











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Recommendations on the minimum total dietary neutral detergent fiber (NDF) and the minimum level of forage NDF (fNDF) for cows are 250 g/kg of total NDF, and 190 g/kg of fNDF, respectively. These values have been defined through a compilation of research studies evaluating the performance of dairy cows under specific feeding conditions (NRC, 2001).
While there is a comprehensive understanding of the role that dietary fiber plays as a nutrient and its function during digestion in cattle, the role of fiber digestion in goats has not received similar attention (Lu et al., 2008). Goats differ from cattle in their feeding behavior, the level of fiber they ingest and their dietary selections (Reid et al., 1990). Based on voluntary dry matter intake (DMI), it is clear that the demand for energy and the physical capacity of the digestive tract direct the level of intake for a particular animal. Therefore, information obtained from other ruminant species cannot be simply extrapolated to goats.
Among the various lines of research related to ruminant nutrition, the study of the relationship between NDF and NFC is of great importance. Suitable NDF levels in a ruminant’s diet during production are usually adjusted in order to maintain normal function of the rumen, maintain microbial protein synthesis (Xu et al., 2014), and optimize its intake and digestibility (Oliveira et al., 2011). Similarly, the NFC content has been considered in ruminant ration formulations as a nutritional criterion that is adjusted to obtain the appropriate fiber fraction in the feed (NRC, 2001). However, it is not known which of these components (fNDF or NFC) most affects the rumen microorganisms of goats.
For goats, there are no recommendations on the maximum and minimum values of fiber and NFC in the diet to maximize intake, animal performance and/or maintainance of normal conditions in the rumen. Thus, the evaluation of diets with different fNDF and NFC ratios under in vitro fermentation conditions can be used to determine the minimum and maximum quantities of fNDF and NFC that should be included in goat feed, since information on this topic is scarce.
Therefore, this study aimed to evaluate the effects of different forage neutral detergent fiber (fNDF) and non-fibrous carbohydrates (NFC) ratios on ruminal fermentation products and in vitro neutral detergent fiber (NDF) digestibility in goats.
Material and methods
Ethical considerations
The experimental procedures were approved by the Committee of Ethics on Animals at the Federal University of Paraiba (approval number: 2105/2013).
Location
In vitro experiments were conducted in the municipality of Areia, Paraiba (Brazil), located at an average altitude of 623 m, with a 6° 57’ 46” S latitude and 35° 41’ 31”W longitude.
Animals
Two ruminally fistulated goats, fed a total mixed ration (TMR) consisting of 60:40 forage to concentrate ratio on a DM basis, were used as rumen liquid donors. The animals were fed corn silage and a concentrate composed of ground corn, soybean meal and a mineral supplement.
Rumen fluid was collected 2 h after the morning feeding, filtered through four layers of cheesecloth, packed in a thermal bottle and immediately taken to the laboratory. The rumen fluid was transferred to 500 mL Erlenmeyer flasks, which were gassed for 20 s with anaerobic-grade CO2. Then, they were sealed with plastic wrap and placed at rest for 30 min in an environmental chamber SP LABOR (SP-500 model, Presidente Prudente, SP, Brasil) at 39ºC. After formation of liquid interfaces, the intermediate liquid was withdrawn in order to obtain an inoculum containing the most active microbial population (Oliveira et al., 2012).
Experimental design
The experiment consisted of a completely randomized design in a 3 × 5 factorial scheme with three levels of fNDF (100, 190, and 290 g/kg) and five NFC levels (350, 400, 450, 500, and 550 g/kg), with three replicates each. The experimental diets were formulated using Tifton-85 hay, ground corn, soybean meal, wheat bran, urea and starch (Table 1).
Table 1 Chemical composition of ingredients used to formulate the experimental diets.

DM = dry matter; g/kg on a fresh matter basis; 1NDF = Neutral Detergent Fiber; NDFap = Neutral Detergent Fiber corrected for ash and protein; NFC = Neutral Fiber Carbohydrate. 2Data from Brazilian Tables of Food Composition for Cattle (Valadares Filho et al., 2015).
