











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los bosques secos tropicales (BST) están entre los ecosistemas más amenazados a nivel mundial. En el neotrópico, donde se encuentra más del 60 % de ellos (Portillo-Quintero & Sánchez-Azofeifa, 2010; Ocón et al., 2021), se estima que se han perdido al menos dos tercios de su extensión original (Miles et al., 2006; Riggio et al., 2020). Las causas de su deterioro incluyen factores como la deforestación, el cambio climático, el crecimiento demográfico y la conversión de tierras para la agricultura y la ganadería (Miles et al., 2006). Sumado a esto está la falta de políticas de protección efectiva; tan solo un 4.5 % del área correspondiente a BST está en zonas protegidas declaradas en todo el neotrópico, cifra por debajo de la de otros bosques tropicales (Portillo-Quintero & Sánchez-Azofeifa, 2010; Scharlemann et al., 2010).
Las características climáticas particulares que definen a los BST (precipitaciones anuales promedio de 250 a 2000 mm, elevada evapotranspiración y temperaturas mayores a los 17 °C) (Holdridge, 1982; Murphy & Lugo, 1995), sumado a su marcada estacionalidad -con temporadas secas que pueden llegar a durar entre 4 y 7 meses- los convierten en un entorno único, donde las especies han tenido que adaptarse de maneras particulares para sobrevivir (Janzen, 1988; Mooney et al., 1995). Las condiciones de estrés hídrico a las cuales se somete la vegetación han hecho que desarrollen patrones ecofisiológicos variados, por lo que gran parte de las especies pierden sus hojas durante la temporada seca, mientras que otras (de hoja perenne) invierten mayores recursos en la construcción de tejidos resistentes a la cavitación (Pennington et al., 2009; Romero-Duque et al., 2019; González et al., 2021). Dichas adaptaciones crean una diversidad florística única, que presenta altos niveles de endemismo (Kalascka et al., 2004; Linares-Palomino et al., 2011).
En Colombia, el estado de conservación de los BST es crítico. Originalmente, su extensión en el país comprendía cerca de 9 millones de ha, distribuidas en la región Caribe, los valles interandinos de los ríos Cauca y Magdalena, algunas zonas al norte de la cordillera Oriental, las cuencas de los ríos Patía y Dagua, y parte de la Orinoquía (Etter, 1993, Etter et al., 2008; Pizano et al., 2014). Sin embargo, actualmente solo se conserva el 8 % de ellos, muchos en diferentes estados de transformación y fragmentación (García et al., 2014).
Por su parte, la cuenca alta del río Magdalena conserva la segunda mayor extensión de BST del país, con poco más de 150 mil ha de bosques transformados y naturales (García et al., 2014; Vargas, 2015). Esta región conserva además más del 50 % de la flora registrada para este tipo de bosques en Colombia (Pizano et al., 2014). A pesar de ello, gran parte de las colecciones botánicas registradas para BST provienen de la región Caribe y del valle del río Cauca (Pizano & García, 2014), siendo comparativamente pocos los estudios de flora que se han publicado sobre esta región (Mendoza, 1999; Figueroa & Galeano, 2007; Fernández-Méndez et al., 2013; López et al., 2019; Romero-Duque et al., 2019).
Una gran parte de los BST que aún se conservan en la cuenca alta del río Magdalena corresponde a bosques riparios (Echeverry & Rodríguez, 2006; Rodríguez & López, 2014), ecosistemas terrestres asociados a sistemas lóticos que se extienden desde el borde de las corrientes de agua hasta los límites de las zonas de inundación (Gregory et al., 1991). Los bosques riparios son fundamentales para la preservación de los ecosistemas acuáticos, ya que funcionan como áreas de amortiguación que preservan las condiciones microclimáticas y químicas del agua, protegiéndola de agentes externos, evitando los procesos de erosión de los márgenes de las corrientes y estabilizando los lechos de los ríos (Gregory et al., 1991; Lowrance et al., 1997; Naiman et al., 2005). De igual forma, debido a la mayor disponibilidad de agua, los bosques riparios son elementos clave en la ecología de los BST, pues albergan una mayor diversidad de especies en comparación con sistemas no riparios y actúan como corredores biológicos en paisajes fragmentados (Murphy & Lugo, 1986; Naiman et al., 2000; Miserendino, 2004; Miles et al., 2006).
