











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Worldwide aquaculture production exceeds 60 million tons per year and contributes with more than 40% to the production of aquatic organisms (FAO, 2016). However, diseases caused by viruses and bacteria are a primary concern for the development of this important activity (Lightner, 2011). White spot disease caused by white spot syndrome virus (WSSV) is a devastating disease for cultured penaeid shrimp (Joseph et al., 2015) and cannot be treated with antibiotics as it happens with diseases caused by bacteria (Gatlin, 2002).
Immunostimulants are an alternative to antibiotics to prevent diseases in cultured animals (Cheng et al., 2014) since they increase animal resistance (Song and Hsieh, 1994). Regarding crustaceans, the immunostimulatory effect of molecules (sodium alginate, glucans, lipopolysaccharides, nucleotides, peptidoglycans, and chitosan) and of whole cells (yeasts and bacteria) have been studied (Luna González et al., 2013a; Ringø et al., 2012; Sajeevan et al., 2009a, b). Regarding yeast, Candida parapsilosis possesses immunostimulatory activity against WSSV and has supported high post-challenge WSSV survival (Wilsi et al., 2015).
Crustaceans, such as Litopenaeus vannamei, have two lines of defense. The first line is a physical barrier consisting of the cuticle, which prevents the entry of pathogens in the body. The second line is based on cellular and humoral effectors, which combine to combat pathogens, heal wounds and respond to biotic and abiotic stressors (Lemaitre and Hoffman, 2007). The cellular effectors in shrimp are hemocytes (granular, semigranular, and hyaline), which play a central role in phagocytosis, melanization, encapsulation, nodulation, cytotoxicity, apoptosis, interfering RNA, autophagy, interferon-type system, and coagulation of the hemolymph (Goncalves et al., 2014; Sun et al., 2010). The humoral effectors are the second line of defense and include agglutinins, profenoloxidase system, coagulation proteins, hydrolytic enzymes, antimicrobial peptides, nitric oxide, respiratory burst, and opsonization (Morales and Cuéllar et al., 2014; Cerenius and Söderhäll, 2004; Destoumieux et al., 2000; Jiravanichpaisal et al., 2006). The expression of immune-related genes in shrimp provides important information about the activation of the immune system and its modulation (Wang et al., 2007).
The aim of this work was to determine the effect of microbial immunostimulants on WSSV infection percentage and on the expression of immune-related genes in white shrimp (Litopenaeus vannamei), cultured in the laboratory and challenged with WSSV.
Materials and methods
Ethical considerations
This study complies with the Mexican Official Standard NOM-062-ZOO-1999, technical specifications for the production, care and use of laboratory animals.
Animals and acclimatization
Shrimp were obtained from Cuate Machado farm (Guasave, Sinaloa, Mexico) and transported to the CIIDIR-IPN facilities in a plastic tank (250 L) with sea water and constant aeration. Animals were maintained in an outside culture system in 120 L plastic tanks with 80 L filtered (20 μm) sea water (30 ‰) and continuous aeration. Shrimp were fed with commercial feed (Purina®, Ciudad de Mexico, Mexico, 35% protein) twice daily at 09:00 and 16:00 h.
Preparation of viral inoculum and tissue paste
A viral inoculum (stored at -70 °C) with high viral load (single PCR) was used to infect shrimp to obtain fresh muscle and gills with WSSV. Muscle and gill tissues of moribund shrimp were analyzed by polymerase chain reaction (PCR) to confirm the presence of the virus. Half of the tissues obtained were stored at -70 °C in Falcon tubes and the other half were placed in cold Eppendorf tubes to be macerated with a plastic pistil. After maceration, 1 g of tissue was placed in a 20 mL Falcon tube with 10 mL of cold saline solution (2%). Individual samples were centrifuged at 17,000 g, for 10 min, at 4 °C. The supernatants were filtered with syringe filters (0.45 and 0.25 μm), placed in cold Eppendorf tubes (1.5 mL) and stored at -70 °C. The presence of WSSV in the inoculum was verified with simple (>1000 copies of viral DNA, Lo et al., 1996a, b) and nested (10-50 copies of viral DNA, Lo et al., 1996a, b) PCR.