Different combinations of fNDF and NFC levels were achieved through the use of experimental diets formulated to be isonitrogenous according to (NRC, 2007) (Tables 2 and 3).
Samples of Tifton-85 hay, ground corn, soybean meal and wheat bran used in the formulation of experimental diets were dried in a forced-air oven at 55°C for 72 h, ground in a Wiley mill (Wiley mill, Arthur H. Thomas, PA, USA) with a 1-mm screen, and stored in plastic containers for further laboratory analyses. Dry matter (DM) was determined with method 934.01. Organic matter (OM) was determined by the ash method (method 924.05). Crude protein (CP) was calculated by determining the total nitrogen content using the micro-Kjeldhal technique (method 920.87), and using a fixed conversion factor (6.25). Ether extract (EE) was determined gravimetrically after extraction with petroleum ether in a Soxhlet apparatus (method 920.85), and acid detergent fiber (ADF) (method 973.18) as described by the AOAC (1990). Neutral detergent fiber (NDF) was determined as described by Mertens (2002), and sulfuric acid lignin as described by Robertson and Van Soest (1981). The NDF content was corrected for ash and protein, and performed according to Licitra et al. (1996) and Mertens (2002).
Table 2 Proportion of ingredients used to formulate the experimental diets (g/kg DM).
fNDF = forage neutral detergent fiber; NFC = non-fibrous carbohydrates.
The NFC were calculated according to Detmann and Valadares Filho (2010): NFC (g/kg) = 1000 − [(CP − CPU + U) + NDF + EE + ash], where: CPU is the CP content from urea (%), and U is the urea content (%). Aliquots of the experimental diets (350 mg of DM) were packed in 50 mL glass bottles. McDougall’s buffer (McDougall, 1948) was prepared in Erlenmeyer flasks, and the pH was adjusted to 6.8 by saturating it with CO2 gas. To each glass bottle, 28 mL of McDougall’s buffer and 7 mL of strained ruminal fluid were added. Bottles containing all experimental ingredients were immediately gassed for 10 s with anaerobic-grade CO2 and then sealed. The final composition of the treatments was 100 mg of forage (DM) in a 10 mL final solution, and 1 mL of ruminal fluid in 4 mL of buffer solution (Tilley and Terry, 1963). Bottles were incubated at 39°C for 48 h. The incubation was conducted according to the partial methodology described by Tilley and Terry (1963). The incubation times were 0, 3, 6, 12, 24 and 48 h. The incubation process was repeated three times, generating three evaluations per incubation time for each treatment. Once the fermentations were complete, 1.0 mL aliquots were collected from each bottle using a syringe with a 0.80 x 25 mm needle in order to maintain anaerobic conditions within the tubes (Lana et al., 2000). The pH was measured using a digital pH meter (TECNOPON, Piracicaba, SP, BRA). Aliquots were acidified with meta-phosporic acid (one part acid to one part sample) and centrifuged (Hettich Mikro 200 Model, Tuttlingen, BW, GER) at 5200 × g for 10 min. The supernatant was frozen (−20°C) and subsequently analyzed for volatile fatty acids (VFA) by high performance liquid chromatography (HPLC) according to Siegfried et al. (1984). The HPLC (SPD-10 AVP, Shimadzu Corporation, Osaka, OS, Japan) apparatus was equipped with a refractive index detector and used an Aminex HPX-87H column (BIO-RAD, Hercules, CA, USA) with a mobile phase containing 5 mM H2SO4 and a flow rate of 0.6 mL/min at 50°C. The VFAs were measured at 0 and 48 h. Molar proportions of VFA were calculated by dividing the concentration of individual acids by the total concentration of VFA. Ammonia was determined by the colorimetric method of Chaney and Marbach (1962). The resulting pellets were washed with a 0.9% (w/v) NaCl solution, centrifuged at 5200 × g for 10 min, resuspended in a 0.9% (w/v) NaCl solution and frozen at −20°C. Microbial protein was estimated using the method of Bradford (1976). At the end of the incubation (48 h), the residue obtained from each bottle was vacuum filtered in a gooch crucible (gross porosity) and conditioned in bottles (120 mL) with the addition of 50 mL of a neutral detergent solution (Mertens, 2002). After sealing, the polyethylene bottles were autoclaved at 105 °C for 1 h in order to extract all of the soluble components (neutral detergent micro-extraction method (Pell and Schofield, 1993). After this procedure, the gooch crucibles were again submited to vacuum filtration followed by sequential washing with hot distilled water and acetone. The NDF residue obtained was dried at 105°C for 16 h. The NDF digestibility was calculated using the following equation: NDF digestibility = (amount of NDF present in extracted sample bottles) - (amount of NDF in sample after washing)/(amount of NDF present in extracted sample bottles) × 100.