A pesar de su importancia en la ecología y conservación de los BST, los bosques riparios de la cuenca alta del río Magdalena también han enfrentado presiones antrópicas particulares, como la expansión de la frontera agropecuaria, las actividades turísticas no controladas y la construcción de grandes hidroeléctricas que han modificado su extensión, estructura y composición en la región (Ingetec, 2008; Vargas, 2015; Díaz, 2016). Por lo anterior, el objetivo de este estudio fue determinar la diversidad y estructura florística de un bosque ripario ubicado a lo largo de un arroyo dentro de un remanente del BST en el municipio de Paicol, Huila, Colombia. Además, se evaluó el grado de conservación del ecosistema y el impacto de las actividades antrópicas recientes sobre el mismo.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en el arroyo La Avería, ubicado en el municipio de Paicol en el departamento del Huila, en la cuenca alta del río Magdalena. El área de estudio presenta temperaturas entre los 23 y 28 °C, con un promedio anual de 23.4 °C. La precipitación anual promedio es de 1494 mm bajo un régimen climático monomodal, con una estación lluviosa que va de octubre a mayo y una estación seca entre junio y septiembre (IDEAM, 2005).
El arroyo tiene una longitud aproximada de 4.5 km y es un sistema de tercer orden perteneciente a la mesocuenca del río Páez en el suroccidente del Huila. El flujo de agua nace a una altitud de 922 m dentro de un remanente conservado de bosque ripario nativo. Luego pasa por distintas áreas donde ha sido perturbado por actividades humanas, como vertimientos urbanos, cultivos de arroz y ganadería bovina, para finalmente desembocar en el río Páez a una altitud de 805 m. Se destaca el hecho de que este sistema fluvial es de gran importancia para los habitantes del municipio, pues de este se realiza la captación de agua para abastecer al acueducto municipal y las fincas cercanas (Díaz, 2016; Peralta-Muñoz, 2016).
Toma de datos
Se establecieron tres estaciones de muestreo como representación de todo el arroyo. Estas estaciones, separadas por una distancia aproximada de 1.5 km, fueron definidas de acuerdo con cambios apreciables en la vegetación riparia, consecuencia de los diferentes procesos de degradación a lo largo del arroyo. La Estación 1 (2° 26' 22.3'' N 75° 46' 31.4'' W ± 922 m), ubicada en la zona alta del arroyo, corresponde a varios parches conservados de bosques interconectados. La Estación 2 (2° 26' 31.8'' N 75° 46' 5.9'' W ± 813 m), en la parte media del arroyo, corresponde a bosques riparios fuertemente degradados, resultado de la presencia de cultivos de arroz y la descarga de aguas residuales provenientes del casco urbano del municipio de Paicol, el cual está ubicado a menos de 350 m. Finalmente, la Estación 3 (2° 26' 57.2'' N 75° 45' 24.6'' W ± 805 m) está en la parte baja del arroyo, con un bosque ripario afectado por áreas ganaderas en las cercanías del sistema fluvial.
En cada estación de muestreo se establecieron tres transectos de 30 x 2 m a cada lado del arroyo y perpendiculares al curso de agua, para un total de seis transectos por estación, con un área de 0.036 ha y un total de área muestreada de 0.108 ha. Debido a la orografía y a los obstáculos en la zona de estudio, las distancias entre transectos no fueron constantes, pero se aseguró que tuvieran al menos 10 m de separación entre ellos. Este diseño se hizo siguiendo la metodología de Godfrey (1988), utilizada para estudiar la vegetación en condiciones bajo las cuales los bosques riparios se encuentran reducidos debido a la presencia de actividades o asentamientos humanos, lo que impide establecer transectos de una extensión mayor. Se tomaron muestras botánicas y datos de altura de todos los individuos con un diámetro a la altura del pecho (DAP) ≥ 2.5 cm.
