











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los matorrales xerófilos son las comunidades vegetales mejor adaptadas a los ambientes áridos o semiáridos (Reyna-González et al., 2021; Rzedowski, 2006). Este tipo de vegetación ofrece distintos servicios ecosistémicos como el suministro de producto maderables y no maderables, la captura de carbono, la dotación de hábitat para fauna silvestre, la infiltración de agua y la belleza paisajística (Alanís-Rodríguez et al., 2021; Molina-Guerra, 2014; Patiño Flores et al., 2019). A pesar de su importante función, el matorral ha sido uno de los tipos de vegetación más afectados en las zonas secas de México (Tarango-Arámbula, 2005). El aprovechamiento desmedido de las especies leñosas y el cambio de uso de suelo son algunas de las principales causas que han propiciado la disminución de la cobertura original de los matorrales (Arriaga, 2009). Tal es el caso del matorral espinoso tamaulipeco, cuya extensión se ha visto afectada debido al cambio de uso de suelo para fines principalmente agropecuarios (R. Foroughbakhch et al., 2009). El posterior abandono de actividades que motivan estos cambios abre la oportunidad para la implementación de programas de restauración, como una alternativa para recuperar la cantidad y calidad de este y otros ecosistemas terrestres (López-Barrera et al., 2017).
En los últimos años, la ecología de la restauración ha demostrado los beneficios que conlleva su aplicación (Howe & Martínez-Garza, 2014). Ya sea que se aplique un sistema con ayuda del ser humano en el que se planten especies para recuperar la vegetación con mayor rapidez (restauración activa) o, por el contrario, se deje que la misma naturaleza se recupere por sí sola, aunque tome más tiempo (restauración pasiva), estos procesos permiten mejorar las condiciones de los ecosistemas deforestados (Morrison & Lindell, 2010). Sin embargo, para poder llevar a cabo una restauración exitosa de una comunidad vegetal degradada, es necesario contar con información adecuada sobre la estructura y composición florística, así como de la dinámica, mecanismos de regeneración y patrones de crecimiento de las especies originales (Alanís-Rodríguez et al., 2021; Alanís-Rodríguez et al., 2016; Pequeño-Ledezma et al., 2016).
En diversas partes del mundo se han desarrollado estudios sobre el crecimiento de árboles orientados a la restauración ecológica, principalmente en especies tropicales y templadas (Dias & Marenco, 2016; Ram et al., 2011; Villar et al., 2014). En México, la mayor parte de los trabajos sobre crecimiento de árboles y arbustos y/o restauración también se han realizado en selvas o bosques templados (Ceccon et al., 2015; Hernández-Valera et al., 2018) y en matorrales en menor proporción (R. Foroughbakhch, 1992).
A pesar de los avances en el conocimiento sobre ecología de la restauración y la regeneración que se han registrado en las zonas de matorrales del noreste de México (Alanís-Rodríguez et al., 2016; Foroughbakhch et al., 2001; Foroughbakhch & Hernández-Piñero, 2014; Jiménez-Pérez et al., 2013; Martínez-Hernández et al., 2013), aún existen vacíos en el entendimiento de los procesos, mecanismos y factores que intervienen en la dinámica de la vegetación y la regeneración de las comunidades vegetales de zonas áridas y semiáridas del país. Por esta razón, el objetivo de este trabajo fue evaluar la composición de especies vegetales de un matorral espinoso tamaulipeco restaurado, así como la respuesta de seis especies leñosas nativas empleadas para este fin y dos establecidas de forma natural, a través de los atributos de la comunidad y las variables dasométricas de las especies utilizadas (cobertura de copa, altura, diámetro normal y volumen).
