










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkColombia Forestal
Print version ISSN 0120-0739
Colomb. for. vol.13 no.1 Bogotá June 2010
EFECTO DE LA INTERVENCIÓN ANTRÓPICA EN LA DISTRIBUCIÓN DE LAS SEMILLAS Y PLÁNTULAS DEL ROBLE (Quercus humboldtii Bonpl., FAGACEAE) EN LA CORDILLERA ORIENTAL COLOMBIANA1
Effect of human disturbance on seed and seedling distribution of the Andean Oak (Quercus humboldtii Bonpl., Fagaceae) in the Colombian Eastern Cordillera
Efeito da intervenção antrópica na distribuição das sementes e mudas de roble(Quercus humboldtii Bonpl., Fagaceae) na cordilheira oriental colombiana
Sandra Bibiana Guerrero-Rodríguez2, Erika Andrea Paz-Camacho2 & Ángela Parrado Rosselli2
1Investigación financiada por la Fundación Natura Colombia en el marco del macroproyecto “Corredor de conservación de robles, Guantivá La Rusia – Iguaque: una estrategia para la conservación y el manejo forestal en Colombia”2Grupo Uso y Conservación de la Diversidad Forestal, Proyecto Curricular de Ingeniería Forestal, Facultad del Medio Ambiente y Recursos Naturales, Universidad Distrital Francisco José de Caldas, Avenida Circunvalar - Venado de oro, Bogotá. aparrador@udistrital.edu.co Autor para correspondencia.
Recepción: Agosto 5 de 2009/Aprobación: Septiembre 9 de 2009
RESUMEN
Los animales afectan los patrones de ocupación de espacio de las plantas de los bosques tropicales a través de la dispersión que generan. Por lo tanto, cambios en las poblaciones de vertebrados por la intervención antrópica, pueden estar afectando las dinámicas de regeneración de las especies de plan tas. Se estudiaron las diferencias en la distribución espacial de plántulas y semillas del roble ( Quercus humboldtii) entre dos bosques andinos de los municipios de Charalá y Encino, Santander, con grados de intervención antrópica contrastantes. Se evaluó la densidad y distribución espacial de plántulas, en diferentes categorías de edad, en 490 y 484 parcelas de 1m2, en 28 ha por sitio. Las semillas se evaluaron en subparcelas de 0.25m2 ubicadas en las parcelas mencionadas. Los resultados muestran que en el sitio de mayor intervención se presenta una mayor densidad y número de plántulas, pero una marcada disminución en la densidad a medida que aumenta la edad. Las distancias observadas al vecino más cercano son considerablemente menores para todas las plántulas en el sitio de mayor intervención. Además, los índices de distribución espacial indican una distribución agregada, mien tras que en el sitio de menor intervención la distribución tiende a la uniformidad. De igual forma, en el sitio de mayor intervención se presentó una mayor agregación de semillas, así como una mayor depredación de las mismas por parte de invertebrados. Posiblemente, la fragmentación y por ende una menor presencia de animales vertebrados dispersores, depredadores y herbívoros asociados al proceso de regeneración de la especie, puede estar generando estos resultados. Esta información tiene implicaciones en el manejo de la restauración de los bosques naturales puesto que para garantizar la viabilidad de las poblaciones en el largo plazo se debe procurar la presencia permanente de vertebra dos en el bosque.
Palabras clave: andes colombianos, dispersión de semillas, distribución espacial, Reserva Biológica Cachalú, roedores caviomorfos.
ABSTRACT
Animals affect the spatial occupation patterns of tropical forest plants throughout the seed disper sal they perform. Therefore, changes in vertebrate populations by human disturbance might affect re generation dynamics of plant species. We studied differences in the spatial distribution of seeds and seedlings of the Andean oak (Quercus humboldtii) between two nearby forests with contrasting levels of anthropogenic influence in the Colombian Andes. Density and spatial distribution of seedlings were evaluated in 490 and 484 1 m2 plots located in a 28 ha area, in the high and low disturbed site, respectively. In each plot, all seedlings found were sampled and classified into three age categories. Density and spatial distribution of seeds were evaluated in 0.25m2 subplots placed in the same plots described above. Results showed a higher number and density of seedlings in the high disturbed site, as well as a marked decrease in seedling density as age increases. Distances to the nearest neighbour were shorter in the high disturbed site for all seedlings and each age category, in contrast to the low disturbed site. Indexes of spatial distribution indicate an aggregated pattern in the most disturbed site, while a uniform pattern in the low disturbed one. Seeds also exhibited an aggregated pattern in the high disturbed site and a higher seed predation by invertebrates. Results seemed to be a conse quence of forest fragmentation and a decrease of vertebrate seed dispersers, seed predators and her bivores associated to the regeneration processes of the species. These results provide important infor mation for the forest management and restoration activities, since in order to maintain plant popula tions in the long term, presence and viable populations of seed dispersers should be also maintained.
Key words: Colombian Andes, seed dispersal, spatial distribution, Cachalú Biological Reserve, caviomorph rodents.
RESUMO
Os animais afetam os padrões de ocupação do espaço das plantas dos bosques tropicais através da dispersão que geram. Portanto, mudanças nas populações de vertebrados pela intervenção antrópica, podem estar afetando as dinâmicas de regeneração das espécies de plantas. Estudaram-se as diferenças de distribuição espacial de mudas e sementes de roble (Quercus humboldtii) entre os bosques andinos dos municípios de Charalá e Encino, Santander, com graus de intervenção antrópica contrastantes. Avaliou-se a densidade e distribuição espacial das mudas, nas diferentes categorias de idade, em 490 e 484 lotes de 1m², em 28 há por lugar. As sementes se avaliaram em sublotes localizadas nos lotes mencionados. Os resultados mostram que o lugar de maior intervenção se apresenta uma maior densidade e número de mudas, mas uma marcada diminuição na densidade a medida que aumenta a idade. As distâncias observadas no vizinho mais perto são consideravelmente menores para todas as mudas no lugar de maior intervenção. Ademais, os índices de distribuição espacial indicam uma distribuição agregada, enquanto que no lugar de menor intervenção a distribuição tende a uniformidade. De igual maneira, no lugar de maior intervenção se apresentou uma maior agregação de sementes, assim como uma maior depredação das mesmas por parte dos invertebrados. Possivelmente, a fragmentação e por ende uma menor presença de animais vertebrados dispersores, depredadores e herbívoros associados ao processo de regeneração da espécie, pode estar gerando estes resultados. Esta informação tem implicações na manipulação da restauração dos bosques naturais posto que para garantir a viabilidade das populações a longo prazo se deve procurar a presença permanente de vertebrados no bosque.
Palavras chave: Andes colombianos, dispersão de sementes, distribuição espacial, Reserva Biológica Cachalú, roedores caviomorfos
INTRODUCCIÓN
La dispersión de semillas es un proceso que define el reclutamiento, rango de expansión, la demogra fía y la estructura espacial y genética de las poblaciones de plantas, especialmente en escalas locales e intermedias (Hubbell 1979, Nathan & Muller-Landau 2000, Parrado-Rosselli 2005, 2007). En los bosques tropicales, donde la mayoría de las especies de plantas presentan frutos y semillas adap tados para el consumo por animales (Snow 1981, Howe & Smallwood 1982, Arbelaéz & Parrado- Rosselli 2005), una gran cantidad de especies de plantas presenta una limitada o ineficaz dispersión de las semillas disponibles y una baja supervivencia de las semillas dispersadas y/o de plantas re cién establecidas por fenómenos denso-distancio dependientes (Janzen 1970). Por lo tanto, la disper sión de semillas que los animales desempeñan estaría siendo fundamental para explicar los patrones de distribución espacial y las dinámicas de regeneración de las poblaciones y comunidades de plan tas (Forget et al. 1998, Jordano & Schupp 2000, Bleher & Böhning-Gaese 2001, Parrado-Rosselli 2005, 2007). Por otro lado, ha sido sugerido que las intervenciones antrópicas como la fragmenta ción, la cacería, la tala, quema y la expansión de la agricultura, afectan la abundancia de importantes especies de aves y mamíferos dispersores de semillas (Kattán & Alvarez-López 1996, Cordeiro & Howe 2001, Wright 2003). En consecuencia, la densidad y la distribución espacial de semillas y plántulas de las especies de árboles dispersadas por animales, pueden verse afectadas por cambios en las poblaciones de animales frugívoros y dispersores (Wright 2003, Dirzo et al. 2007, Forget & Jansen 2007).