Las muestras recolectadas fueron herborizadas y depositadas bajo la numeración de Nossa-Silva en el Herbario de Botánica Económica de la Universidad Militar Nueva Granada (UMNG-H). Para su identificación se utilizaron guías taxonómicas (Gentry, 1993; Romero-Hernández, 2017), listados de especies (Mendoza-C, 1999; Pizano et al., 2014; Vargas, 2015) y comparación con ejemplares de varios herbarios. La escritura correcta de los nombres científicos se verificó con las plataformas Trópicos (2022) y el Catálogo de Plantas y Líquenes de Colombia (Bernal et al., 2019).
Análisis de los datos
Para comprobar la efectividad del esfuerzo de muestreo se utilizó el método de rarefacción con números de Hill, según lo propuesto por Chao et al. (2014): N0 (riqueza de especies), N1 (número efectivo de especies comunes, eH’) y N2 (número efectivo de especies dominantes, 1/D). Los índices de diversidad de Shannon (H’) y de Simpson (1-D) para cada una de las estaciones se calcularon con el software PAST, versión 4.0.2. (Hammer et al., 2001) y para el cálculo de la rarefacción se utilizó el software EstimateS, versión 9.1.0. (Colwell et al., 2012).
Para los análisis estructurales, se calcularon el índice de valor de importancia (IVI) y el área basal para todas las especies de la comunidad de plantas. La determinación de estos valores se realizó siguiendo la metodología propuesta por Rangel-Ch y Velázquez (1997), con modificaciones para ajustar el valor máximo del IVI. Adicionalmente, se establecieron clases diamétricas y de altura, tomando en cuenta los valores máximos y mínimos en relación con el número de individuos, siguiendo la metodología de Lema (1995).
Para comparar las condiciones del bosque ripario entre las estaciones de muestreo, se calcularon tres variables ambientales en cada una de las estaciones establecidas. En primer lugar, se calculó el índice de calidad de bosques riparios (QBR), utilizando una metodología adaptada para Colombia (Munné et al., 2003). El índice QBR genera un valor cuantitativo en una escala de 0 a 100, donde un mayor número representa un mayor estado de conservación de la calidad del hábitat. Para complementar la información, se calcularon el coeficiente de densidad forestal (CDF) mediante el uso de un densiómetro de espejo convexo (Galeano-Rendón et al., 2017; Munné et al., 2003; Suárez et al., 2002) y la entrada de luz solar (Watts. m-2) por medio de un luxómetro (LI-COR LI-210R®).
RESULTADOS
Se registró un total de 199 individuos, repartidos en 26 familias, 42 géneros y 47 especies. La familia Fabaceae tuvo el mayor número de especies (9), seguida de Malvaceae (4), Euphorbiaceae, Salicaceae y Urticaceae, cada una con 3 especies. En términos de abundancia, el taxón más abundante fue Guadua angustifolia Kunth con 59 individuos, seguida de Casearia sylvestris Sw. con 22 y Zygia longifolia (Willd.) Britton y Rose con 14. De las especies encontradas, 24 (51.1 %) registraron un solo individuo en toda el área muestreada.
Las curvas de rarefacción de especies (Figura 1) muestran que los valores más altos fueron obtenidos por N0 (riqueza de especies), seguido de N1 (número de especies frecuentes) y, por último, N2 (número de especies muy frecuentes). Se observó que la curva de riqueza de especies continuaba aumentando, mientras que las curvas que representan N1 y N2 crecieron considerablemente solo al comienzo y, al llegar a la extrapolación, se estabilizaron en ambos casos. Lo anterior indica que, si bien en términos de riqueza aún podría aumentarse el esfuerzo de muestreo, N1 y N2 muestran un crecimiento limitado frente a más muestreos y se estabilizan antes de la extrapolación.