Microbial immunostimulants (MI)
Lactic acid bacteria (Pediococcus parvulus Lta2) and yeast (Candida parapsilosis Lt6) were isolated and characterized according to Apún Molina et al. (2009) and Luna González et al. (2013b), and tested in L. vannamei challenged with Vibrio sinaloensis according to Flores Miranda et al. (2011).
Preparation of the experimental diet with powdered immunostimulants
Microorganisms were grown and washed as described in Apún Molina et al. (2009). Cells were centrifuged at 5,000 g for 10 min and dried in an oven (Felisa, Zapopan, Jalisco, Mexico) at 74 °C for 4 h. The dried cell pellet was ground in a mortar and then included in the commercial feed pellet (35% protein) at 8.5 mg/kg feed (50% P. parvulus and 50% C. parapsilosis). The amount of microbial immunostimulants (MI) was based on the work by Flores Miranda et al. (2011).
Experimental design
The bioassay was conducted for 33 d with shrimp weighing 9.9 ± 3.1 g. Ten shrimp were placed per tank. Treatments in triplicate were as follows: I) Control group, shrimp fed commercial feed + WSSV; II) shrimp fed commercial feed + MI (8.5 mg/kg feed) daily + WSSV; III) shrimp fed commercial feed + MI (8.5 mg/kg feed) every 2 d + WSSV; IV) shrimp fed commercial feed + MI (8.5 mg/kg feed) every 3 d + WSSV. Shrimp were fed feed plus MI, except for the days (9 and 19) when animals were fed 250 mg per tank of shrimp paste (muscle and gills) with a high WSSV load (single PCR, > 1000 viral particles). Healthy shrimp were fed on dead shrimp with WSSV to simulate a natural infection. Initially, animals were fed 5% of the mean body weight and adjusted thereafter according to the feeding response in each tank. Uneaten food and waste material were removed every 3 d before feeding, and 50% of the water was exchanged every 5 d.
Physicochemical parameters like pH (HI 98127 pH, Hanna Instruments, Woonsocket, RI, USA), salinity (refractometer W/ATC 300011, Sper Scientific, Scottsdale,AZ, USA), dissolved oxygen, and temperature (YSI model 55 oxygen meter, Yellow Spring Instruments, Yellow Springs, OH, USA) were monitored every 2 d. At the beginning and the end of the bioassay, ammonium, nitrites, and nitrates were determined by the method of Strickland and Parsons (1972).
At the end of the bioassay, weigth and survival of shrimp were determined. Shrimp were analyzed separately for WSSV by single or nested PCR. In addition, hemolymph was extracted for the immune system analysis. During the bioassay, the physicochemical parameters (temperature, salinity, pH, dissolved oxygen, ammonium, nitrites, and nitrates) were kept within the optimal intervals for shrimp culture according to Brock and Main (1994) and Boyd and Tuker (1988). Temperature ranged from 29.0 ± 0.2 to 29.5 ± 0.3 °C; salinity was between 31.5 ± 0.6 and 32.2 ± 0.6‰; disolved oxygen ranged from 5.6 ± 0.06 to 5.8 ± 0.05 mg/L; pH was between 8.4 ± 0.02 and 8.5 ± 0.02; ammonium ranged from 0.2 ± 0.2 to 0.6 ± 0.20 mg/L; nitrite was between 0.012 ± 0.001 and 0.014 ± 0.002 mg/L; and nitrate ranged from 0.05 ± 0.05 to 0.7 ± 0.01 mg/L.