Statistical analysis
Data were analyzed using the GLM procedure of SAS software, version 9.4 (SAS Institute, Inc, Cary, NC, USA) (2012). Regression models from different fNDF levels were submitted to a model identity test. The choice of regression equation was based on the determination coefficient and the significance of regression coefficients using the t test, where α = 0.05.
Results
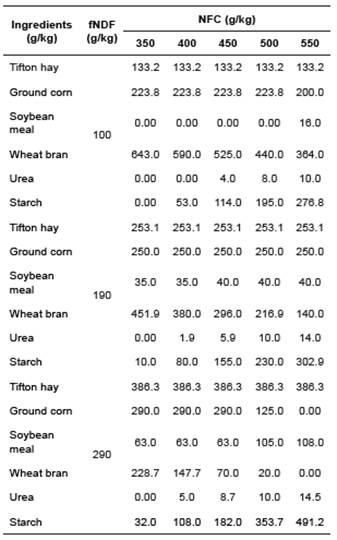
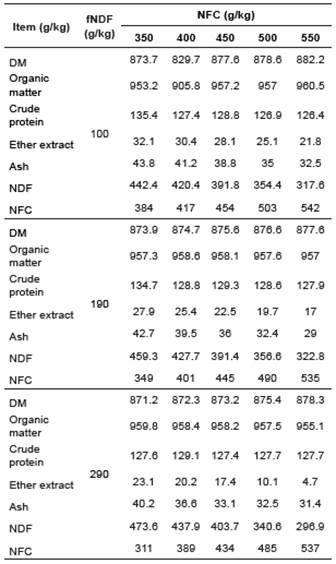
The ammonia concentration varied throughout the fermentation process as a function of NFC levels in a similar pattern for the three fNDF levels studied, all reaching maximum concentrations after 48 h (Figures 1a, 2a, and 3a). Microbial protein concentration had a decrease after 24 h of incubation (Figure 1b, 2b and 3b), except in the diet containing 29% fNDF with 45 and 55% NFC.
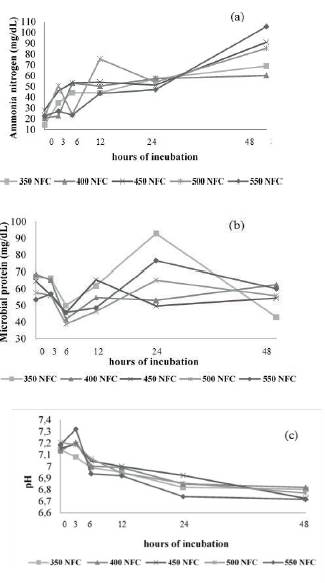
Figure 1 Effect of NFC levels on ammonia (a) and microbial protein (b), and changes in acidity (c) throughout the fermentation time by ruminal microorganisms in diets containing 100 g/kg fNDF.
Figure 2 Effect of NFC levels on ammonia (a) and microbial protein (b) concentration, and changes in acidity (c) throughout the fermentation time by ruminal microorganisms in diets containing 190 g/kg fNDF.