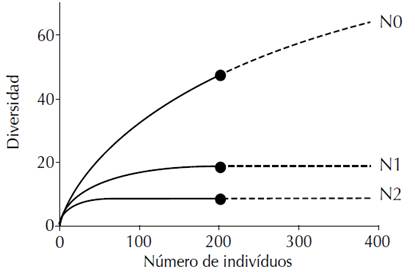
Figura 1 Curvas de rarefacción basadas en el tamaño de la muestra (líneas continuas) y extrapolación (líneas punteadas, hasta el doble del tamaño de la muestra) en función de los números de Hill (N0, N1 y N2) para el muestreo de la vegetación hecho en un BST ripario asociado al arroyo La Avería en la cuenca alta del río Magdalena, municipio de Paicol, Huila, Colombia
Con respecto a la diversidad, para la Estación 1 se reportaron los valores más bajos, con 16 especies, un índice de Shannon de 1.81 y diversidad de Simpson de 0.69. En contraste, la Estación 2 tuvo los valores más altos, con 23 especies, un índice de Shannon de 2.94 y diversidad de Simpson de 0.94. Finalmente, la Estación 3 mostró valores menores pero cercanos a los de la Estación 2, con 17 especies, un índice de Shannon de 2.29 y diversidad de Simpson de 0.82.
La especie con el de IVI más alto fue Z. longifolia, seguida por G. angustifolia y C. sylvestris (Tabla 1). Solamente Z. longifolia, C. sylvestris y Astronium graveolens Jacq. fueron encontradas en las tres estaciones muestreadas, mientras que 41 especies (87.2 %) fueron encontradas solo en una de las estaciones de muestreo. En cuanto a la dominancia, la especie con mayor área basal fue Z. longifolia, que contribuyó con el 51.57 % del área basal total (Tabla 1). Otras especies con valores altos de dominancia fueron Ficus insipida Willd. (12.59 %), Pithecellobium lanceolatum (Willd.) Benth. (6.82 %) y G. angustifolia (5.33 %), destacando que el valor de F. insipida proviene de un único individuo de gran porte registrado en la Estación 3 (Tabla 1). El área basal total para el bosque ripario fue de 16.06 m2. ha-1 (Tabla 1).
Tabla 1 Valores de área basal y ocurrencia de las especies con mayores valores del Índice de Valor de Importancia (IVI) reportados para un BST ripario asociado al arroyo La Avería en la cuenca alta del río Magdalena, municipio de Paicol, Huila, Colombia. Estación 1 (S1), Estación (S2), Estación (S3).
| Especie | IVI | Área Basal (m2.a-1) | S1 | S2 | S3 |
|---|---|---|---|---|---|
| Zygia longifolia (Willd.) Britton y Rose | 21.34 | 8.28 | X | X | X |
| Guadua angustifolia Kunth | 12.35 | 0.86 | X | ||
| Casearia sylvestris Sw. | 5.72 | 0.10 | X | X | X |
| Ficus insipida Willd. | 4.96 | 2.02 | X | ||
| Astronium graveolens Jacq. | 4.34 | 0.82 | X | X | X |
| Pithecellobium lanceolatum (Willd.) Benth. | 3.37 | 1.09 | X | ||
| Myrcia aff. popayanensis Hieron | 2.81 | 0.21 | X | X | |
| Machaerium capote Dugand | 2.77 | 0.19 | X | X | |
| Petrea pubescens Turcz. | 2.36 | 0.77 | X | ||
| Lacistema aggregatum (P.J.Bergius) Rusby | 2.19 | 0.04 | X | ||
| Guazuma ulmifolia Lam. | 2.18 | 0.23 | X | X | |
| Hirtella americana L. | 2.01 | 0.19 | X | ||
| Total especies con mayor IVI | 66.39 | 14.18 | |||
| Total todas las especies | 16.06 |
Se establecieron diez clases diamétricas y diez clases de altura para la comunidad vegetal del bosque ripario. La estructura diamétrica presentó una forma de J invertida, siendo la Clase 1 la que concentró el mayor número de tallos, con 150 individuos (75.38 %) (Figura 2). Para la Clase 2, el número de individuos se redujo significativamente a 24 (12.06 %), y la Clase 9 no presentó individuos. La distribución por clases de altura tuvo tendencias poco claras, con aumentos y caídas aleatorios en el número de tallos y los valores más altos en las Clases 2 y 7 (32 individuos en ambos casos, que representan el 16.08 %) (Figura 3). Sin embargo, para la Clase 7, la mayoría de los tallos (25) corresponden a G. angustifolia. Debido a esto, se decidió no tener en cuenta los individuos de esta especie en el cálculo de las clases de altura, quedando un total de 140 individuos y dando como resultado un máximo de tallos de 32 para la Clase 2 (22.86 %), que luego presentó una disminución gradual en el número de individuos para cada una de las clases siguientes. Las tendencias descritas se pueden observar en la Figura 3.
