











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Genetic variability of livestock populations is fading. This is due, in part, to genetic improvement programs (FAO, 1998; Gutiérrez and Goyache, 2005). Boichard et al. (1997) mentioned that change in genetic variability across generations could be described by genealogical characterization; and the conservation and consanguinity status of populations can be monitored (Vicente et al., 2012; Pienaar et al., 2015). Genealogical analysis requires the creation and analysis of sorted databases (Gutiérrez and Goyache, 2005); it has been widely used in equines (Vicente et al., 2012; Roos et al., 2015), and canines (Głażewska, 2008; Leroy, 2011). The most important population descriptors are based on the gene probability origin and include the effective number of founders (fe) and ancestors (fa), effective population size (Ne), genetic integrity index (PEC), inbreeding coefficient (F), average relatedness (AR), genetic conservation index (GCI), and generation intervals (GI) (Lacy, 1989; Boichard et al., 1997; Gutiérrez et al., 2003).
Tropical Milking cattle of Mexico (LT) is a criollo breed naturalized to hot climates (de Alba, 2011). Due to the small size of its current population -less than 1,000 pure heads (AMCROLET, 2015)- LT is considered endangered (FAO, 2013; 2015). The LT original population developed from cattle brought to the Americas by the Spaniards during the XV century. The current LT population originates from 17 cows and two bulls from Central America brought to Mexico in 1965 (de Alba, 2011). The original herd was disbanded in 2000, from which the initial selection nucleus of 61 females, 5 males and AI straws from 15 bulls was shaped. Currently, this nucleus is the only source of breeding stock with a recognized genetic improvement program with evaluations for milk production (Santellano-Estrada et al., 2011). A review has been published on LT (Rosendo-Ponce and Becerril-Pérez, 2015). The LT population was divided in six families in a mating scheme of sire rotation to avoid the detrimental effects of inbreeding (Yamada, 1981). The LT has genealogical and production records collected for over 65 years. The Mexican Romosinuano and the Tropical Milking Breeder Association (AMCROLET) is responsible for the LT breed Herd Book. If necessary studies and precautionary measures are not taken, small criollo populations can lose genetic variability, increase inbreeding and become extinct. The aim of this study was to determine the genetic variability and inbreeding levels of the Tropical Milking criollo cattle of Mexico.
Materials and Methods
A total of 3,427 LT genealogical records between years 1945 and 2013 registered in the AMCROLET´s Herd Book (HB) were used. The database was constructed with the individual animal identification, parents, birthdate and sex. Animals with offspring not used for breeding were discarded. Two populations were defined from the HB: i) 3,427 of the LT breed population (PLT), and ii) 608 animals born between 1950 and 2013, ancestors of the current selection nucleus (PCP). Analysis of gene origin is useful for divided populations to assess the selection effects in small populations (Boichard et al., 1997). Animals that did not have at least one known parent were considered founders. Estimates of genetic variability parameters were calculated with the ENDOG v4.8 program (Gutiérrez et al., 2010) as described.
Animals with unknown genealog are fe. It is assumed they are the ancestors´ parents and the origin of the current genetic variability in the population; calculated as 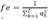 where the sum of qk corresponds to the gene proportion of the offspring population provided by each founder (k), and f is the total of alleged founders in the population genealogy. When each founder has the same contribution (1/f), population variability is in equilibrium (Boichard et al., 1996). The minimum of ancestors is fa (founders or not), which provided genes into the population and explain all the variability. It is calculated as
where the sum of qk corresponds to the gene proportion of the offspring population provided by each founder (k), and f is the total of alleged founders in the population genealogy. When each founder has the same contribution (1/f), population variability is in equilibrium (Boichard et al., 1996). The minimum of ancestors is fa (founders or not), which provided genes into the population and explain all the variability. It is calculated as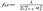 the sum of
the sum of 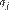 corresponds to the marginal genetic contribution of the ancestor j, which is not explained by other ancestors that were previously chosen, and is the total alleged ancestors in the genealogy.
corresponds to the marginal genetic contribution of the ancestor j, which is not explained by other ancestors that were previously chosen, and is the total alleged ancestors in the genealogy.