MATERIAL Y MÉTODOS
Área de estudio
El estudio se desarrolló en la región noreste de México, en el Área de Conservación del Complejo Industrial Ternium Pesquería en el municipio de Pesquería, Nuevo León (Figura 1). El sitio está en las coordenadas 25°45’17.78” de latitud norte y 99° 58’01.40” de longitud oeste, sobre un suelo predominantemente denominado vertisol (INEGI, 2021). Según la clasificación de Köppen, modificada por García (2004), el clima predominante es muy seco semicálido (BWhw), con una temperatura media anual entre los 20 y 21 °C. La precipitación media anual es de 550 mm. El sitio se ubica a una altitud de 330 msnm, a una distancia de 400 m del río Pesquería, en terrenos planos o con ligera pendiente, los cuales forman parte de la Planicie Costera Nororiental (CONABIO, 2018; Patiño Flores et al., 2019). La comunidad vegetal representativa es el matorral espinoso tamaulipeco, con vegetación secundaria arbustiva y con vecindad a zonas agrícolas y bosque de galería (Patiño-Flores et al., 2019).
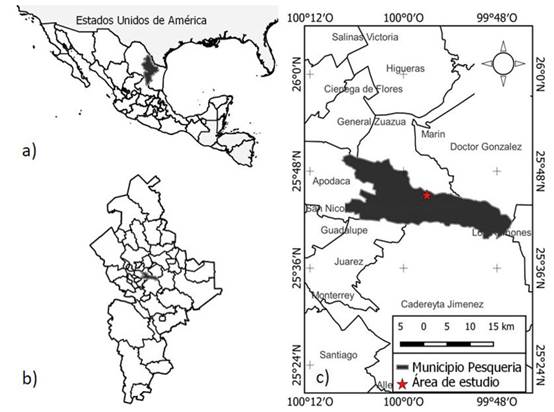
Figura 1 Localización del área de estudio: A) México resaltando el estado de Nuevo León, B) Nuevo León resaltando el municipio de Pesquería y C) Pesquería resaltando el área de estudio
De acuerdo con su historial de uso, el área donde se desarrolló la reforestación tenía una vegetación perteneciente a un matorral espinoso tamaulipeco, pero durante dos décadas (1989-2009) se le dio un uso agrícola, mediante el cual se cultivó avena, trigo y maíz. Los cultivos se sembraron de manera rotativa y fueron regados mediante un canal de agua a gravedad. Cuatro años después del abandono del cultivo, se eliminó la vegetación (herbáceas y especies que anteriormente habían sido cultivadas) utilizando un tractor con chapoleadora. La restauración se realizó en abril de 2013, con seis especies arbóreas y arbustivas, cuyas plantas fueron adquiridas en viveros forestales de la localidad (Tabla 1). Las semillas utilizadas en la producción de plantas se obtuvieron en comunidades vegetales maduras de la localidad. Las plántulas tenían un año de edad, estaban lignificadas y tuvieron una etapa de 1.5 meses de endurecimiento.
Tabla 1 Especies consideradas en la evaluación (nombre científico, familia, forma de vida, abreviatura y nombre común).
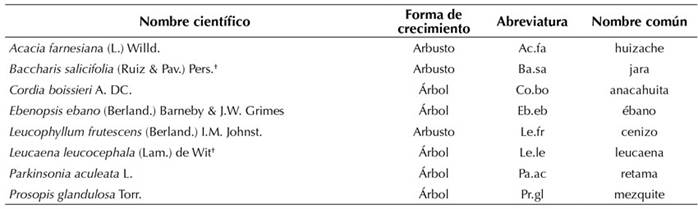
Las especies marcadas con el símbolo (†) son aquellas establecidas en el sitio de manera natural.
La selección de especies se realizó de acuerdo con su alta presencia en las primeras fases de la sucesión natural del MET (Alanís-Rodríguez et al., 2008; Jiménez-Pérez et al., 2012; Jiménez-Pérez et al., 2013). Los individuos seleccionados presentaron buen vigor y una altura promedio de 50 ±10 cm (valor promedio ± error estándar), así como un diámetro basal promedio de 1.0 ±0.2 cm. La restauración se realizó en una superficie de 3.3 ha, con un diseño a tresbolillo con una distancia 1.5 m entre individuos y líneas y una densidad total de plantación de 992 individuos por hectárea (N.ha-1). Con el uso de maquinaria agrícola se generaron cepas de 1 m3 para ablandar el suelo compactado. Después, en cada cajete se aplicó hidrogel (20 g a cada planta) y enraizador (se diluyó 2 g por cada litro de agua, y se aplicó a razón de 20 litros por planta), y se colocó cada planta dejando un cajete de 5 cm de profundidad para favorecer la captación de agua (CONAFOR, 2010). A cada plántula se le colocó un tutor para asegurar el crecimiento recto del individuo y evitar que el tallo se rompiera o quebrara por la acción del viento, así como una etiqueta para su seguimiento. Se realizaron riegos de auxilio diarios de 10 L durante los primeros dos meses y, del tercer al doceavo mes, se regó una vez por semana. Para evitar la competencia por luz y nutrimentos, durante los primeros 18 meses se realizó la eliminación física de las herbáceas en el área mediante una desbrozadora (Alanís-Rodríguez et al., 2016).