En los bosques andinos, uno de los ecosistemas de mayor importancia para realizar esfuerzos de conservación son los bosques de roble (Quercus humboldtii Bonpl. Fagaceae). A pesar de su no table disminución, aún se encuentran remanentes de estos bosques en las tres cordilleras (Etter & Wijngaarden 2000, Armenteras et al. 2003, Solano 2006). En el caso particular de la cordillera oriental, Armenteras et al. (2003) plantea que aún permanecen 128350 ha, pero sólo un 13%, se en cuentra bajo la figura de áreas protegidas tales como los Santuarios de Fauna y Flora de Iguaque (Boyacá) y Guanentá–Alto Río Fonce (Santander) y las reservas privadas como la Reserva Biológi ca Cachalú (Santander). Esta fragmentación y los diferentes procesos antropogénicos, han generado una simplificación estructural del bosque (Ocaña 2005) y variaciones en la abundancia y el compor tamiento de las poblaciones de animales dispersores (Otálora-Ardila 2003). Algunos autores han sugerido que, en estos sitios, posiblemente la fauna de las zonas más fragmentadas y más cercanas a los asentamientos humanos, se haya agotado o haya migrado a zonas más alejadas y con menos intervención (Otálora-Ardila 2003, Hidalgo-Jaramillo 2006, Vargas 2006).
En el género Quercus la dispersión primaria se pro duce cuando las bellotas caen por acción de la gravedad al llegar a la madurez. Un gran porcentaje de éstas no se logra distanciar del árbol parental (con excepción de árboles ubicados en zonas de lade ras con cierta pendiente) y por ende sufren una alta mortalidad debido a la acción de los depredadores (Herrera 1995, Barik et al. 1996). En contraste, un menor porcentaje de semillas suele ser consumido y dispersado de forma secundaria principalmente por roedores, quienes a pesar de ser depredadores dispersan por olvido las semillas, al enterrarlas en diferentes sitios para comérselas eventualmente en épocas de escasez (Smythe 1970, Pulido 2002, Gómez et al. 2008). Su eficiencia depende de su capacidad de recuperar y perder las semillas o de si están vivos para hacerlo (Van der Wall 2002). En el caso particular de los bosques de roble de la cordillera oriental, observaciones personales y algunos reportes (ver Vargas 2006) se han registrado las ardillas (Sciurus granatensis), los tinajos (Coniculus taczanowskii, C. paca) y picuros (Dasyprocta punctata) como sus principales dispersores/ depredadores de semillas. Por lo tanto, cambios en las poblaciones de estos dispersores secundarios que alejan, entierran y/o olvidan las semillas del Quercus en el bosque pueden afectar seriamente las dinámicas de reclutamiento de la especie.
Teniendo en cuenta que la fragmentación y la pérdida de fauna pueden generar impactos en la dispersión de las especies de plantas y por ende afectar sus distribuciones naturales ( e.g. Forget et al. 1998, Bleher & Böhning-Gaese 2001, Wright 2003, Terborgh et al. 2008), este estudio tiene como objetivo evaluar las diferencias en el patrón espacial de distribución de las plántulas y semillas del roble en dos bosques andinos de la cordillera oriental, de los municipios de Encino y Charalá (Santander) con grados de intervención antrópi ca contrastantes. Para tal fin, se caracterizaron las abundancias de semillas y plántulas en los dos sitios, se obtuvieron las distancias al vecino más cercano de semillas y plántulas y posteriormente se compararon y analizaron los patrones espaciales entre sitios. Se supondría, entonces, que para una especie común, altamente dependiente de animales para su dispersión como es el roble, se encontrará un patrón de distribución espacial de semillas y plántulas agregado y menores distancias al vecino más cercano en zonas de mayor perturbación, debido a la disminución de las poblaciones de ma míferos que cumplen la función de dispersores (Parrado-Rosselli 2005, Forget & Jansen 2007). En contraste, en los sitios menos intervenidos, con una mayor presencia de dispersores, se esperaría encontrar un patrón de distribución más espaciado por el bosque.
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
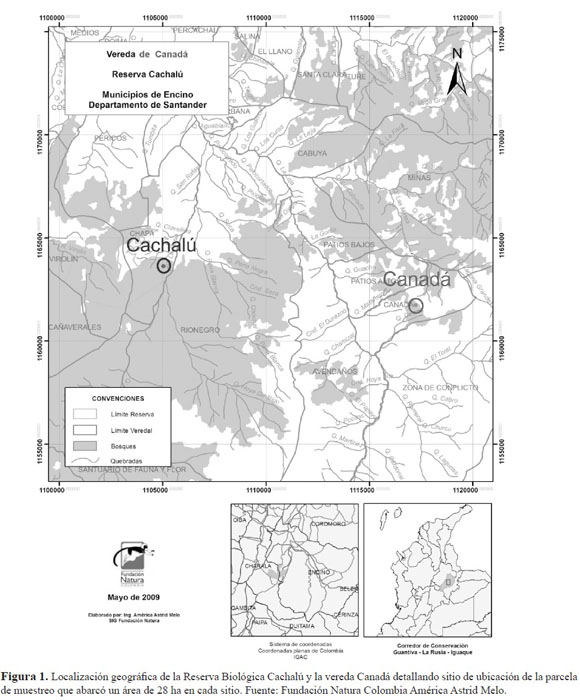
Esta investigación hace parte del mega-proyecto “Corredor de conservación de robles Guantivá – La Rusia – Iguaque: una estrategia para la conservación y el manejo forestal en Colombia” que ha rea lizado la Fundación Natura-Colombia en conjunto con el grupo Uso y Conservación de la Diversidad Forestal. Este tiene como fin generar información que apoye la formulación de planes de manejo para así generar estrategias para el aprovechamiento sostenible y la conservación del recurso forestal, los robles y sus ecosistemas asociados. Los sitios de estudio fueron la Reserva Biológica Cachalú y la vereda Canadá, localizadas entre los Municipios de Charalá y Encino, respectivamente, en el depar tamento de Santander. Los dos sitios se encuentran en la vertiente occidental de la cordillera oriental, en el corredor biológico de conservación Guantivá – La Rusia – Iguaque, ubicado en los departamen tos de Boyacá, Santander y Cundinamarca (Solano et al. 2006). Con base en la zonificación del corre dor, los dos sitios corresponden a Bosques Andinos Húmedos en Crestones Homoclinales (FhCh).
El sitio que se definió como el de menor intervención antropogénica es la Reserva Biológica Cacha lú. Ésta se encuentra ubicada entre los municipios de Charalá y Encino a 06º 05’ 19.0’’ N y 73º 07’ 56.5’’ O y ocupa un área de 1300 ha (Figura 1). Se encuentra en un gradiente altitudinal que va de los 1850 m a los 2750 m de altura. Presenta una precipitación media anual que supera los 3000 mm con una distribución bimodal (Cújar 2006). Este sitio es una Reserva de la Fundación Natura, creada con el ob jetivo de conservar los boques andinos y robledales allí presentes y hace parte de la zona amortiguadora del Santuario de Fauna y Flora Guanentá–Alto Río Fonce. El sitio ha sido protegido desde la creación del Santuario en 1993 y la posterior creación de la reserva en 1996. Además, se encuentra relativamen te alejado de veredas y asentamientos humanos. A pesar de la fragmentación de toda la zona, este sitio se caracteriza por presentar coberturas boscosas relativamente continuas y corresponde al relicto con tinuo más grande de bosque en la cordillera oriental (Otálora-Ardila 2003; Figura 1).