The existing number of effective breeding individuals giving rise to the inbreeding rate is Ne (Falconer and Mackay, 1996). It is estimated by the inbreeding increase between two consecutive generations (ΔF) in the form Ne= ½ΔF. The genealogy integrity is the amount of genealogical information of each individual, which can be evaluated with the PECind = 4Cmales × Cfemales ⁄ Cmales + Cfemales (MacCluer et al., 1983), where the paternal and maternal line contributions are Cmales and Cfemales, respectively, calculated  , where a is the proportion of current ancestors in the generation i, and d is total generations. The genealogy depth of each individual is determined by the number of complete generations (CG), which indicates the farthest generation in which all the ancestors are known, the maximum number of generations (MG) between the individual and its oldest known ancestor, and the number of equivalent full generations (EG) calculated by the sum of the coefficients (1/2)n, where n is the number of generations separating the individual from each known ancestor (Goyache et al., 2003).
, where a is the proportion of current ancestors in the generation i, and d is total generations. The genealogy depth of each individual is determined by the number of complete generations (CG), which indicates the farthest generation in which all the ancestors are known, the maximum number of generations (MG) between the individual and its oldest known ancestor, and the number of equivalent full generations (EG) calculated by the sum of the coefficients (1/2)n, where n is the number of generations separating the individual from each known ancestor (Goyache et al., 2003).
The probability that two haplotypes at any locus sampled at random from all loci in the genome are identical by descendant is F (Curik et al., 2014). The increase of inbreeding (ΔF) per generation is calculated as 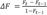 where Ft and Ft-1 are the average inbreeding in generation t, and the previous. The coefficient average in the corresponding row of the additive relationship matrix (A) for each individual is AR (Gutiérrez and Goyache, 2005). The allele percentage of the current population coming from the initial one is GCI, which is calculated as
where Ft and Ft-1 are the average inbreeding in generation t, and the previous. The coefficient average in the corresponding row of the additive relationship matrix (A) for each individual is AR (Gutiérrez and Goyache, 2005). The allele percentage of the current population coming from the initial one is GCI, which is calculated as 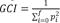 , where pi is the founder´s gene proportion, i in one animal’s genealogy. The time elapsed to replace one generation with the next one is GI.
, where pi is the founder´s gene proportion, i in one animal’s genealogy. The time elapsed to replace one generation with the next one is GI.
Results
The animals that made up the databases are shown in Table 1. In the PLT 458 bulls and 2,969 cows were registered, and 59.8 and 50.1% of these animals were breeding animals. The PCP genealogy had 219 bulls and 389 cows, of which 60.2 and 78.1% were chosen to mate. The PLT had 41.2% of the animals with both parents unknown, while in the PCP only 13.3%, while animals with offspring accounted for 51.4 and 71.7% for each population. The number of offspring per sire in the PCP was reduced to more than half of the PLT, and the number of cows increased by almost 12%. The sire/dam ratio, and the maximum number of offspring per sire, and per dam were 43.4%, and 23 and 5 in the PLT, and 18.3%, and 65 and 8 in the PCP.
Table 1 Number of animals included in the databases of all registered animals (PLT), and in the selection nucleus (PCP) of the Tropical Milking Criollo cattle.
| Animals | PLT | PCP |
|---|---|---|
| Database | 3,427 | 608 |
| Both parents known | 2,013 | 461 |
| Both parents unknown | 1,055 | 81 |
| Sire unknown | 230 | 42 |
| Dam unknown | 129 | 24 |
| Base population (animals with at least one parent unknown) | 147 | 147 |
| Males with progeny | 274 | 132 |
| Females with progeny | 1,489 | 304 |
| Offspring per sire | 7.79 | 3.73 |
| Offspring per dam | 1.51 | 1.69 |
The gene origin parameters were estimated to determine genetic variability (Table 2). The founders (f) and alleged ancestors (a) in the PLT were lower than 900, of which only fe/f= 12.5 and fa/a= 8.4% were effective, in the PCP f and a barely exceeded 100, although still in reduced proportion, and fe/f= 37.7 and fa/a= 25.5%, three times bigger than in the PLT. The small fe indicates more frequent use of some sires, one of them had 46 descendants. The fe was reduced in the PCP (38%) compared to the PLT. The fa/fe indexes calculated were 0.64 and 0.60 in PLT and PCP.