DNA extraction and WSSV infection percentage
Genomic DNA was extracted from abdominal muscle and gill lamellas (1:1 w/w) (100 mg) with DNAzol reagent (MRC®, Cincinnati, OH, USA) following manufacturer’s instructions. Quantification and quality assessment of DNA were performed in a Pearl nanophotometer (Implen, Inc. Westlake Village, CA, USA). DNA was stored at -20 °C, until use. Twelve shrimp per treatment (four per tank) were sampled for WSSV infection percentage by single and nested PCR, using the primers WSSV out-1/WSSV out-2 (5’-ATC ATG GCT GCT TCA CAG AC-3’ [forward]/5’-GGC TGG AGA GGA CAA GAC AT-3’ [reverse]) and WSSV in-1/WSSV in-2 (5’-TCT TCA TCA GAT GCT ACT GC-3’ [forward]/5’-TAA CGC TAT CCA GTA TCA CG-3’ [reverse]) (Kimura et al., 1996), which amplified genome fragments of 982 and 570 bp, respectively. Negative samples were analyzed by one-step PCR with an internal control that amplified a 298 bp segment of shrimp glyceraldehyde- 3-phosphate dehydrogenase (GAPDH) DNA, using the primers GAPDH298 and GAPDH298 (5’-TCA CCG TCT TCA ACG AGA TG-3’ [forward]/5’-ACC CTC CAG CAT CTC GAA CT-3’ [reverse]) (Tang and Lightner, 2001).
Hemolymph extraction
Two-hundred microliters of hemolymph were sampled from 3 intermolt shrimp per tank, 9 per treatment, from the pleopod base of the first abdominal segment with a sterile 1 mL siringe (27 G × 13 mm needle). Before hemolymph extraction, the siringe was loaded with 400 μL of a precooled (4°C) solution (SIC-EDTA, Na2) (450 mM NaCl, 10 mM KCl, 10 mM Hepes, and 10 mM EDTA Na2, pH 7.3) used as an anticoagulant (Vargas Albores et al., 1993). Hemolymph was placed in cold Eppendorf tubes (1.5 mL) and then centrifuged at 800 g for 6 min at 4 °C. Plasma was discharged and the celular pellet was rinsed with 250 µL of cold anticoagulant solution and resuspended in 300 µL of Trizol reagent (Invitrogen, Carlsbad, CA, USA). Samples were stored at a -70 °C until used.
Total RNA isolation and cDNA synthesis
Total RNA of hemocytes was extracted with Trizol reagent according to manufacturer’s protocol. The RNA concentration and purity was analyzed in a Pearl nanophotometer. The RNA was treated with DNAse I (1 U/μL) (Sigma-Aldrich, St. Louis, MO, USA).
Reverse transcription was used to synthesize the first strand of cDNA using reverse transcriptase (Improm II, Promega, Madison, WI, USA) with the oligo dT20, 500 ng of total RNA and an incubation temperature of 42 ºC for 60 min. The cDNA was diluted with 80 μL of ultrapure water and stored at -70 °C until analysis. Five microliters of this cDNA dilution was used as template in each qPCR reaction.
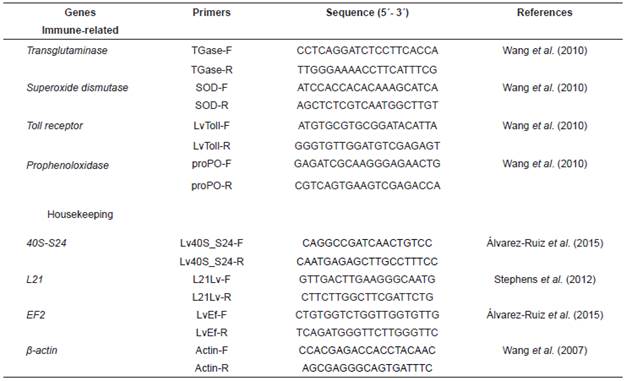
Expression analysis of immune-related genes by qPCR
The expression of four immune-related genes (toll receptor [LvToll], superoxide dismutase [SOD], prophenoloxidase [proPO], and transglutaminase [TGase]) in hemocytes (Table 1) of experimental shrimp was measured by quantitative real-time PCR using a CFX96 system and the CFX Manager version 3.0 (Bio-Rad Laboratories, Hercules, CA, USA). To find the best reference genes to normalize the expression of target genes, a stability analysis of ribosomal protein (40S-S24), actin (β-actin), elongation factor 2 (EF2), and ribosomal protein (L21) genes (Table 1) was done with the algorithms GeNorm (Vandesompele et al., 2002) and NormFinder (Andersen et al., 2004), using the RefFinder web application (http://150.216.56.64/referencegene.php). The expression of target genes was normalized to the geometric mean of the most stable reference genes (EF1α, β-actin, and L21).