La cobertura vegetal corresponde a bosques subandinos y andinos primarios y secundarios con más de 20 años de regeneración y predominio del roble (Q. humboldtii) en asocio con especies de las familias Melastomataceae y Clusiaceae (Solano 2006, Avila et al. este volumen). Adicional al roble, las especies más importantes son los gaques Clusia spp., tunos Blakea spp y encenillos ( Weinmania spp; Solano 2006, Avila et al. este volumen). Según Avila, et al. (este volumen), la reserva presenta niveles de diversidad y de individuos por unidad de área similar a otros bosques andinos con presencia de roble. Adicionalmente, diferentes estudios han reportado que la fauna en estos bosques es abundante (Camargo & Vargas 2006, Otálora-Ardila & López-Arévalo 2006, Solano 2006). Según Otálora-Ardila (2003), debido a que la zona se encuentra en el fragmen to de mayor tamaño en la región, las densidades de mamíferos en la zona de la reserva son considera bles. Entre los principales mamíferos frugívoros se encuentran varias especies de murciélagos ( Carollia brevicauda, Artibeus lituratus y A. glaucus; Otálora-Ardila & López-Arévalo 2006), roedores como tinajos (Agouti taczanowskii, A. paca), picuros (Dasyprocta punctata), ardillas (Sciurus granatensis), aulladores (Alouatta seniculus), maiceros (Cebus albifrons), oso de anteojos (Tremarctos ornatus) y pequeños mamíferos tales como especies del género Akodon y Oryzomys (Alberico et al. 2000, Otálora-Ardila 2003, Vargas 2006,). También se encuentra una alta diversidad de aves con más de 250 especies (Cújar 2006), incluyendo importantes frugívoros como las familias Turdidae, Tyrannidae, Thraupidae y las pavas Aburría aburri y Penelope montagni (Camargo & Vargas 2006).
Por otro lado, el bosque ubicado en la vereda Canadá situada en el municipio de Encino (06º02’21,0’’N y 73º00’42.6’’ O), se consideró como el sitio con mayor grado de intervención (Figura 1). La zona se encuentra en un gradiente altitudinal entre los 2879 m y 3399 m, con una precipitación promedio anual de 3281 mm distribuidos en un régimen bimodal (Ocaña 2005). En la zona se presenta una mayor fragmentación del paisaje por sobreexplotación principalmente para actividades agrícolas (Figura 1) y no presenta ninguna figura de protección. Adicionalmente, ésta se encuentra permanentemente habitada por comunidades humanas. El fragmento en donde se realizó el estudio mantiene cierta con tinuidad con los demás fragmentos y cuenta con un área de aproximadamente 400 ha (Figura 1). La per turbación del sitio también se presenta a través de la extracción de productos del bosque por los cam pesinos de la zona, tales como la madera para usos domésticos y la cacería. Según los habitantes locales y observaciones personales, esta última comprende algunas especies como los tinajos y lapas o pa cas (Cuniculus taczanowskii, C. paca, Dasyprocta punctata) principalmente, aunque también se en cuentran los guaches, conejos y de manera menos frecuente venados, armadillos y puercoespines.
La cobertura vegetal corresponde a bosques andinos en donde predomina el roble (Q. humboldtii) en asocio con especies como gaques (Clusia spp.), encenillos (Weinmania spp.), mano de oso (Schefflera uribei), tunos (Miconia spp.) y yarumos (Cecropia telenitida; Ocaña 2005). Según Ocaña (2005) la composición y estructura de la vegetación en estos bosques sugiere un alto grado de intervención, que se manifiesta por la presencia de especies características de bosques secundarios y claros y por el bajo porcentaje de regeneración natural. Respecto a la fauna, según Otálora-Ardila (2003), en este sitio la riqueza de mamíferos medianos y grandes disminuye con el tamaño del fragmento, por lo que la presencia de especies de mamíferos es baja, estando completamente ausentes especies frugívoras como Tremarctos ornatus, Nasuella olivacea y Didelphis marsupialis. Hoy en día las especies más abundantes de mamíferos pertenecen a los géneros Akodon y Oryzomys. Las aves Aburria aburri y Penelope montagni han sido reportadas por ser especies bastante escasas y estar sometidas a una gran influencia de cacería (Vargas 2006).
TOMA DE DATOS
DISTRIBUCIÓN ESPACIAL DE PLÁNTULAS
En cada uno de los bosques previamente definidos, se instaló una grilla de 5 transectos de 700 m x 6 transectos de 400 m, distanciados regularmente cada 100 m, para un total de 5.9 km lineales de transecto en un área total de 28 ha por sitio. Sobre estos transectos se instalaron 490 parcelas de 1 m 2 en el sitio de menor intervención (Reserva Biológica Cachalú) y 484 parcelas de 1 m2 en el sitio de mayor intervención (Vereda Canadá). Las parcelas se ubicaron a través de un muestreo sistemático – aleatorio, en un rango de distancia de 875 – 15 m entre una y otra. De tal forma que un transecto contenía en promedio 44 parcelas. En cada parcela se registró el número total de plántulas de roble con altura < 1 m y diámetro de tallo < 1 cm (brinzales). Cada plántula se clasificó en categorías de edad A, B, o C, tomando como referencia una muestra previamente colectada de 40 plántulas. A éstas se les registró altura total, altura superficial y número de hojas. Las plántulas A se consideraron como aquellas recientemente establecidas y provenientes de la última estación de fructificación, con altura total promedio de 40.34 cm (rango 21 – 58 cm, n=23),una altura superficial promedio de 24 cm (rango 11 – 35 cm, n= 23) y un número de hojas promedio de 9.6 (rango 3 – 15 hojas, n = 23). Las plántulas B corresponden a aquellas con altura total promedio de 68.3 cm (rango 62 - 77 cm, n = 10), una altura superficial promedio de 43 cm (rango 38 - 52 cm, n = 10) y 19 hojas en promedio (rango 16 – 22 hojas, n = 10). Según observaciones personales y conocimiento local éstas pertenecen al mismo año y posiblemente a la misma estación de fructificación que las de categoría A, pero tienen más de tres meses de edad producto de la fructificación extendida del roble (> 5 meses sensu Newstrom et al. 1994, ver Pacheco & Pinzón 1997, Pulido 2002). Las plántulas C se consideraron como aquellas con una altura total promedio de 92 cm (rango 82 - 107cm, n=7), una altura superficial pro medio de 64 cm (rango 61 – 75 cm, n= 7) y 29 hojas en promedio (rango 24– 37, n=7), observaciones en campo sugieren que corresponden a la estación de fructificación anterior (> 1 año). Cada una de las ca tegorías de edad presentó diferencias significativas entre una y otra (análisis de varianza de una vía, N= 40 para altura total, F2.39 = 70.70; P< 0.05; altura superficial F2.39 = 90.26, P< 0.05; número de hojas F2.39 = 25.96, P< 0.05).
DISTRIBUCIÓN ESPACIAL DE SEMILLAS
En cada una de las parcelas de 1 m2 utilizadas para el muestreo de plántulas, se instaló una subparcela de 0.25 m2 para evaluar la densidad y la distribución de semillas. Las semillas encontradas en cada subparcela se separaron en viables y no viables, siendo estas últimas aquellas semillas abortadas, podridas o de destino indeterminado (sólo se encontraba el receptáculo). Cada semilla considerada como viable se clasificó de acuerdo a su estado de madurez y/o de ataque si estaban depredadas. En ese caso se registró la fuente de depredación (i.e. vertebrados, invertebrados –larvas y/o escaraba jos) observando la presencia de marcas de dientes, si estaban trituradas en pedazos o si exhibían larvas o pupas de insectos y/u orificio de salida. Con base en González-Melo & Parrado-Rosselli (en este vo lumen) la fructificación en los dos sitios durante el 2008 se presentó de marzo a julio. Por lo tanto, y puesto que el muestreo de las semillas en cada sitio se realizó durante un periodo de 1 mes, siendo marzo el mes de muestreo en el bosque de mayor intervención y abril en el de menor intervención, debe tenerse en cuenta que los datos analizados subestiman las semillas dispersadas en los dos si tios, principalmente en el bosque de mayor intervención, donde la producción de frutos hasta ahora estaba comenzando.