Table 2 Descriptors of gene origin in all registered animals (PLT) and in the selection nucleus (PCP) of Tropical Milking Criollo cattle.
| Parameter | PLT | PCP |
|---|---|---|
| Founders assumed (f) | 890 | 114 |
| Effective number of founders (fe) | 111 | 43 |
| Ancestors assumed (a) | 855 | 102 |
| Effective number of ancestors (fa) | 72 | 26 |
| Effective population size (Ne) | 68.1 | 64.6 |
| Inbreeding coefficient (F, %) | 1.07 | 2.14 |
| Average relatedness (AR, %) | 1.19 | 5.55 |
The Ne of PLT was similar to that of PCP and both exceeded the 50 threshold. Approximately 70% of the PLT had limited genealogical information and appears in the categories under 0.4 of PEC; only 10% occurred in the category above 0.8. All the categories of the inbreeding coefficient F had wide ranges, and tended to increase as the PEC category did. The estimated F of 4.32% of the highest PEC category is the most appropriate estimate of homozygosity in the PLT. Although only 22.5% of the animals belonged to the top two categories of 0.60 of PEC, more than 80% of inbred animals occurred there, in the PLT 22.5% of the animals showed some consanguinity (Table 3).
In the PCP, the increased occurrence of almost 40% of the animals was presented in the superior category of 0.8 of PEC, which had an F of 3.48% (Table 4).
Table 3 Estimated inbreeding coefficient (F) by genealogy integrity index classes (PEC) of all registered animals (PLT) of Tropical Milking Criollo cattle.
| PEC | Animals | F (%) | Animals with F (%) > 0 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Interval | Mean | No. | % | Interval | Mean | Interval | No. | % | |
| 0.000 - 0.239 | 0.02 | 1520 | 44.4 | 0.0 -16.80 | 0.04 | 0.31 - 16.80 | 17 | 2.2 | |
| 0.240 - 0.399 | 0.31 | 850 | 24.8 | 0.0 - 27.12 | 0.26 | 0.07 - 27.12 | 32 | 4.2 | |
| 0.400 - 0.599 | 0.52 | 287 | 8.4 | 0.0 - 25.00 | 2.32 | 0.05 - 25.00 | 75 | 9.7 | |
| 0.600 - 0.799 | 0.70 | 427 | 12.5 | 0.0 - 37.50 | 2.96 | 0.02 - 37.50 | 326 | 42.2 | |
| 0.800 - 1.000 | 0.88 | 343 | 10.0 | 0.0 - 29.49 | 4.32 | 0.07 - 29.49 | 322 | 41.7 | |
| Total | 0.49 | 3427 | 100.1 | 772 | 100.0 | ||||
Table 4 Estimated inbreeding coefficients (F) by genealogy integrity index classes (PEC) of the selection nucleus (PCP) of Tropical Milking Criollo cattle.
| PEC | Animals | F (%) | Animals with F (%) > 0 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Interval | Mean | No. | % | Interval | Mean | Interval | No. | % | |
| 0.000 - 0.239 | 0.02 | 159 | 26.2 | 0.0 - 12.82 | 1.15 | 0.04 - 12.82 | 76 | 22.8 | |
| 0.240 - 0.399 | 0.30 | 59 | 9.7 | 0.0 - 6.38 | 0.78 | 0.53 - 6.38 | 20 | 6.0 | |
| 0.400 - 0.599 | 0.52 | 45 | 7.4 | 0.0 - 8.82 | 0.67 | 0.92 - 8.82 | 11 | 3.3 | |
| 0.600 - 0.799 | 0.71 | 107 | 17.6 | 0.0 - 15.88 | 1.88 | 0.07 - 15.88 | 52 | 15.6 | |
| 0.800 - 1.000 | 0.92 | 238 | 39.1 | 0.0 - 26.09 | 3.48 | 0.20 - 26.09 | 175 | 52.4 | |
| Total | 0.56 | 608 | 100.0 | 334 | 100.1 | ||||
However, 55% of the animals are considered with some consanguinity; out of these, more than 50% occurred in the top category of PEC. In the PCP, the percentage of animals in the two lowest categories of PEC was medium (36%). The F and AR estimated in the whole populations were less than 1.2 in PLT, which doubled and quadrupled in PCP (Table 2). Emphasis should be placed on avoiding matings between closely related animals, as the PCP population is small and has high AR.