The expression of target and reference genes was determined using the primers found in Table 1. Amplifications were performed in triplicate in a 96-well plate with a final volume of 15 μL, containing 7.5 μL of PCR Master Mix 2x (1.5 μL of 10x reaction buffer, 0.75 μL of 50 mM MgCl2, 0.3 μL of 10 mM dNTPs, 0.75 μL of EvaGreen 20x [Biotium, Hayward, CA, USA], 0.1 μL of 5 U/µL Biolase DNA polymerase [BiolineTM, Tauton, MA, USA], and 4.1 µL ultrapure water), 0.5 μL of each primer (10 μM, Sigma-Aldrich, St. Louis, MO, USA), 3.5 μL of ultrapure water, and 3 μL of cDNA. The thermocycler conditions were as follows: 95 °C for 3 min followed by 40 cycles at 95 °C for 10 s, 60 °C for 15 s, 72 °C for 30 s, and 79 °C for 5 s. After each reaction, a dissociation curve from 65 to 90 °C was recorded at increments of 0.5 °C, and examined for unique and specific products.
For each gene, efficiency (E) of the PCR reaction was determined by calculating a slope with five serial dilutions (dilution factor of 5 or 10) of a representative pool of cDNA [E = 10(− 1/slope) − 1]. The expression of target genes was calculated by transforming the Cq values to relative quantities (RQ), using the equation RQij = E[(Cq mean) − Cq(ij)], where E is the gene-specific efficiency, and [(Cq mean) - Cq(ij)] is the absolute difference for each Cq sample against the mean Cq in the dataset for each gene. Relative expression of each gene was calculated with the equation RQtarget/Geometric mean of RQreference genes (Vandesompele et al., 2002).
Specific growth rate
At the end of the bioassays, the specific growth rate (SGR) was determined using the following formula:
SGR (%/d) = 100 (Ln Wt - Ln W0)/t
Where t is the culture period in days, W 0 is the natural logarithm of the weight of the shrimp at the beginning of the bioassay, and ln Wt is the natural logarithm of the weight of the shrimp at day t (W 0 and Wt are in grams).
Statistical analysis
One-way variance analysis (ANOVA) using the F test was performed among treatments to examine the differences in growth performance and gene expression. Where significant ANOVA differences were found, a Tukey’s HSD test was used to determine the nature of these differences at (p<0.05) (Zar, 1996).
Results
Survival and growth
Survival in the bioassay was 100% in all treatments (Table 2). Initial weight ranged from 0.72 ± 0.01 to 0.75 ± 0.02 g. Final weight was between 4.04 ± 0.22 and 4.14 ± 0.18 g. Regarding SGR, values ranged between 5.82 ± 0.1 and 5.34 ± 0.33%/d. No significant differences among treatments were found in growth (Table 2).
Table 2 Effect of MI on the survival, growth, and WSSV infection percentage in L. vannamei cultured in the laboratory.
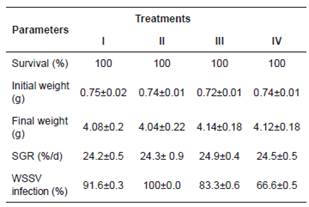
Treatments: I) Control group, shrimp fed commercial feed + WSSV; II) shrimp fed commercial feed + MI (8.5 mg/kg feed) daily + WSSV; III) shrimp fed commercial feed + MI (mg/kg feed) every 2 d + WSSV; IV) shrimp fed commercial feed + MI (mg/kg feed) every 3 d + WSSV. Data are mean±SE.