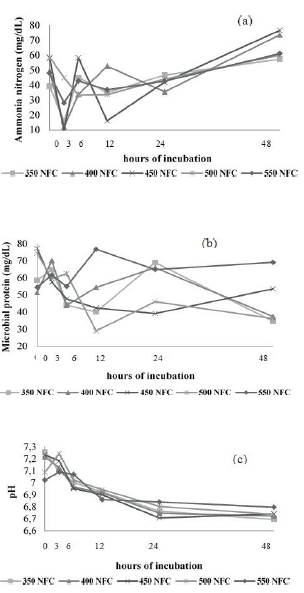
Figure 3 Effect of NFC levels on ammonia (a) and microbial protein (b) concentration, and changes in acidity (c) throughout the fermentation time by ruminal microorganisms in diets containing 290 g/kg fNDF.W
Microbial protein concentration after 48 h of fermentation had a linear crescent effect (p<0.05) on all fNDF levels as a function of increased NFC levels, except for diets containing 190 g/kg fNDF (Table 4). When the model identity test was carried out, the linear model of NFC levels for the fNDF levels of 100 and 290 g/kg were similar (p>0.05).
Table 4 Effect of forage neutral detergent fiber (fNDF) and non-fibrous carbohydrates (NFC) levels on microbial protein (MP), ammonia concentration and pH during in vitro fermentation.
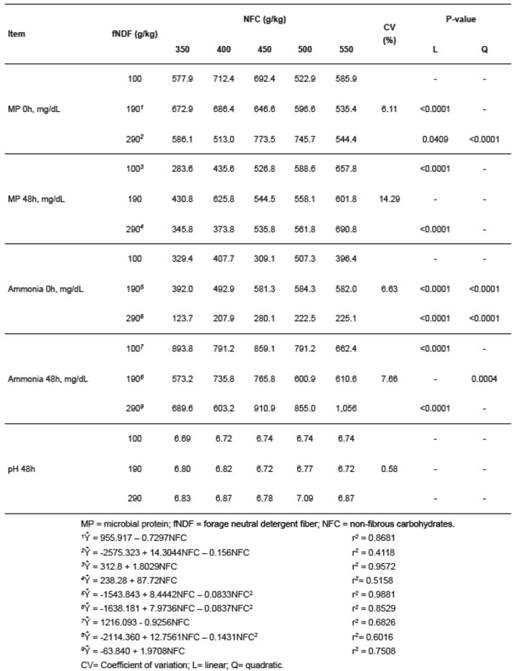
Increasing NFC levels in media containing 100 and 290 g/kg fNDF had a linear decrease and increase (p<0.05) in ammonia concentration until 48 h of fermentation, respectively (Table 4). The experimental diet containing 190 g/kg fNDF resulted in a quadratic increase (p<0.05) in ammonia concentration as a function of NFC levels. When the model identity test was carried out, the linear models chosen among fNDF levels differed according to the NFC levels (p<0.05). Thus, NFC levels act differently among the fNDF levels in ammonia production. With the exception of media containing 100 g/kg fNDF, the increase in NFC levels promoted an increase in the ammonia concentration (Table 4).
Different combinations of fNDF and NFC levels had no effect (p>0.05) on the pH (Table 4).
Even with significant increases in total VFA production at 48 h of fermentation (Table 5), the amount of acid was not enough to decrease the pH.
Table 5 Effect of forage neutral detergent fiber (fNDF) and non-fibrous carbohydrates (NFC) levels on volatile fatty acids (VFA) concentration and individual molar proportion during in vitro fermentation.