For genealogy depth, the PLT was incomplete in all generations, with CG= 0.95, MG= 4.64 and EG= 2.00, with an F increase of 0.24% between an individual and its farthest known ancestor by generation. The two recent generations and 68 animals described the current PLT variability, the increase in consanguinity between these generations was 0.73%. The genealogy depth of PCP was better with CG= 1.82, MG= 7.04 and EG= 3.53, and an inbreeding increase of 0.23% between an individual and its farthest known ancestor; the number of generations that explained the genetic variability was 3.5 with an average increase of inbreeding of 0.77% per generation.
Over time, the GCI appears to be related to F changes (Figure 1). In the PLT and PCP there are similar trends between GCI and F; however, both estimators in the PCP showed a tendency to decrease. The estimated GCI of the PCP with PEC higher than 0.8 was 3.08%, indicating a reduction in the gene conservation of the original population.
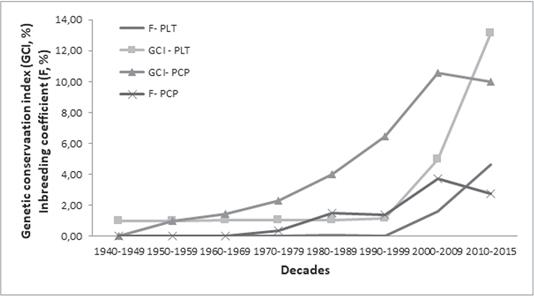
Figure 1 Genetic conservation index (GCI, %) and inbreeding coefficient (F, %) through decades in all registered animals (PLT), and in the selection nucleus (PCP) of Tropical Milking Criollo cattle.
The GIs were shorter than seven years in both populations; the shortest were observed in the paternal line, except in the father-son line in PCP. The biggest difference between the two populations was in the mother-son line, 8.4 months (Table 5).
Table 5 Generation interval (years) for complete populations with register certificate (PLT) and in the selection nucleus (PCP) of Tropical Milking Criollo cattle.
| Pathway | PLT | PCP |
|---|---|---|
| Father/son | 7.2 ± 3.6 | 7.0 ± 3.4 |
| Father/daughter | 6.3 ± 3.2 | 6.6 ± 3. |
| Mother/son | 7.4 ± 4.5 | 6.7 ± 3. |
| Mother/daughter | 7.0 ± 3.8 | 7.3 ± 3. |
| Parent/offspring | 6.8 ± 3.6 | 6.9 ± 3. |
Discussion
The difference of f and fe indicates the loss of genetic variability in the original PLT population, as a result of the unequal contributions of the founders by preferential use of some parents (Boichard et al., 1997); this decrease also occurred in the PCP to a lesser extent. The fe (38%) decrease in the PCP is due to a limited number of individuals coming from the PLT (Cervantes et al., 2009). In an ideal population f equals fe and this in turn fa (Kliman et al., 2008). The genetic variability could be reduced by the effect of the genetic drift by its small size (Lacy, 1989). The fe/f ratio estimated in the PLT was 0.13, which indicates the effects of selection. In the PCP the ratio was 0.38, higher than in Colombian creole (criollo) breeds such as Costeño Con Cuernos (CCC, 0.23), Sanmartinero (SM, 0.21), and Romosinuano (Romo, 0.12), but lower than in Blanco Orejinegro (BON, 0.50) (Martínez et al., 2008). The larger estimate in the PCP nucleus is related to the selection program for dams of bulls within each of the six families. The relatively high number of ancestors regarding founders in both populations indicates a shallow genealogy and a short founder - last generation period. The fa is useful for detecting recent significant changes in the breeding improvement strategy (Goyache et al., 2003). The existence of bottlenecks can be assessed by the ratio fa/fe. These indexes were greater than 0.6 in both populations and indicate that these bottlenecks have not been narrow (Boichard et al., 1997). Similar estimates were obtained in the CCC (0.66), BON (0.65), and Limonero (0.60) breeds, and lower in SM (0.53), and Romo (0.34) (Martínez et al., 2008; Villasmil-Ontiveros et al., 2008). Close to one fa/fe values are common in populations oriented to genetic conservation (Korrida et al., 2013).