ANÁLISIS DE DATOS
Se determinó la abundancia total de plántulas, la densidad promedio y la distribución por categorías de edad por parcela y por sitio. De igual forma, se obtuvo la abundancia total y la densidad promedio de semillas en cada subparcela y en cada sitio. Se calculó el porcentaje de semillas potencialmente viables, que correspondieron al total encontrado menos aquellas que no llegaron a desarrollarse (abortadas, podridas) y aquellas sobre las que no se pudo determinar su destino (aparición del cas co vacío - receptáculo). De las semillas potencialmente viables se calculó el porcentaje de semillas depredadas tanto por vertebrados como por invertebrados. Se realizaron análisis de varianza de una vía para establecer si la abundancia y la densidad promedio de plántulas (total y por categorías de edad) presentó diferencias significativas entre sitios. También se utilizó para evaluar las diferencias entre la abundancia, densidad promedio y porcentaje de depredación de semillas entre sitios.
El análisis de distribución espacial se realizó a nivel de parcelas con plántulas y semillas, considerando cada parcela como unidad muestreal. En primer lugar, se calculó la proporción de parcelas que no presentaron plántulas o semillas por sitio estudiado. Las parcelas con plántulas y las subparcelas con semillas se graficaron en el programa AUTOCAD V. 14 (Autodesk Inc. 1999) para el posterior análisis de su distribución. Esta distribución se obtuvo en cada sitio para todas las parcelas en general y por categorías de edad (A, B y C). Para cada parcela, en cada uno de los si tios, se obtuvo la distancia observadas al vecino más cercano. Con base en Krebs (1989) y He et al. (1997) el patrón de distribución espacial de las subparcelas con semillas y las parcelas con plántulas se obtuvo a través de la comparación de las distancias observadas y esperadas al vecino más cercano a través del índice de Clark & Evans (1954), corregido para efecto borde a través del índice de Donnelly (1978). Este índice se basa en el cálculo de distancias al vecino más cercano para evaluar si estas se alejan de un patrón aleatorio tipo Poisson y a través de simulaciones experimentales se modifica para minimizar los efectos de borde (Donnelly 1978). Con base en He et al. (1997), el índice consiste en:
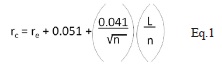
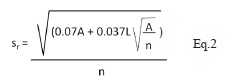
donde rc es la distancia esperada al vecino más cercano corregida para efecto borde; s r es el error estándar; re = 0.5/√ρ es la distancia esperada al ve cino más cercano sin considerar el efecto borde, siendo ρ la densidad de individuos (en este caso parcelas) del área (No. parcelas/m2); sr es el error estándar del estadístico de Donnelly; A es el área del plot (m2); L es la longitud del borde del área de estudio (m) y n es el número de parcelas en el área de estudio. El valor del estadístico de Donnelly se transforma en una desviación estándar normal Z por medio de la prueba-z:
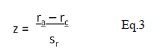
donde ra es la distancia media observada al vecino más cercano. Cuando |z| es menor que 1,96, el patrón es de uniformidad y cuando los valores son mayores el patrón es de agrupación. He et al. (1997) sugieren que el test-z es imparcial si el número de puntos es mayor a siete, sin importar si el área es rectangular, cuadrada o circular. Aunque la función K de Ripley es bastante utilizada para me dir la distribución espacial, esta parte de la suposición que los datos son homogéneos (estacionarios) o de lo contrario el patrón puede ser producto de variaciones en intensidad. Teniendo en cuenta que los datos de Cachalú no son homogéneos se optó por el índice de Clark y Evans, que según Condes & Martínez-Millán (1998) y He et al. 1997 es uno de los índices más eficientes en el caso de discrimi nar entre distribuciones aleatorias y distribuciones agregadas, si se logran minimizar los efectos de borde a través de la corrección de Donnelly.
RESULTADOS
PLÁNTULAS
Densidad
En el sitio de mayor intervención, correspondiente a la vereda Canadá, se encontró una mayor den sidad y un mayor número de plántulas total y por categorías de edad en relación al sitio de menor in tervención correspondiente a la Reserva Biológica Cachalú. Para ambos sitios se encontró una baja densidad promedio que en ningún caso superó el valor de 1 plántula/m2 (Tabla 1).
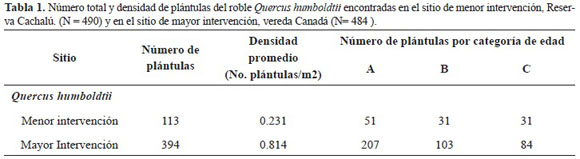
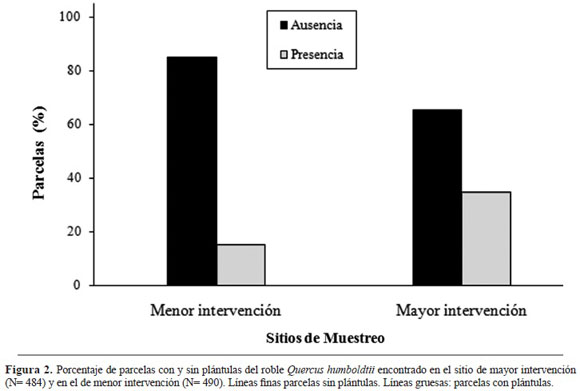
En cuanto a parcelas con plántulas por sitio estudiado, se encontró que en ambos sitios el porcenta je de parcelas con plántulas no fue superior al 40% (Figura 2). No obstante, el porcentaje de parcelas con plántulas en el bosque más intervenido supera en más del doble a lo encontrado en el de menor intervención. Al realizar un análisis de varianza de una vía se encontró que existen diferencias signi ficativas entre el número promedio de plántulas entre sitios de muestreo (ANOVA una vía F 1.966 = 52.76; N = 967 P< 0.05).
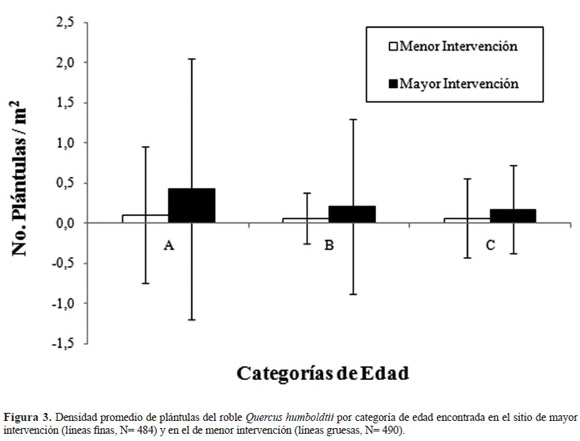
Con relación a las categorías de edad para plántulas, la densidad promedio por categoría de edad fue menor a 1 plántula/m2 (Figura 3). La densidad promedio fue mayor en el sitio de mayor interven ción para todas las categorías de edad. Se encontró que en ambos sitios, aunque la densidad promedio de plántulas/categoría de edad disminuye a medida que aumenta la edad, los cambios entre una cate goría a otra son más marcados en el sitio de mayor intervención (Figura 3). Sin embargo, las diferen cias no son significativas (p > 0.05).
Distrbución espacial
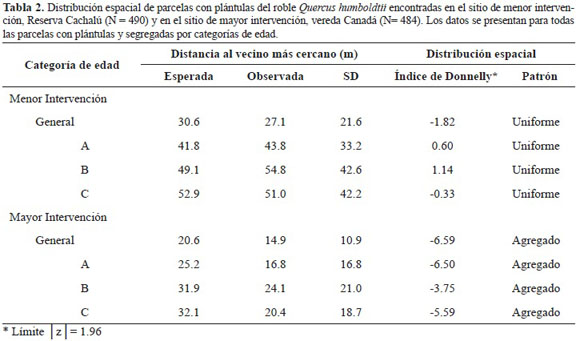
En el sitio de menor intervención, la distancia observada al vecino más cercano para las parcelas con plántulas A (43.8 m) y B (54.8 m) es mayor a la distancia esperada, lo que indica una tendencia a la uniformidad (Tabla 2). La distancia observada al vecino más cercano para todas las parcelas y para las parcelas con plántulas C es menor a la esperada, lo que indica cierta tendencia a la agregación. Al evaluar a través del índice de Donnelly (Krebs 1989) la distribución fue uniforme en todas las ca tegorías de plántulas (Tabla 2).