WSSV infection percentage
Nested PCR analysis showed that 58.3% of the organisms from the stock were infected with WSSV (control without WSSV added). At the end of the bioassay, the WSSV infection percentage increased 8.4% in treatment II, where shrimp were fed daily with MI, as compared with control group. However, in treatments III and IV, where shrimp were fed every two and three days with the MI, WSSV infection percentage decreased by 8.3 and 25%, respectively, as compared with control group (Table 2).
Immune-related gene expression
Figure 1 summarizes the results of gene expression in white shrimp fed MI. The additive modulated the expression of the LvToll, TGase, and proPO genes. In TGase and proPO, significant differences (p<0.05) were found among treatments I, II, and III, with higher gene expression in treatment II (shrimp fed daily with MI). In LvToll, significant differences (p<0.05) were found in treatment II as compared with treatments I, III, and IV. The mRNA expression of SOD gene remains unchanged (p>0.05).
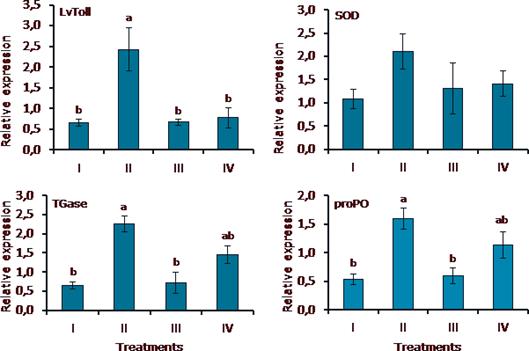
Figure 1 Immune-related gene expression in L. vannemei fed MI and challenged with WSSV. Treatments: I) Control group, shrimp fed commercial feed + WSSV; II) shrimp fed commercial feed + MI (mg/kg feed) daily + WSSV; III) shrimp fed commercial feed + MI (mg/kg feed) every 2 d + WSSV; IV) shrimp fed commercial feed + MI (mg/kg feed) every 3 d + WSSV. Relative expression of each gene was calculated with the equation RQtarget/Geometric mean of RQreference genes. Data are mean±SE. Different letters indicate significant differences.
Discussion
The negative impact of white spot disease has been found in shrimp cultured worldwide. Because of the above, substances that strengthen the immune system of L. vannamei are now being investigated and to make it more resistant to diseases caused by pathogenic microorganisms; however, there is few scientific support for its function and dose/ frequency of application (Andrews et al., 2011; Sajeevan et al., 2009a). Therefore, in this work, the immunostimulating effect of a mixture of microorganisms (lactic acid bacteria and yeast) on WSSV challenged shrimp was evaluated.
The growth of organisms is influenced by environmental conditions such as temperature, pH, salinity, and food availability (Oduleye, 1982). However, in the present work, growth was not negatively affected by the additives in the feed and because physicochemical parameters and water quality were kept within the optimal intervals for shrimp cultivation (Boyd and Tucker, 1988; Brock and Main, 1994). With respect to the survival of WSSV-infected shrimp, it was 100% in all treatments; results that agree with those reported by Partida Arangure et al. (2013), who worked with WSSV- infected shrimp using a mixture of lactic acid bacteria and bacilli in feed at a concentration of 1 x 105 CFU/g feed, as well as inulin at 0.4 to 0.8 g/kg feed. These findings corroborate that culture conditions in the laboratory are more stable that those found in the commercial shrimp farms.
