were detected in sow bone ash content, piglet bone strength and ash content showed clear improvements related to phytase supplementation of the sows. Previous research has reported that phytase supplementation both in gestation and lactation increased P level in milk as well as colostrum (Grela et al., 2010; Czech et al., 2011). Even though the nutrient content of milk was not evaluated in this experiment, it could be hypothesized that the increased P digestibility from phytase supplementation positively affected milk P level and thereby bone integrity of the progeny was improved.
There were several positive correlations among piglet BW, bone strength and ash content at birth and weaning. Piglet bone strength was correlated with BW regardless of sampling times. In the case of bone ash content, there were correlations with bone strength both at birth and weaning and with piglet BW at weaning. Crenshaw et al. (1981) reported that an increase in bone mineralization led to an increased maximum stress and bending moment of bone. When the bone mineral density was high, not only ash, Ca and P concentration of the bone were increased, but also bone strength increased (Nielsen et al., 2007). These previous studies provide the possible explanations for the correlations in this study where higher bone ash contents led to increased bone strength.
In conclusion, phytase supplementation increased the ATTD of Ca and P both for gestating and lactating sows, with more pronounced effects in lactation than in gestation, and has the potential to improve litter performance. Although there were no significant effects on bone characteristics of piglets at birth, adding phytase to sow diets in gestation and lactation improved the bone integrity of their progeny at weaning, which could result in subsequent improvement of pig bone status.
Notes
¤ To cite this article: Jang YD, Lindemann MD, van Heugten E, Jones RD, Kim BG, Maxwell CV, Radcliffe JS. Effects of phytase supplementation on reproductive performance, apparent total tract digestibility of Ca and P and bone characteristics in gestating and lactating sows. Rev Colomb Cienc Pecu 2014; 27:178-193.
¶ This manuscript is published by the Kentucky Agricultural Experiment Station as paper number 13-07-055.
References
Aaron DK, Hays VW. Statistical techniques for the design and analysis of swine nutrition experiments. In: Lewis AJ, Southern LL, editors. Swine Nutrition. 2nd ed. Washington D.C.: CRC Press; 2001. p. 605-622. [ Links ]
AOAC, Association of Official Analytical Chemists. Official Methods of Analysis. 15th ed. Arlington, VA; 1990. [ Links ]
AOAC, Association of Official Analytical Chemists. Official Methods of Analysis. 16th ed. Arlington, VA; 1995. [ Links ]
Baidoo SK, Yang QM, Walker RD. Effects of phytase on apparent digestibility of organic phosphorus and nutrients in maize-soya bean meal based diets for sows. Anim Feed Sci Technol 2003; 104:133-141. [ Links ]
Brãna DV, Ellis M, Castañeda EO, Sands JS, Baker DH. Effect of a novel phytase on growth performance, bone ash, and mineral digestibility in nursery and grower-finisher pigs. J Anim Sci 2006; 84:1839-1849. [ Links ]
Crenshaw TD, Peo ER Jr, Lewis AJ, Moser BD. Bone strength as a trait for assessing mineralization in swine: a critical review of techniques involved. J Anim Sci 1981; 53:827-835. [ Links ]
Czech A, Stachyra K, Woznica A. Influence of feed enzymes on the content of mineral elements in sows milk and pigs blood plasma. Ann UMCS, Zootechnica sec EE 2011; 29:22-32. [ Links ]
FASS, Guide for the Care and Use of Agricultural Animals in Research and Teaching. Federation of Animal Science Societies. 1st rev. ed. Champaign, IL; 1999. [ Links ]
Grela ER, Matras J, Czech A, Krasucki W. The influence of microbial phytase supplementation to diets with high or low native phytase activity on sow reproductive traits and composition of colostrum and milk. J Anim Feed Sci 2010; 19:418-429. [ Links ]