A Ne of at least 50 animals per generation leads to an F increase of 1% per generation. The PLT Ne= 68.1 would not be endangered (FAO, 1998, 2013; Falconer and MacKay, 1996) nor the PCP Ne= 64.6. The Ne of the PCP was higher than in other populations with per se genetic conservation programs; such as bull fight cattle populations (55.5 and 44.5), Reyna breed (≤46) and Moroccan Houbara Bustard (>40) (Corrales et al., 2010; Domínguez et al., 2010; Korrida et al., 2013). Some small-size dairy populations have shown that genetic improvement may involve the loss of genetic variability (Boichard et al., 1996); Ne is particularly sensitive to unequal number of females and males in the population (Kliman et al., 2008). As the genealogy integrity influences F and GCI, only genealogies with PEC index >0.8 are reliable for estimating these parameters (MacCluer et al., 1983; Corrales et al., 2010). The inbreeding estimated was 3.48% in the PCP, with 8.5% animals with F higher than 6.25%. As for the genealogical information kept within the generations, the PCP was slightly lower than the values of CG= 2.1, MG= 8.2, and EG= 4.5 estimated for double-purpose Slovenian spotted cattle (Hazuchová et al., 2012a); the highest MG= 6.9 estimated in Colombian criollos was similar to the one estimated in the PCP, although the maximum EG= 4.8 in Romo was higher; Romo had the largest population size and genealogical depth, in relation to other Colombian breeds (GE interval 3.1-3.8) (Martínez et al., 2008).
The estimated interval of AR in Spanish native breeds was 0.1-1.7% (Gutiérrez et al., 2003). The AR is useful for predicting inbreeding in the long run (Gutiérrez et al., 2010). The GCI for the PCP population was 3.73%, similar to 4.18% estimated in the dual-purpose Slovak bull population (Hazuchová et al., 2012b). However, animals with high F are not those with the greatest GCI; this index assumes that the genetic goal is to preserve and retain the full allele range that the initial population has (Gutiérrez et al., 2010). Through time, the matings made it possible for an individual to have information from more than one of the founders so the GCI increased (Figure 1), as has happened with the population of Slovak bulls (Hazuchová et al., 2012b). This difference indicates that in the PCP selection nucleus, rotational mating has decreased the tendency of F, and the selection of animals with better milk yields has decreased GCI; a different situation in the PLT, where the family rotation mating has been blurred. The generation intervals (GI) for the PLT were similar to those of Reyna (6.9) and SM (6.8), although higher than other Colombian criollos, BON (4.5), CCC (5.4) and Romo (5.7) (Corrales et al., 2010; Martínez et al., 2008). The GI through the mother-son line was the longest in the PLT; in the PCP selection nucleus, genetic improvement is carried out through the dams of bulls having the highest genetic values for milk production (Rosendo-Ponce and Becerril- Pérez, 2015). The GIs through the paternal line were shorter than those through the maternal line, which agrees with previous studies (Corrales et al., 2010, Martínez et al., 2008). The father-daughter line was the shortest; the cattlemen retain all females born in their farms. To shorten the GI by the paternal and maternal lines, sires must be replaced every two- three years and reproductive life of heifers starts at a younger age. The overall GI in the PLT was 6.8±3.9 years higher than in Brangus, Santa Gertrudis and Saler (Delgadillo, 2014), but similar to ring fight cattle (Domínguez et al., 2010). The differences found in the GI can be attributed to variations in age at puberty and first parturition; as well as to the tropical fluctuating environmental conditions in years. The mating of bulls and females that had better genetic evaluation for milk production was favored. The loss of genetic variability happens by generation; thus, as GI shortens, the annual genetic variation loss can be accelerated.
Apparently, the Tropical Milking population (PLT) is not currently endangered. Its effective population size allows for continued genetic improvement through selection for milk production, avoiding matings between related individuals. Mating of sires and females with better genetic evaluation for milk production in the selection nucleus (PCP) has led to differences between the estimates of the effective founder number and the effective ancestor number. Trends toward increasing the inbreeding coefficient and the genetic conservation index were found; from year 2000 they decreased in the PCP. Genealogy integrity in the PLT could influence the obtained estimates, reflecting higher inbreeding levels and reduced effective population size.