SEMILLAS
Densidad
Aunque, como se mencionó en los métodos, los datos analizados corresponden a sólo un mes de los cinco que dura la estación de fructificación, se encontró que, tanto en términos del total encontrado en las subparcelas como en términos de las semillas potencialmente viables, la abundancia de semillas en el sitio de mayor intervención fue significativamente mayor respecto al sitio de menor intervención (Tabla 3; F 1,972 = 10.84, N= 973 P < 0.05).

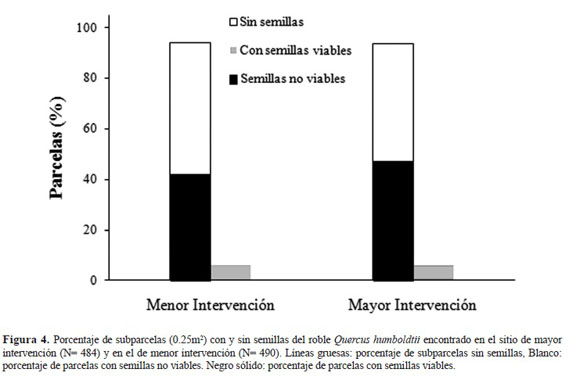
El porcentaje de subparcelas que presentaron semillas viables fue similar y muy bajo para ambos sitios, encontrándose valores por debajo del 10% del total muestreado (Figura 4). La presencia de subparcelas con semillas no viables así como sin semillas (ausencia) fue igualmente muy similar para ambos sitios.
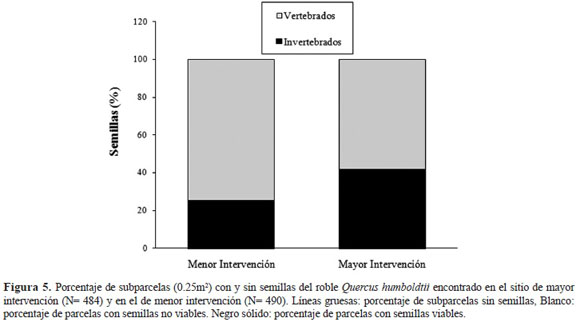
Respecto a la depredación de semillas, esta afectó a más del 90% del total de las semillas que fueron clasificadas como viables (menor intervención N = 315 semillas, mayor intervención N = 520 semi llas). La depredación fue producto de dos fuentes: invertebrados y vertebrados, siendo estos últimos los principales responsables de la misma. Los invertebrados pertenecieron a las familias Coleop tera y Lepidoptera, mientras que los depredadores vertebrados correspondieron a ardillas, loros, gua ches y lapas. Al comparar entre sitios, se encontró una mayor depredación de las semillas en el sitio de mayor intervención. En este sitio la depredación por invertebrados se duplica en relación con el si tio de menor intervención (Figura 5). Además, al comparar el número promedio de semillas/subpar cela que registraron depredación por invertebrados esta fue de 1.25 (semillas/subparcela) en el sitio de menor intervención, mientras que en el de mayor intervención fue de 2.15 (semillas/subparcela ).
Distribución espacial
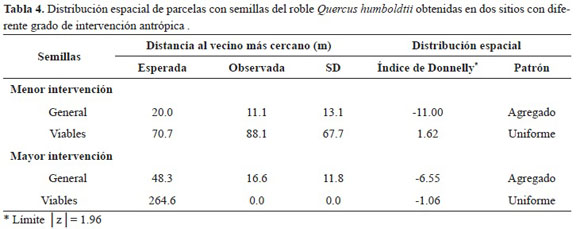
En el sitio de menor intervención, la distancia observada al vecino más cercano para todas las semi llas (viables y no viables) es menor que la distancia esperada (Tabla 4). Sin embargo, para las semillas viables la relación de la distancia observada (88.1 m) es mayor que la distancia esperada (79.7 m), lo que revela una tendencia a la uniformidad. Esto lo confirma el índice de Donnelly que muestra un patrón uniforme (Tabla 4).
En el sitio de mayor intervención, no fue posible evaluar las distancias al vecino más cercano, pues sólo se encontraron semillas viables, puesto que los datos no corresponden a la estación completa de fructificación. Por lo tanto, los resultados no son concluyentes. Al analizar las semillas en general (viables y no viables) la relación entre distancia observada a esperada en el sitio de mayor intervención indica una tendencia a la agregación (Tabla 4). No se presentaron diferencias significativas entre las distancias al vecino más cercano entre sitios (F 1, 335 = 25.63; N= 336 P < 0.05).
DISCUSIÓN DE RESULTADOS
Los resultados de este estudio apoyan la premisa de que la fragmentación y la perturbación an trópica afectan el reclutamiento de las plántulas (Laurance et al. 1998) así como la depredación de semillas. En un muestreo relativamente similar en dos sitios cercanos, el cual abarcó un área de 28 ha por sitio (490 vs 484 parcelas) se encontraron diferencias importantes tanto en la densidad como en la distribución espacial de plántulas y semillas del roble. Por un lado, respecto a las plántulas, que son el reflejo de la dispersión de semillas una vez han ocurrido los eventos de reorganización (dispersión primaria y secundaria), germinación y establecimiento (Hamill & Wright 1986, Bleher & Böhning-Gaese 2001), se encontró una mayor abundancia y agregación en el sitio de mayor intervención (vereda Canadá) en general y para cada una de las categorías de edad (A, B y C). Además, aunque en los dos sitios se encontró una marcada disminución en la densidad de plántulas entre categorías de edad, el patrón de distribución perma neció constante. Por otro lado, a pesar de que no se pudo muestrear toda la estación de fructificación, el sitio de mayor intervención presentó una mayor depredación de semillas y un incremento de la de predación por invertebrados.
Posiblemente, la fragmentación y por ende una menor presencia de animales vertebrados asociados al proceso de regeneración de la especie, pueden estar generando estos resultados. En este estudio, los prin cipales dispersores de las semillas del Q. humboldtii son mamíferos de suelo como tinajos y picuros (e.g. Cuniculus taczanowskii, C. paca, Dasyprocta punctata) los cuales pueden verse afectados tanto por eventos de cacería como de fragmentación, que generan la desaparición o el desplazamiento de los animales hacia el interior del bosque (Otálora-Ardi la 2003, Hidalgo-Jaramillo 2006, Vargas 2006). Debido a que estos vertebrados usualmente dispersan semillas a distancias iguales o mayores a 30 m del parental (Brewer 2001, Brewer & Webb 2001, The imer 2003, Ramírez et al. 2009), su ausencia estaría generando que las semillas, y por ende las plántulas, estuvieran menos distribuidas por el bosque y más cercanas entre sí. Adicionalmente, la densidad de plántulas tiende a ser más alta en el sitio de mayor intervención, pues ante la disminución de los verte brados herbívoros, el consumo de plántulas disminuye y aumenta la probabilidad de germinación y supervivencia de las mismas (Asquith et al. 1999, Cordeiro & Howe 2001, Wright 2003, Wyatt & Sil man 2004, Wang et al. 2006, Andresen & Laurance 2007).
En este contexto, aunque la vereda Canadá y la Reserva Biológica Cachalú están ubicadas en la cordillera oriental y pertenecen a la misma unidad de paisaje, los bosques de la vereda Canadá han sido sometidos a una mayor alteración, dada por la presencia permanente de comunidades humanas (campesinos) que ha generado una disminución de los vertebrados que cumplen papeles de dispersores de semillas, depredadores y herbívoros (Otálora-Ardila 2003, Hidalgo-Jaramillo 2006). En contras te, al estar en una Reserva de la Sociedad Civil, la fauna en los bosques de Cachalú se encuentra bajo protección. La zona se caracteriza por estar alejada de comunidades humanas, por presentar coberturas boscosas relativamente continuas y se ubica en el relicto continuo más grande de bosque en la cordi llera oriental (Otálora-Ardila 2003), en donde se encuentran las más altas densidades de mamíferos de toda la zona.