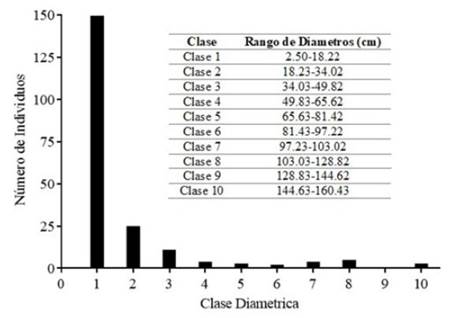
Figura 2 Distribución en clases diamétricas de la vegetación con DAP ≥ 2.5 cm, encontrada en un BST ripario asociado al arroyo La Avería en la cuenca alta del río Magdalena, municipio de Paicol, Huila, Colombia
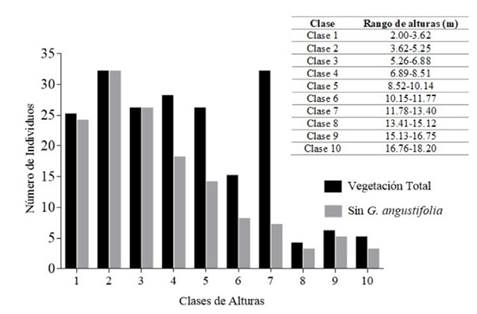
Figura 3 Distribución en clases de alturas de la vegetación con DAP ≥ 2.5 cm encontrada en un BST ripario asociado al arroyo La Avería en la cuenca alta del río Magdalena, municipio de Paicol, Huila, Colombia
En cuanto a los valores obtenidos para el índice QBR, el CDF y la entrada de radiación solar indican que las estaciones de muestreo difieren en el estado de calidad del bosque ripario y la cobertura de dosel (Tabla 2). Se observa que la Estación 1, ubicada en el tramo alto del arroyo, corresponde a una zona con poca perturbación antrópica, por lo que se deduce que la estación proyecta las condiciones naturales del ecosistema ripario y lótico. Por su parte, la Estación 2 presenta la mayor perturbación, causada por la casi desaparición del sistema ribereño como consecuencia de los impactos antrópicos, lo que a su vez reduce el CDF y el área cubierta por el dosel. Es por esto que la Estación 2 registra los valores más bajos del área de estudio. Asimismo, la Estación 3 presentó valores medios en cuanto al estado de la ribera, debido a que se observa únicamente el impacto ganadero, que ha permitido en parte la recuperación del bosque ripario.