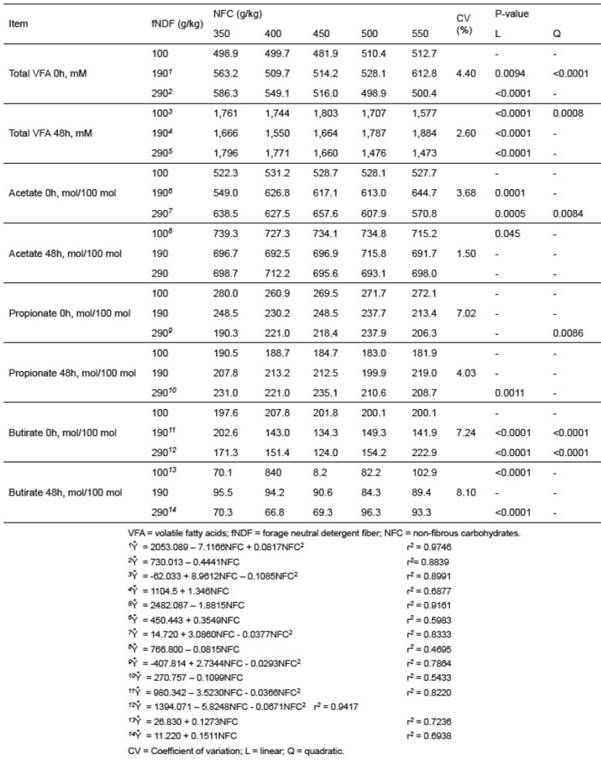
Total VFA and individual molar ratios were different (p<0.05) among fNDF and NFC levels (Table 5). Total VFA concentrations at 48 h were higher than at 0 h for all combinations of fNDF and NFC. The diet containing 190 g/kg fNDF quadratically increased (p<0.05) total VFA with the addition of NFC; however, a linear increase (p<0.05) was observed in diets containing 290 g/kg of fNDF. When the model identity test was carried out, the models of fNDF as function of NFC levels were different (p<0.05). Thus, the change in NFC levels acted differently among the fNDF levels for total VFA production.
In general, significant changes were not observed between the individual molar ratios of acetate, propionate, and butyrate with the addition of NFC at each fNDF level (Table 5).
Changes in fNDF and NFC levels resulted in interaction (p<0.05) on in vitro NDF digestibility. Only diets containing 290 g/kg fNDF had a quadractic relationship (p<0.05) to increasing NFC levels and in vitro NDF digestibility with maximum values estimated at 659.6 g/kg for 373.2 g/kg NFC (Table 6).
Table 6 Effect of forage neutral detergent fiber (fNDF) and non-fibrous carbohydrates (NFC) levels on in vitro NDF digestibility.
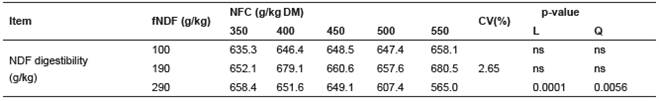
fNDF = forage neutral detergent fiber; NFC = non-fibrous carbohydrates. Ŷ = 212.67 + 23.763CNF - 0.315CNF2r2 = 0.9845 CV = Coefficient of variation; L = linear; Q = quadratic.
Discussion
As in vitro fermentation experiments take place in closed environments, and no ammonia is absorbed through the rumen walls as it occurs in in vivo experiments, the ammonia concentration was directly related to microbial growth since it is used as a nitrogen source for microbial protein synthesis (Russell, 2002; Seo et al., 2010; Seo et al., 2013) (Figures 1a, 2a and 3a).
Thus, we observed that the microbial standard in goats varied more with changes in NFC concentration than changes in fNDF concentration (Figures 1b, 2b and 3b). This difference in the rumen environment of goats must be associated with their feeding behavior as they are concentrate pickers and much of their diet comes from less coarse foods. Thus, it is possible that changes in the fNDF concentration less significantly modify the ruminal environment relative to variations in NFC concentration. Similar behavior observed in the three fNDF levels studied reinforces this hypothesis.
Thus, NFC levels affected microbial protein production similarly, regardless of the amount of fNDF contained in the media (Table 4). Thus, we concluded that microbial protein concentration in goats is influenced more by NFC levels than by fNDF levels. Providing a greater amount of rapidly fermentable carbohydrates in the form of NFC created an environment favorable for bacterial growth and increased the amount of energy available to the rumen microoganisms for multiplication (Russell et al., 1992; Costa et al., 2008; Ma et al., 2014; Ma et al., 2015). Coupled with an increase in energy levels for microbial protein synthesis, ruminal ammonia levels greater than 5 mg/dL should be an adequate nitrogen source for rumen microrganisms (Satter and Styler, 1974).
The increase in the ammonia concentration, promoted by the increase in NFC levels, may be occur due to the increased amounts of urea added to the experimental diets, as ammonia is produced both by deamination of amino acids and hydrolysis of nonprotein nitrogenous compounds (Valadares Filho and Pina, 2011) (Table 4).