Grela ER, Matras J, Czech A. Effects of supplemental phytase on nutrient digestibility and performance of sows fed diets with high or low native phytase activity. Czech J Anim Sci 2011; 56:443-450. [ Links ]
Hanczakowska E, Swiatkiewicz M, Kühn I. Effect of microbial phytase supplement to feed for sows on apparent digestibility of P, Ca and crude protein and reproductive parameters in two consecutive reproduction cycles. Medycyna Wet 2009; 65:250-254. [ Links ]
Jones CK, Tokach MD, Dritz SS, Ratliff BW, Horn NL, Goodband RD, DeRouchey JM, Sulabo RC, Nelssen JL. Efficacy of different commercial phytase enzymes and development of an available phosphorus release curve for Escherichia coli-derived phytases in nursery pigs. J Anim Sci 2010; 88:3631-3644. [ Links ]
Jongbloed AW, van Diepen JThM, Kemme PA, Broz J. Efficacy of microbial phytase on mineral digestibility in diets for gestating and lactating sows. Livest Prod Sci 2004; 91:143-155. [ Links ]
Kemme PA, Radcliffe JS, Jongbloed AW, Mroz Z. The effects of sow parity on digestibility of proximate components and minerals during lactation as influenced by diet and microbial phytase supplementation. J Anim Sci 1997a; 75:2147-2153. [ Links ]
Kemme PA, Jongbloed AW, Mroz Z, Beynen AC. The efficacy of Aspergillus niger phytase in rendering phytate phosphorus available for absorption in pigs is influenced by pig physiological status. J Anim Sci 1997b; 75:2129-2138. [ Links ]
Kemme PA, Jongbloed AW, Mroz Z, Kogut J, Beynen AC. Digestibility of nutrients in growing–finishing pigs is affected by Aspergillus niger phytase, phytate and lactic acid levels 2. Apparent total tract digestibility of phosphorus, calcium and magnesium and ileal degradation of phytic acid. Livest Prod Sci 1999; 58:119-127. [ Links ]
Lyberg K, Andersson HK, Simonsson A, Lindberg JE. Influence of different phosphorus levels and phytase supplementation in gestation diets on sow performance. J Anim Physiol Anim Nutr 2007; 91:304-311. [ Links ]
Mahan DC, Fetter AW. Dietary calcium and phosphorus levels for reproducing sows. J Anim Sci 1982; 54:285-291. [ Links ]
Mahan DC, Vallet JL. Vitamin and mineral transfer during fetal development and the early postnatal period in pigs. J Anim Sci 1997; 75:2731-2738. [ Links ]
Mahan DC, Watts MR, St-Pierre N. Macro- and micromineral composition of fetal pigs and their accretion rates during fetal development. J Anim Sci 2009; 87:2823-2832. [ Links ]
Männer K, Simon O. Effectiveness of microbial phytases in diets of sows during gestation and lactation. J Anim Feed Sci 2006; 15:199-211. [ Links ]
Nielsen DH, McEvoy FJ, Madsen MT, Jensen JB, Svalastoga E. Relationship between bone strength and dual-energy X-ray absorptiometry measurements in pigs. J Anim Sci 2007; 85:667-672. [ Links ]
NRC, National Research Council. Nutrient Requirements of Swine. 10th rev. ed. Washington D.C.: National Academic Press; 1998. [ Links ]
NRC, National Research Council. Nutrient Requirements of Swine. 11th rev. ed. Washington D.C.: National Academic Press; 2012. [ Links ]
Nyachoti CM, Sands JS, Connor ML, Adeola O. Effect of supplementing phytase to corn- or wheat-based gestation and lactation diets on nutrient digestibility and sow and litter performance. Can J Anim Sci 2006; 86:501-510. [ Links ]
Selle PH, Ravindran V. Phytate-degrading enzymes in pig nutrition. Livest Sci 2008; 113:99-122. [ Links ]
Selle PH, Cowieson AJ, Ravindran V. Consequences of calcium interactions with phytate and phytase for poultry and pigs. Livest Sci 2009; 124:126-141. [ Links ]
Sulabo RC, Thaler RC, Stein HH. Effect of body weight and reproductive status on phosphorus digestibility and efficacy of phytase in pigs. J Anim Sci 2004; 82 Suppl 2: 215. [ Links ]
Trottier NL, Johnston LJ. Feeding gilts during development and sows during gestation and lactation. In: Lewis AJ, Southern LL, editors. Swine Nutrition. 2nd ed. Washington D.C.: CRC Press; 2001. p. 725-769. [ Links ]
Williams CH, David DJ, Iismaa O. The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry. J Agric Sci 1962; 59:381-385. [ Links ]