Tabla 2 Registro del índice de calidad de bosque ripario (QBR), coeficiente de densidad forestal (CDF) y entrada de radiación solar para las tres estaciones de muestreo estudiadas en un BST ripario asociado al arroyo La Avería en la cuenca alta del río Magdalena, municipio de Paicol, Huila, Colombia
| Estación de muestreo | Valor Cuantitativo Índice QBR | Valor Cualitativo Índice QBR | Coeficiente de Densidad Forestal (%) | Entrada promedio de radiación solar (Watts .m-2) |
|---|---|---|---|---|
| Estación 1 | 95 | Sistema ripario en condiciones naturales | 91.80 | 0.20 |
| Estación 2 | 45 | Fuertes alteraciones, calidad baja del sistema ripario | 57.70 | 0.69 |
| Estación 3 | 68 | Alteraciones importantes, calidad media del sistema ripario | 83.87 | 0.35 |
DISCUSIÓN
La composición del bosque ripario estudiado es similar a la típicamente reportada para los BST y bosques riparios de Colombia. La mayor riqueza de Fabaceae frente a otras familias botánicas es una condición observada en la mayoría de los BST del neotrópico (Gentry, 1995; Álvarez et al., 1998; Pizano et al., 2014; Londoño-Lemos et al., 2022). También se ha reportado a las especies de Fabaceae como elementos frecuentes en los bosques riparios de las tierras bajas de Colombia (Correa-Gómez & Stevenson, 2010; Luna, 2019). De igual manera, la alta riqueza de Malvaceae y Euphorbiaceae es frecuente en este tipo de bosques, y en Colombia se ha estimado que ambas familias representan hasta el 7.6 % del total de especies encontradas en los BST (Rodríguez et al., 2012; Pizano & García, 2014).
La riqueza de la familia Salicaceae estuvo representada por tres especies del género Casearía, que tuvo el mayor número de especies de todo el estudio. La frecuencia de este género dentro de la composición de los BST ha sido informada en otras localidades del país, además de ser un elemento frecuente en áreas que han presentado disturbios (Mendoza-C, 1999; Carrillo-Fajardo et al., 2007; Gutiérrez, 2019; Ruíz & Saab, 2020). Un dato atípico encontrado fue una mayor riqueza de Urticaceae, que no se había reportado como un elemento común dentro de la composición de los BST (Gentry, 1995). A pesar de esto, la mayoría de las especies de esta familia prefiere ambientes húmedos, por lo que son comunes cerca a cursos de agua y se han registrado como elementos frecuentes en la composición de los bosques riparios del neotrópico (Friis, 1993; Wittmann et al., 2008).
La riqueza de 47 especies está dentro del rango reportado para otros BST en el neotrópico y en Colombia. Las comparaciones se centran en datos obtenidos de bosques secos de Centroamérica y Colombia porque existen afinidades florísticas con la zona de estudio (Dryflor, 2016). En Costa Rica y Nicaragua, en áreas de 0.1 ha, se reportaron cifras de riqueza de 44 y 75 especies respectivamente (Gentry, 1995; Gillespie et al., 2000). Para Colombia, la variación va de 14 a 121 especies en la misma área, y de 14 a 71 especies en la cuenca alta del río Magdalena (Gentry, 1995; Mendoza, 1999; Carrillo-Fajardo et al., 2007; IAvH, 2019).
En cuanto a la abundancia, se puede decir que el número de individuos reportados fue bajo (199). Tomando como referencia el estudio realizado por Mendoza (1999) en siete parcelas de BST en Colombia, tres de ellas en la cuenca alta del río Magdalena (DAP ≥ 2.5 cm en 0.1 ha), se observa una variación considerable en el número de individuos. Mendoza (1999) reportó entre 339 y 597, cifras que están por encima de las registradas en este trabajo. Esto podría deberse a las características propias de los bosques riparios o el efecto de la intervención antrópica.
Algunos análisis en bosques riparios han mostrado que estos tienden a tener una densidad menor en comparación con bosques del interior (Suzuki et al., 2002; Damasceno-Junior et al., 2005; Adel et al., 2018). Si bien no existe evidencia directa de este fenómeno en BST, Gentry (1995) muestreó bosques de tierras altas y de galería en Guanacaste (Costa Rica), encontrando que los bosques riparios presentaban un menor número de individuos a pesar de tener una mayor riqueza que los bosques de tierras altas. Una baja cantidad de individuos también fue reportada en llanuras de inundación en BST de Cundinamarca (López et al., 2019), indicando que los bosques riparios del BST podrían presentar naturalmente una baja densidad en comparación con otros tipos de bosques.