Análisis de la vegetación
En el invierno de 2018 (seis años después de realizada la plantación) se establecieron 12 sitios de muestreo de 10 × 10 m (100 m2) distribuidos aleatoriamente. En cada sitio se registraron todos los individuos con un diámetro basal ≥ 2 cm (d 0.10 ≥ 2 cm). Para la identificación taxonómica de las especies regeneradas, se utilizó la literatura de Molina Guerra et al. (2019), registrando para cada individuo la altura total (h), diámetro basal medido a 10 cm de la base de la planta (d 0.10) y dos diámetros de copa (k): en dirección norte-sur (k n-s) y este-oeste (k e-o) (Alanís-Rodríguez et al., 2020).
Para estimar el diámetro promedio de cada individuo, se utilizó la fórmula k- = (k n-s + k e-o) / 2, donde k- es el diámetro promedio de la copa, k n-s el diámetro medido en dirección norte-sur y k e-o el diámetro medido en dirección este-oeste. La cobertura de la copa se calculó mediante la fórmula A = π/4 × d 2, donde A es el área de copa, π un valor constante de 3.14 y d el diámetro promedio de la copa. El volumen (V) de cada individuo se estimó mediante la fórmula V = g × h × CM, donde g es el área basal, h la altura total de individuo y CM el coeficiente mórfico, que, para el caso del matorral espinoso tamaulipeco, es 0.5 (Alanís-Rodríguez et al., 2020).
Se realizaron pruebas para verificar que se cumplieran los supuestos de distribución normal mediante la prueba de Shapiro-Wilk y la de homogeneidad de varianzas mediante la prueba de Levene, ambas con un nivel de significancia p<0.05. Una vez validado que se cumplieran los supuestos anteriormente descritos, se realizó un análisis de varianza (ANOVA) de un factor (especie) para los valores de cobertura (m2), diámetro basal o diámetro de la base (cm), altura total (m) y volumen (m3). Para el análisis post-hoc, se utilizó la prueba de Duncan, en aras de determinar si existían diferencias significativas (p≤0.05) entre especies con respecto a las variables analizadas. Dichos datos fueron analizados en el software para análisis estadístico R-project (R Core Team, 2019) y en RStudio v1.0.136 (RStudio Team, 2020), utilizando stats v3.3.3.
RESULTADOS
Composición
Además de las seis especies inicialmente plantadas, se registraron otras dos (Leucaena leucocephala y Baccharis salicifolia) que no se plantaron durante reforestación, pero que geminaron, se establecieron y crecieron de manera natural (Tabla 1).
Cobertura, altura y diámetro normal
Las especies que presentaron estadísticamente mayor cobertura fueron Parkinsonia aculeata (13.88 ±2.70 m2) (valores promedios ± desviación estándar), Acacia farnesiana (12.11 ±1.86 m2) y Cordia boissieri (7.90 ±1.08 m2). La especie arbustiva Leucophyllum frutescens fue la que presentó el menor valor (0.77 ±0.17 m2) (Figura 2a).

Figura 2 Valor promedio ± error estándar: a) cobertura (m2), b) diámetro basal (cm2 ), c) altura total (m) y d) volumen (m3) de las ocho especies evaluadas. Letras diferentes indican diferencias significativas (p≤0.05) entre las especies de plantas, de acuerdo con la prueba de comparación de medias de Duncan. Ac.fa: Acacia farnesiana; Ba.sa: Baccharis salicifolia; Co.bo: Cordia boissieri; Eb.eb: Ebenopsis ebano; Le.fr: Leucophyllum frutescens; Le.le: Leucaena leucocephala; Pa.ac: Parkinsonia aculeata; Pr.gl: Prosopis glandulosa
Las dos especies con mayor diámetro basal fueron Parkinsonia aculeata (13.10 ±1.03 cm) y Acacia farnesiana (10.09 ±1.00 cm), registrando promedios de 2.18 y 1.68 cm por año respectivamente (Figura 2b).