Una mayor frecuencia de claros y una menor competencia entre especies como consecuencia de una menor diversidad, también puede estar generando una mayor densidad de plántulas en el sitio de ma yor intervención. Ocaña (2005), encontró que en la zona de mayor intervención, se presenta una simplificación estructural que se manifiesta por la presencia de especies características de bosques se cundarios y claros. Por lo tanto, al ser el roble una especie de rápida germinación, fuerte competidora y con requerimientos intermedios de luz (Cárdenas et al. 2000), una simplificación estructural podría tener efectos positivos en la abundancia relativa de las plántulas. A pesar de esta mayor densidad de plántulas en el sitio de mayor intervención, se observa una marcada disminución de la densidad de una categoría de edad menor a la siguiente (Figura 3). Esto probablemente se debe a factores de mortalidad denso-distancio dependientes, ya que al existir una mayor agregación y por ende menores distancias entre una plántula y otra, las posibilidades de ataque por parte de depredadores vertebra dos e invertebrados, patógenos y mortalidad por competencia aumentan (Janzen 1970). Aunque es normal para las especies de plantas que a medida que aumenta la edad se presente una disminución en la densidad y en el espaciamiento por diferentes causas implícitas en el funcionamiento del bosque, la mayor depredación de semillas encontrada en el sitio de mayor intervención apoyaría esta hipótesis.
SEMILLAS
Respecto a la densidad y la distribución de semillas, los altos niveles de depredación, la baja dis ponibilidad de semillas viables y el corto periodo de muestreo (sólo un mes de los cinco que com prende la estación de fructificación), no permitieron obtener datos robustos sobre los patrones de distribución, ni evaluar la probabilidad de cambio de semillas a plántulas como ha sido realizado en otros estudios. A pesar de lo anterior, los datos muestran que el alto porcentaje de depredación de semillas en ambos sitios es similar a lo encontrado para otras especies de roble tropical (Barik et al. 1996). En el sitio de mayor intervención se encontró una mayor depredación de semillas, lo que podría ser consecuencia de un efecto denso-distancio dependiente (Janzen 1970), en dónde ma yores densidades generan mayores probabilidades de depredación. Adicionalmente, la depredación por vertebrados disminuyó en el bosque de mayor intervención, lo cual puede reflejar la ausencia de tinajos y picuros en el área, mientras que la depredación por invertebrados aumentó. Este patrón ha sido también encontrado para algunas especies de palmas en el Brasil y en Colombia (Bonjorne & Galetti 2007, Ramírez et al. 2009), en las que la depredación por invertebrados aumenta con la in tervención antropogénica. Posiblemente, esto se debe a que al presentarse una disminución en las poblaciones de vertebrados, puede generarse una compensación funcional, en donde vertebrados pe queños e invertebrados depredadores de semillas, comienzan a presentar incrementos compensa torios en abundancia (Asquith et al. 1997, 1999, Cordeiro & Howe 2001, Wright 2003, Wyatt & Sil man 2004, Wang et al. 2006, Andresen & Laurance 2007). Futuros estudios que abarquen la estación completa de fructificación y estudios acerca del destino de las semillas dispersadas por vertebrados e invertebrados, podrían confirmar estos hallazgos.
CONSIDERACIONES FINALES
En esta investigación se planteó un muestreo que permitió el estudio de la dispersión y sus conse cuencias a nivel de sitio. Esto contrasta con una gran cantidad de estudios (e.g. Kitajima & Augs purger 1989, Forget et al. 1999, Cordeiro & Howe 2001, Parrado-Rosselli 2005, 2007) que consideran el árbol parental como punto de partida y por ende se genera una restricción en las distancias máximas a abordar al tratar de garantizar que las plántulas muestreadas procedan del mismo parental (máxi mo 100 m de distancia desde la base). Esto limita desde el comienzo la óptica del estudio, pues no permite una evaluación a nivel de bosque o de la población, sino a nivel de árboles aislados. Otra desventaja de los muestreos en situaciones donde una sola especie es dominante, como en el caso del roble, es que establecer el parental más cercano no es posible a no ser que se utilicen técnicas genéticas/moleculares. Por lo tanto, con el fin de minimizar este tipo de sesgos y generar informa ción a nivel poblacional, se muestreó la población de plántulas y semillas distribuidas en el bosque, sin tener como referencia al parental, abarcando un área de 28 ha por sitio. Esto permitió conocer la distribución en todo el bosque, detallar la heterogeneidad del mismo y abordar el mayor número de lugares con probabilidad de llegada y colonización de nuevos propágulos.
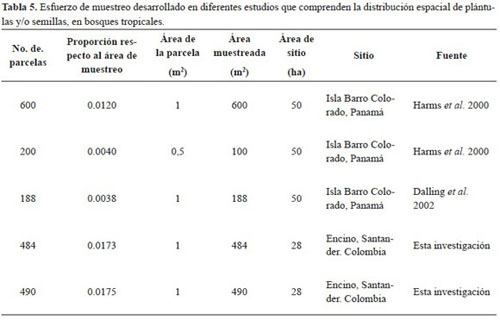
A nivel de poblaciones, este estudio comprende un gran esfuerzo de muestreo, al comparar con otros estudios realizados sobre densidad y distribución espacial de plántulas y semillas en bosques tropicales (Tabla 5). Por ejemplo, estudios que han utilizado el sistema de parcelas pequeñas para evaluar den sidad han utilizado un menor número de parcelas o una menor proporción respecto al área total. El con templar cada parcela de muestreo (1m2) como un individuo, permitió abarcar una gran área y a la vez facilitó el muestreo pues no sólo supone plántulas aisladas sino grupos de plántulas, por lo que se reco mienda este tipo de muestreo para futuros estudios. Sin embargo, se debe tener en cuenta que la toma de datos incluya por lo menos un periodo completo de fructificación. Se recomienda además, realizar una demarcación de los puntos muestreados que permita una posterior captura de datos y un efectivo segui miento, en el mediano y largo plazo, del desarrollo de los individuos referenciados.
Muchos autores han sugerido que dada la complejidad de las interacciones entre la pérdida de fauna y las poblaciones de plantas, los impactos pueden ser específicos a cada sitio y su biota asociada (Asquith et al. 1997, Guariguata et al. 2002, Wright 2003, Wyatt & Silman 2004). Aunque es claro que una gran cantidad de factores pueden estar involucrados en la regeneración del roble, los resultados aquí presentados muestran que la variación en la perturbación y las posibles variaciones en la biota asociada, tienen efectos importantes en la población de plántulas, la cual puede generar cambios en los juveniles y adultos y por ende cambiar considerablemente la composición y estructura de los bos ques andinos. Al determinar que la mayor densidad y agregación de las semillas y plántulas puede ser consecuencia de una disminución en vertebrados dispersores, depredadores y herbívoros por efecto de la perturbación, debe tenerse presente que para estrategias de restauración de los bosques ubicados en el corredor de conservación Guanentá – La Rusia – Iguaque se debe procurar la presencia permanente de vertebrados dispersores pues el sólo reintroducir especies vegetales no necesariamente garantizará la viabilidad y estructura del bosque en el largo plazo.
Finalmente, esta información, puede ser de gran importancia para entender el papel que desempe ñan los animales en las dinámicas de regeneración de los bosques y por tanto, anticipar los efectos de su desaparición. Esto, sin duda puede ser de gran utilidad para el establecimiento de criterios y es trategias de manejo y restauración, tanto para las especies de plantas como para las especies de ani males en los ecosistemas andinos tropicales. Además, el conocimiento de los patrones espaciales de dispersión resulta de gran importancia ya que determina no solamente el área potencial del reclu tamiento de las plantas, sino además otros procesos asociados como son la depredación, la competen cia y el establecimiento (Nathan & Muller-Landau 2000). Por lo tanto, ayuda a entender los factores que determinan la demografía y los patrones de ocupación de espacio de las especies de plantas, tema que actualmente es centro de debate en ecología tropical.