The increase in total VFA was not enough to decrease the pH, probably due to the strong buffering capacity of McDougall’s solution (Table 5). Several in vitro studies using this solution reported minor variations in the pH, regardless of the substrate used and whether its degradation was slow or rapid (Costa et al., 2008; Costa et al., 2009; Berthiaume et al., 2010; Purcell et al., 2014).
We expected an increase in the concentration of total VFA as NFC levels increased; however this was not observed for the fNDF levels 100 and 290 g/kg (Table 5). Under ideal conditions with excess fNDF, providing more rapidly fermentable carbohydrates promoted the growth of fermenting NFC microrganisms and increased VFA concentrations in the media (Vieira et al., 2008). Berthiaume et al. (2010) and Ma et al. (2015) observed similar increases in total VFA concentration when the NFC/ NDF ratio was improved. However, increased NFC concentrations in the media may not influence or even decrease the total VFA concentration due to a simultaneous increase in fNDF concentration or total NDF in the diet (Archimede, 1996; Cantalapiedra- Hijar et al., 2009).
The absence of significant changes in individual molar ratios of acetate, propionate and butirate may have occurred due to small variations on H+ concentrations, which affect pH and the metabolic pathways of some microrganisms. In this case, the metabolic fermentation pathways of microrganisms were not modified, which would have interfered with the production of acetate and propionate (Russel, 2002; Jiang et al., 2013); thus the proportions of these metabolites were similar in the different combinations of fNDF and CNF studied. Russel (1988) reported that some ruminal microrganisms are capable of producing acetate and propionate in combination with other products such as formate, ethanol and CO2, and that a change in acidity is capable of modifying the fermentation metabolic pathways of microrganisms (Aschenbach et al., 2011; Castillo-González et al., 2014). As an example, when fermentation is slow and extracellular pH is near neutral, Streptococcus bovis produces acetate, formate and ethanol, which are products of primary fermentation when growth rate is slow (Russel, 1988; Castillo-González et al., 2014). These products were produced by the enzyme pyruvate formate lyase. On the other hand, if extracellular and intracellular pH declines, as is the case with more rapidly fermentable carbohydrates available than what is needed to increase growth rate, the pyruvate produced is converted into lactate. In this specific case, the change in metabolic pathways and the change in fermentation end products happens because pyruvate lyase becomes inactive at low pH (Russel, 2002).
Studies reported changes in the proportions of the main VFA, where low and high NFC/NDF ratios are evaluated in sheep (Ma et al., 2015), and goat (Cantalapiedra-Hijar et al., 2009). In contrast, Costa et al. (2008) did not find changes in the proportions of the mains VFA. In this last study, authors noted that there was no rapid degradation of carbohydrates, which was indicated by the high pH. Thus, pH is an important variable that responds to variations of the main VFA in the rumen. As observed in this study, the absence of pH variation did not cause significant changes in VFA molar ratios, even at high NFC/NDF ratios.
Diets with high NFC/NDF ratio resulted in lower NDF digestibility, probably due to the decline in pH (Cantalapiedra-Hijar et al., 2009; White et al., 2010; Ma et al., 2015) that directly impacted how different microbial groups ferment carbohydrates (Homen Júnior et al., 2015). Increased VFA production caused a decrease in pH and inhibited growth of bacteria that ferment fibrous carbohydrates, thus compromising NDF digestibility (Van Soest, 1994; Krajcarski-Hunt et al., 2002; Sung et al., 2007; Cerrato-Sánchez et al., 2008 Russell et al., 2009). However, in the present study, the minimum pH value observed was approximately 6.70 (Figure 1c). It likely did not decrease significantly due to the buffering capacity of the media, and there was not a decrease in NDF digestibility when higher NFC concentrations were used (Table 6).
In conclusion, the relationship between fNDF and NFC concentrations affects fermentation end products and in vitro NDF digestibility. The NFC levels appear to play a more consistent role over rumen microbial ecosystem during in vitro fermentation. Changes in the microbial standard of goats during in vitro fermentation are more affected by changes in NFC concentrations than fNDF concentrations. Thus, a reduction in NFC levels will allow goat diet formulations with minimum levels of NDF.