La cercanía a entornos urbanos es otro factor que puede explicar el bajo número de individuos, tal como reportaron Gillespie et al. (2000) en dos de nueve parcelas estudiadas en Costa Rica y Nicaragua, que presentaron la menor abundancia y a su vez eran las que más cerca se encontraban de los centros urbanos. Esto respaldaría las diferencias observadas entre las Estaciones 3 y 2 con respecto a la Estación 1, pues esta última registró más del doble de individuos que las dos primeras y es la que más alejada se encuentra del centro urbano. No obstante, se debe tener en cuenta que la mayor parte de los individuos encontrados en la Estación 1 corresponden a G. angustifolia. Ambas explicaciones no son necesariamente excluyentes, por lo que la baja densidad de individuos podría deberse a un efecto combinado de los dos factores. En estudios futuros se recomienda profundizar más en estos aspectos para establecer posibles relaciones de causalidad de manera más certera, analizando los cambios en el número de individuos en relación con diferentes coberturas vegetales.
La estructura del área de estudio mostró elementos típicos del bosque ripario en Colombia y deja en evidencia la degradación que este ha sufrido debido a las actividades humanas. La importancia de Z. longifolia dentro de la comunidad vegetal mostrada por el IVI es un resultado esperado, pues esta especie tiene una amplia distribución a lo largo del país y es un elemento habitual de los bosques riparios, al igual que las demás especies del género Zygia (Romero-Hernández, 2017). Esto se debe a que sus semillas son transportadas por el agua, al rápido crecimiento que presentan y a la capacidad de sobrevivir a las crecientes. Además, se considera una especie benéfica para el mantenimiento y conservación de fuentes hídricas en el BST, ya que sus raíces evitan la erosión de los márgenes (Cerdán et al., 2012; Vargas, 2015).
La especie G. angustifolia presentó el segundo valor más alto de IVI. Sin embargo, esta especie solamente se registró en la Estación 1, donde fue un elemento abundante, lo que indica estados sucesionales tardíos para la zona de muestreo. G. angustifolia suele ser frecuente en los márgenes de los ríos, pero tiende a desaparecer en zonas con presencia antrópica debido a su explotación para la obtención de madera (Vargas, 2015). Otras especies con valores altos de IVI, como C. sylvestris, A. graveolens, P. lanceolatum, Guazuma ulmifolia Lam. y Machaerium capote Dugand, son elementos ampliamente distribuidos en el país y han sido reportados en BST de la cuenca alta del río Magdalena (Mendoza, 1999; Pizano et al., 2014; Vargas, 2015; López et al., 2019; Romero-Duque et al., 2019; Ruíz & Saab, 2020).
La distribución diamétrica en forma de jota invertida, con ausencia de individuos en algunas de las clases y una disminución abrupta en el número de individuos entre categorías diamétricas, es un comportamiento típico de bosques intervenidos en proceso de regeneración (Louman et al., 2001; López et al., 2019). Esto es respaldado por los datos de altura, pues son consistentes con los encontrados en BST del valle del río Cauca, donde los bosques en etapas de sucesión temprana muestran patrones en donde la mayoría de los individuos se concentran en segundas categorías de altura (sin los datos de G. angustifolia) y tienen una altura máxima no superior a los 18 m (Adarve et al., 2010; Londoño y Torres, 2015). La escasa presencia de elementos de sucesión intermedia típicos de BST, tales como Senna spectabilis (DC.) H.S.Irwin y Barneby, G. ulmifolia o Piper amalago L. y un área basal total por debajo del rango reportado para estos bosques (17-40 m2.ha-1), también son indicativos del estadio de sucesión temprano del bosque estudiado (Murphy & Lugo, 1986; Vargas, 2015).