CONCLUSIONES Y RECOMENDACIONES
La abundancia y la distribución espacial de plántulas de Quercus humboldtii presentó diferencias entre los sitios, siendo superior la abundancia y la agregación en el bosque con mayor grado de intervención antrópica.
La distribución espacial de semillas coincidió con el patrón encontrado para las plántulas, presentan do una mayor agregación en el bosque de mayor intervención.
En el bosque más intervenido se presentó una mayor depredación de las semillas en relación al sitio de menor intervención. En este sitio la depredación por invertebrados aumentó, mientras que la depre dación por vertebrados disminuyó.
Es importante determinar otros efectos sobre las poblaciones de plántulas tales como dinámica de claros, requerimientos nutricionales, de luz, entre otros. También se sugiere estudiar más de un pe riodo de fructificación que incluya el seguimiento de los individuos en diferentes periodos de tiempo para determinar la tasa de cambio de una categoría de edad a la siguiente.
Para programas de restauración y conservación de los bosques de roble, se deben proponer estrategias de manejo de fauna, incluyendo conservación de las poblaciones existentes, reintroducción y meca nismos de conectividad, así como sistemas agroforestales que atraigan y mantengan las poblaciones de dispersores.
AGRADECIMIENTOS
Esta investigación estuvo financiada por la Fundación Natura Colombia y la fundación MacArthur en el marco del macroproyecto “Corredor de Conservación de Robles: una estrategia para la conser vación y el manejo forestal en Colombia” y por la Universidad Distrital Francisco José de Caldas. Agradecemos al personal de la reserva de Cachalú y a las familias de la vereda Canadá por su apoyo en la fase de campo. A Luís Mario Cárdenas por el apoyo en la realización de cada una de las fases de esta investigación. A América Astrid Melo por el mapa del área de estudio, a René López, a Edgar Cantillo y a Natalia Norden por sus comentarios y aportes en versiones previas de este manuscrito.
REFERENCIAS BIBLIOGRÁFICAS
Alberico, M., A. Cadena, J. Hernández-Camacho & Y. Muñoz-Saba. 2000. Mamíferos (Synapsida: Theria) de Colombia. Biota Colombiana 1: 43-75. [ Links ]
Andresen, E. & S.G.W. Laurance. 2007. Possible indirect effects of mammal hunting on dung beetle assemblages in Panama. Biotropica 39: 141-146. [ Links ]
Arbeláez, M.V. & A. Parrado-Rosselli. 2005. Seed dispersal modes of the vegetation on the sandstone plateaus of the middle Caquetá river region, Colombian Amazonia. Biotropica 37: 64-72. [ Links ]
Armenteras, D., F. Gast & H. Villareal. 2003. Andean forest fragmentation and the represen tativeness of protected natural areas in the eastern Andes, Colombia. Biological Conservation 113: 245-256. [ Links ]
Asquith, N.M., S.J. Wright & M.J. Clauss. 1997. Does mammal community composition control recruitment in neotropical forests? Evidence from Panama. Ecology 78: 941-946. [ Links ]
Asquith, N.M., J. Terborgh, A.E. Arnold & C.M. Riveros. 1999. The fruits the agouti ate: Hymenaea courbaril seed fate when its dis perser is absent. Journal of Tropical Ecology 15: 229-235. [ Links ]
Barik, S.K., R.S. Tripathi, H.N. Pandey & P. Rao. 1996. Tree regeneration in a subtropical humid forest: effect of cultural disturbance on seed production, dispersal and germination. Journal of Applied Ecology 33: 1551-1560. [ Links ]
Bleher, B. & K. Böhning-Gaese. 2001. Consequences of frugivore diversity for seed dis persal, seedling establishment and the spatial patterns of seedlings and trees. Oecologia 129: 385-394. [ Links ]
Bonjorne, L. & M. Galetti. 2007. Seed dispersal and spatial distribution of Attalea geraensis (Arecaceae) in two remnants of Cerrado in Sotueastern Brazil. Acta Oecologica 32: 180-187. [ Links ]
Brewer, S.W. 2001. Predation and dispersal of large and small seeds of a tropical palm. Oikos 92: 245-255. [ Links ]
Brewer, S.W. & M.A. Webb. 2001. Ignorant seed predators and factors affecting the seed survival of a tropical palm. Oikos 93: 32-41. [ Links ]
Camargo, C. & S. Vargas. 2006. La relación dispersor-planta de aves frugívoras en zonas sucesionales tempranas como parte de la restauración natural del bosque andino, pp.: 157-172. En: Solano, C. & N. Vargas. (eds.) Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Fundación Natura-Pontificia Universidad Javeriana Bogotá [ Links ]
Cárdenas, F., H. Cordero, C.A. Devia, H. Arenas, D. Díaz. 2000. Desarrollo Sostenible en los Andes de Colombia. Provincias del Norte, Gutiérrez y Valderrama (Boyacá, Colombia). Pontificia Universidad Javeriana-Unión Europea. Bogotá. 301 p [ Links ]
Clark, P.J. & F.C. Evans. 1954. Distance to nearest neighbor as a measure of spatial relations hips in populations. Ecology 35: 445-453. [ Links ]
Condes, S. & J. Martínez-Millán. 1998. Comparación entre los índices de distribución espacial de árboles más usados en el ámbito forestal. Investigación Agraria. Sistemas y Recursos Forestales 7: 173-187. [ Links ]
Cordeiro, N.J. & H.F. Howe. 2001. Low recruitment of trees dispersed by animals in African forest fragments. Conservation Biology 15: 1733-1741. [ Links ]
Cújar, A. 2006. Uso del hábitat del venado (Mazama rufina) en la Reserva Biológica Cachalú y su área de influencia, pp. 101-117. En: Vargas, W. & C. Solano. (eds.) I Simposio Internacional del Roble y Ecosistemas Asociados. Fundación Natura Bogotá [ Links ]
Dalling, J.W., H.C. Muller-Landau, S.J. Wright & S.P. Hubbell. 2002. Role of dispersal in the recruitment limitation of neotropical pioneer species. Journal of Ecology 90: 714-727. [ Links ]
Dirzo, R., E. Mendoza & P. Ortíz. 2007. Size-related differential seed predation in a heavily defaunated neotropical rain forest. Biotropica 39: 355-362. [ Links ]
Donnelly, K.P. 1978. Simulations to determine the variance and edge effect of total nearest-neigh bor distance, pp: 91-95. En: Hodder, I. (ed.). Simulation Studies in Archaeology. Cambridge University Press Cambridge London [ Links ]
Etter, A. & W. vanWyngaarden. 2000. Patterns of Landscape Transformation in Colombia, with Emphasis in the Andean Region. Ambio 29: 432-439. [ Links ]
Forget, P.M., T. Milleron & F. Feer. 1998. Patterns in post-dispersal seed removal by neotropical rodents and seedfate in relation to seed size. pp. 25-49. En: Newberry, D.M., Prins, H.H.T. & Brown, N.D. (eds.) Dynamics of Tropical Communities. Blackwell Science. Oxford. [ Links ]
Forget, P.M., K. Kitajima & R.B. Foster. 1999. Pre- and post-dispersal seed predation in Tachigali versicolor (Caesalpiniaceae): effects of timing of fruiting and variation among trees. Journal of Tropical Ecology 15: 61-81. [ Links ]
Forget, P.M. & P.A. Jansen. 2007. Hunting Increases Dispersal Limitation in the Tree Carapa procera, a Nontimber Forest Product. Conservation Biology 21: 106-113. [ Links ]
Gómez, J.M., C. Puerta-Piñero & E.W. Shupp. 2008. Effectiveness of rodents as local seed dispersers of Holm oaks. Oecologia 155: 529-537. [ Links ]
Guariguata, M., H. Arias-Le Claire & G. Jones. 2002. Tree seed fate in a logged and fragmen ted forest landscape, Northeastern Costa Rica. Biotropica 34: 405-415. [ Links ]