The analysis showed that before the bioassay, 58.3% of the stock shrimp were infected with the virus (low viral load). Therefore, the infection in experimental shrimp was reinforced with shrimp paste. In this sense, it was observed that reinfection worked since 96.1% of the shrimp from the control group were WSSV positive. WSSV infection percentage decreased 8.3% in treatment III (shrimp fed MI every 2 d) and 25% in treatment IV (shrimp fed MI every 3 d) as compared with the control group (shrimp reinfected with WSSV). On the other hand, WSSV infection percentage in treatment II (shrimp fed MI daily) was increased 8.4% as compared with control group. The decrease in the WSSV infection percentage when the MI was administered discontinuously agree with the results obtained by Itami et al. (1998) in Penaeus monodon challenged with V. penaeicida and fed peptidoglycans (0.2% of the diet) administered discontinuously. Similarly, Sajeevan et al. (2009a) reported a higher survival of Fenneropenaeus indicus challenged with WSSV orally and fed a glucan diet (0.2%) every seven days. In the same way, Flores Miranda et al. (2011) found a higher survival of L. vannamei challenged with strains of V. sinaloensis and fed every three days with the same MI of this study.
In all living organisms, cells regulate their activities by modulating the expression of their genes. Gene expression is generally proportional to the number of copies of messenger RNA (mRNA) of a given gene. This is a crucial fact when it comes to identifying the presence of specific cellular products since the mRNA is translated into the ribosomes to form proteins. Thus, it is possible to obtain data concerning the production of biological elements if the expression of the genes of a cell is known (McPherson et al., 2008).
Toll receptors are highly conserved transmembrane proteins that are found in immune cells and recognize PMAPs (pathogen-associated molecular patterns) of microorganisms (lipopolysaccharides, peptidoglycans, betaglycans, flagellins, and others) (Akira and Hemmi, 2003). Transglutaminase (TGase) is involved in the polymerization of the coagulation protein in the plasma of decapod crustaceans (Liu et al., 2007) and is released as an immediate response of hemocytes to the presence of LPS and β-1,3-glucans (Montaño Pérez et al., 1999). Prophenoloxidase can be found in hemocytes as the inactive form of phenoloxidase and plays an important rol in the inate immune response because it allows the rapid response to bacteria and fungi (Amparyup et al., 2013). On the other hand, the SOD enzyme converts the superoxide anion to hydrogen peroxide (Wang et al., 2010) and thus maintains the state of health and survival of organisms (Campa Córdova et al., 2005). In the present study, we found that the mRNA expression of LvToll, TGase, and proPO genes was significantly increased in treatment II, where the shrimp were fed daily with the MI as compared with control group and treatments III and IV, where shrimp were fed every two and three days with feed additives. However, in SOD gene expression, the trend was similar, but there were no significant differences. It is posible that the increased gene expression of treatment II was due to the peptidoglycans and β-glucans of P. parvulus Lta2 and C. parapsilosis since WSSV load in all treatments was low (10-50 viral particles). Moreover, it is known that the expression of LvToll, TGase, proPO, and SOD genes can be suppressed or remain unchanged when shrimp are infected with WSSV (Ai et al., 2008; Arts et al., 2007; Wang et al., 2010; Yeh et al., 2009). Although genes were up-regulated in treatment II, the lower WSSV infection percentage occurred in treatment IV, where shrimp were fed every 3 days with MI. It is possible that continuous administration of immunostimulants causes immune fatigue in shrimp as reported by Bai et al. (2010) in L. vannamei fed β-glucan or glycyrrhizin. The above findings point out the importance of concentration and the frequency of application of immunostimulants in shrimp culture systems for effective protection against pathogens (Babu et al., 2013; Sajeevan et al., 2009a). Further research is needed about the effect of higher MI concentrations, added to the feed, in shrimp immune response and WSSV infection percentage.
In conclusion, despite the short period of time (14 days) from the last infection and the end of the bioassay, the tested MI decreased WSSV infection percentage in L. vannamei infected with low viral load when it was applied every three days. Also, results showed that MI up-regulates LvToll, TGase, and proPO genes, when it is given daily. Further research is needed about microbial immunostimulants as prophylactic treatment in commercial shrimp farms against WSSV.