En la Estación 1 se identificaron bajas perturbaciones antrópicas y estadios sucesionales tardíos, corroborados por bajos valores de diversidad en comparación con las Estaciones 2 y 3, junto con la presencia de especies como G. angustifolia (Olascuaga-Vargas et al., 2016). Además, los valores altos del índice QBR soportan esta conclusión, pues se atribuyen a la presencia de una cobertura riparia continua, compuesta en gran medida por vegetación nativa como Z. longifolia, lo cual resulta en un bosque que limita la entrada de radiación solar (Vargas, 2015). Esta información se complementa con los valores altos del CDF y la baja entrada de luz, que son características de los BST maduros (Denward & Tranvik, 1998; Olascuaga-Vargas et al., 2016; Melo et al., 2017).
Por el contrario, la Estación 2 exhibió un valor bajo en el índice QBR, lo cual es propio de ambientes muy perturbados donde la cobertura vegetal riparia ha sido eliminada casi en su totalidad (Suárez et al., 2002). Sumado a esto, los valores de diversidad mayores pueden explicarse como el efecto de etapas sucesionales tempranas en las que existen varios mecanismos que favorecen la coexistencia de especies (Roxburgh et al., 2004; Olascuaga-Vargas et al., 2016). Estos resultados se asocian con la cercanía de la estación al centro urbano del municipio de Paicol, a la descarga de vertimientos municipales en el lugar y a la presencia de cultivos de arroz (Díaz, 2016; Peralta-Muñoz, 2016), actividades que son capaces de modificar el flujo natural del arroyo y alterar la estructura de los bosques riparios (Fernández-Mendez et al., 2013; Sabaj et al., 2014). Estas perturbaciones permiten una mayor entrada de luz, generando cambios en las dinámicas de los BST que pueden retrasar los procesos sucesionales (Melo et al., 2017)
Los valores medios del índice QBR registrados en la Estación 3 son típicos de ambientes con un bajo nivel de perturbación o que se encuentran en proceso de restauración, donde existe una discontinuidad de los parches de árboles en el ambiente ripario (Suárez et al., 2002). En este caso se detectó la presencia de actividades ganaderas, cuya reducción en la diversidad vegetal de los BST se ha demostrado (Ballesteros-Correa et al., 2019). Sin embargo, paulatinamente se ha ido reduciendo la presión ganadera sobre esta zona del bosque, lo que ha permitido el inicio de los procesos sucesionales.
Finalmente, es importante señalar el valor que tienen los bosques riparios en los procesos de restauración de los BST, ya que conservan especies importantes para las diferentes etapas sucesionales de estos bosques (Vargas, 2015). A pesar de esto, las diferentes perturbaciones antrópicas identificadas, sumadas a la falta de información que hay sobre las áreas riparias, ponen en riesgo la conservación de estos bosques. Se recomienda que los trabajos futuros amplíen la información sobre la diversidad y el estado de conservación de los bosques riparios en los BST del país, centrándose en aspectos como el bajo número de individuos o la composición de estos bosques en diferentes estadios sucesionales.
CONCLUSIONES
El bosque ripario estudiado presenta una diversidad florística semejante a la de otros BST y bosques riparios de Colombia y el norte de Sudamérica, como lo muestra la abundancia de la familia Fabaceae y la presencia de especies como Z. longifolia, G. angustifolia, A. graveolens, entre otras. La baja cantidad de individuos reportados en el área de estudio puede ser una característica propia de los bosques riparios del BST o un efecto de las perturbaciones antrópicas que se presentan en dos de las estaciones de muestreo. Los análisis estructurales muestran que, si bien el bosque ripario retiene ciertas características propias de los BST, este tiene condiciones típicas de bosques en etapas sucesionales tempranas. Los valores de diversidad, sumados a los índices QBR y CDF y la entrada de radiación solar indican que la Estación 1 presenta la mejor calidad, con un dosel más cerrado que limita la entrada de luz y muestra un estado sucesional avanzado, como lo indica la presencia de G. angustifolia. Los valores más bajos de diversidad y la menor calidad de las Estaciones 2 y 3 se asociaron con la presencia de actividades agropecuarias y la cercanía al centro urbano del municipio de Paicol, manteniendo estas zonas del bosque ripario en estados sucesionales tempranos.