Hamill, D. & S.J. Wright. 1986. Testing the dispersion of juveniles relative to adults: a new analytic method. Ecology 67: 952-957. [ Links ]
Harms, K.E., S.J. Wright, O. Calderón, A. Hernández & E.A. Herre. 2000. Pervasive density dependent recruitment enhances seedling diversity in a tropical forest. Nature 404: 493-495. [ Links ]
He, F., P. Legendre & J.V. LaFrankie. 1997. Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science 8: 105-114. [ Links ]
Herrera, J. 1995. Acorn predation and seedling production in a low-density population of Cork oak (Quercus suber L). Forest Ecology and Management 76: 197-201. [ Links ]
Hidalgo-Jaramillo, F. 2006. Evaluación de los factores que inciden en la sosteniblidad de la cacería de subsistencia en el municipio de Encino, Santander, Colombia, pp: 235-258. En: Solano, C. & N. Vargas. (eds.) Memorias del I Simposio Internacional de Robles y Ecosiste mas Asociados. Fundación Natura - Pontificia Universidad Javeriana Bogotá [ Links ]
Howe, H.F. & J. Smallwood. 1982. Ecology of seed dispersal. Annual Review of Ecology and Systematics 13: 201-228. [ Links ]
Hubbell, S.P. 1979. Tree dispersion, abundance and diversity in a tropical dry forest. Science 203: 1299-1309. [ Links ]
Janzen, D. 1970. Herbivores and the number of tree species in tropical forests. The American Naturalist 104: 501-528. [ Links ]
Jordano, P. & E.W. Schupp. 2000. Seed disperser effectiveness: The quantity component and patterns of seed rain for Prunus mahaleb. Ecological Monographs 70: 591-615. [ Links ]
Kattan, G. & H. Alvarez-López. 1996. Preservation and management of biodiversity in frag mented landscapes in the Colombian Andes, pp: 3-18. En: Schelhas, J. & R. Greenberg. (eds.) Forest Patches in Tropical Landscapes. Island Press Washington D.C [ Links ]
Kitajima, K. & C.K. Augspurger. 1989. Seed and seedling ecology of a monocarpic tropical tree, Tachigali versicolor. Ecology 70: 1102-1114. [ Links ]
Krebs, C. 1989. Ecological methodology. Harper & Row Publishers. New York. [ Links ]
Laurance, W.F., L.V. Ferreira, J.M. Rankin-De Merona, S.G. Laurance, R.W. Hutchings, T. E. Lovejoy. 1998. Effects of forest fragmentation on recruitment patterns in Amazonian tree communities. Conservation Biology 12: 460-464. [ Links ]
Nathan, R. & H.C. Muller-Landau. 2000. Spatial patterns of seed dispersal, their deter minants and consequences for recruitment. Trends in Ecology & Evolution 15: 278-285. [ Links ]
Newstrom, L.E., G.W. Frankie & H.G. Baker. 1994. A new classification for plant phenology based on flowering patterns in lowland tropical rain forest trees at La Selva, Costa Rica. Biotropica 26: 141-159. [ Links ]
Ocaña, Y.G. 2005. Caracterización florística y es tructural de unidades de bosque altoandino en las veredas de Minas y Patios Altos, Encino, Santander. Colombia Forestal 9: 70-86. [ Links ]
Otálora-Ardila, A. 2003. Mamíferos de los bosques de roble. Acta Biológica Colombiana 8: 57-71. [ Links ]
Otálora-Ardila, A. & H.F. López-Arévalo. 2006. Incidencia de algunos elementos del paisaje fragmentado de Encino (Santander, Colombia) sobre la riqueza y diversidad de murciélagos, pp. 83-94. En: Solano, C. & N. Vargas. (eds.) Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Fundación Natura - Pontificia Universidad Javeriana Bogotá [ Links ]
Pacheco, R. & C. Pinzón. 1997. El roble Quercus humboldtii. Notas Divulgativas. Jardín Botánico de Bogotá José Celestino Mutis. Bogotá. [ Links ]
Parrado-Rosselli, A. 2005. Fruit availability and seed dispersal in terra firme rain forests of Colombian Amazonia. Tropenbos Ph.D. Series 2. Tropenbos-International. Wageningen. [ Links ]
Parrado-Rosselli, A. 2007. La dispersión de semillas: una herramienta para comprender la composición y estructura de los bosques amazónicos, pp. 109-116. En Ruiz, S.L., E. Sánchez, E. Tabares, A. Prieto, J. C. Arias, R. Gómez, D. Castellanos, P. García & L. Rodríguez (eds.). Diversidad biológica y cultural del sur de la Amazonia Colombiana - Diagnóstico. CorpoAmazonia, Instituto Alexander von Humboldt, Instituto SINCHI, UAESPNN Bogotá [ Links ]
Pulido, F.J. 2002. Biología reproductiva y conservación: el caso de la regeneración de bosques templados y subtropicales de robles (Quercus spp.). Revista Chilena de Historia Natural 75: 5-15. [ Links ]
Ramírez, B.H., A. Parrado-Rosselli & P.R. Stevenson. 2009. Seed dispersal of a useful palm (Astrocaryum chambira Burret) in three Amazonian forests with different human inter vention. Colombia Forestal 12: 5-16. [ Links ]
Smythe, N. 1970. Relationships between fruiting seasons and seed dispersal methods in a neo tropical rain forest. American Naturalist 104: 25-35. [ Links ]
Snow, D. 1981. Tropical frugivorous birds and their food plants: A world survey. Biotropica 13(1): 1-14. [ Links ]
Solano, C. 2006. Reserva Biológica Cachalú, pp. 11-13. En: Vargas, W. & C. Solano. (eds.) I Simposio Internacional del Roble y Ecosistemas Asociados. Fundación Natura Bogotá [ Links ]
Solano, C., C. Roa & Z. Calle. (eds.) 2006. Estrategia de desarrollo sostenible Corredor de Conservación Guantiva - La Rusia - Iguaque. Fundación Natura y The Nature Conservancy. Bogotá. 92 p. [ Links ]
Sork, V.L., J. Bramble & O. Sexton. 1993. Ecology of mast fruiting in three species of Nor th American decidious oaks. Ecology 74: 528-541. [ Links ]
Theimer, T.C. 2003. Intraspecific variation in seed size affects scatter-hoarding behaviour of an Australian tropical rainforest rodent. Journal of Tropical Ecology 19: 95-98. [ Links ]
Terborgh, J., G. Nuñez-Iturri, C.A. Nigel-Pitman, F.H. Cornejo-Valverde, P. Alvarez, V. Swamy, E. G. Pringle & C. E. Timothy Paine. 2008. Tree recruitment in an empty forest. Ecology 89: 1757-1768. [ Links ]
Van der Wall, S.B. 2002. Secondary dispersal of Jeffrey pine seeds by rodent scatter-hoarders: the roles of pilfering, recaching and a variable environment, pp. 193-208. En: Levey, D., J.W.R. Silva. & M. Galetti. (eds.). Seed dispersal and frugivory: ecology, evolution, and con servation. CABI Publishing Willingford UK [ Links ]
Vargas, N. 2006. El manejo colectivo de la fauna silvestre en bosques andinos: sus limitaciones y potencialidades, pp.: 183-202. En: Solano, C. & N. Vargas. (eds.)Memorias del I Simposio Internacional de Robles y Ecosistemas Asociados. Bogotá: Fundación Natura -Pontificia Universidad Javeriana [ Links ]
Wang, B.C., M.T. Leong, T.B. Smith & V.L. Sork. 2006. Ecological repercussions of extir pating mammals: reduced seed removal and dispersal of the Afrotropical tree, Antrocaryon klaineanum (Anacardiaceae). Biotropica 38: 792-793. [ Links ]
Wright, S.J. 2003. The myriad effects of hunting for vertebrates and plants in tropical forests. Perspectives in Plant Ecology, Evolution and Systematics 6: 73-86. [ Links ]
Wyatt, J.L. & M.R. Silman. 2004. Distance-dependence in two Amazonian palms: effects of spatial and temporal variation in seed predator communities. Oecologia 140: 26-35. [ Links ]
SOFTWARE: Autodesk INC. 1999. AutoCAD® version 14. [ Links ]