La especie que presentó mayor altura fue Parkinsonia aculeata (4.92 ±0.17 m), con un crecimiento medio de 0.77 m año-1. Las especies que le siguen son Acacia farnesiana (3.54 ±0.18 m), Baccharis salicifolia (3.23 ±0.24 m) y Ebenopsis ebano (2.93 ±0.20 m) (Figura 2c).
Las dos especies con mayor volumen fueron Parkinsonia aculeata (0.0356 ± 0.0062 m3) y Acacia farnesiana (0.0253 ± 0.0041 m3), mientras que la de menor volumen se registró en Leucaena leucocephala (0.0004 ±0.0001 m3) (Figura 2d).
DISCUSIÓN
Composición
La composición florística del área restaurada no solo fue el resultado de las especies plantadas, sino también producto de la regeneración natural. La presencia de las especies no plantadas en la actividad de restauración activa pero registradas en este estudio se debió a la disponibilidad de propágulos en el sitio, como consecuencia de la dispersión desde áreas aledañas, así como del desempeño ecofisiológico característico de estas especies pioneras y oportunistas. La especie nativa B. salicifolia produce gran cantidad de semillas con apéndices que facilitan la dispersión anemócora de larga distancia (Boland, 2017). Su dispersión ocurre entre mayo y junio, meses posteriores al inicio de la intervención realizada en este estudio. Además, sus semillas son fotoblásticas (Bonner & Karrfalt, 2008) y, a pesar de ser recalcitrantes (Ferri et al., 2009), las semillas que componen el banco de semillas transitorios, formado luego de la lluvia de semillas, estuvieron disponibles cuando las condiciones fueron propicias, lo que puede explicar su presencia en la comunidad restaurada. De igual forma, su buen desempeño ecofisiológico, producto de sus adaptaciones a ambientes xéricos que le otorgan protección a la desecación, así como su alta tasa fotosintética (Tosoratto et al., 2016), pueden explicar el establecimiento y sobrevivencia de B. salicifolia, aun sin haber sido favorecida a través de los tratamientos aplicados.
La presencia de la otra especie que no fue incorporada al sitio como parte del procedimiento de restauración activa fue L. leucocephala. En cuanto a la disponibilidad de fuentes de propágulos en el sitio, la dispersión de sus semillas es a cortas distancias, provenientes principalmente de árboles semilleros colindantes a la zona restaurada. Esta especie ha sido reportada en áreas urbanas y bosques de galería del noreste de México (Canizales-Velázquez et al., 2021; Leal-Elizondo et al., 2018; Mata-Balderas et al., 2020). También se ha reportado esta especie con alta densidad de individuos en una comunidad vegetal ribereña regenerada en el área metropolitana de Monterrey, México. Autores como Hata et al. (2010) observaron una correlación negativa entre la presencia de plántulas y la distancia de los árboles padres, lo que sugiere que las semillas de L. leucocephala se dispersan a distancias cortas. En la vecindad del área restaurada en el presente estudio (a 400 m) se ubica una sección del río Pesquería, con vegetación ribereña y presencia de L. leucocephala. Además, su presencia podría ser explicada por la germinación y establecimiento de semillas presentes en el suelo, ya que esta especie presenta semillas ortodoxas que pueden formar bancos de semillas, las cuales están disponibles ante las condiciones favorables para la germinación (Marques et al., 2014). El establecimiento y la supervivencia de los individuos de L. leucocephala puede deberse a su resistencia a las condiciones de sequía y su capacidad de establecerse en suelos pobres. Yige et al. (2012) demostraron la habilidad de L. leucocephala para mantener el contenido de agua en las hojas, así como para incrementar el contenido de clorofila y el uso eficiente del agua, mediante la reducción de la tasa fotosintética y transpiración durante la sequía. De igual forma, su capacidad de prosperar en suelos pobres se debe a su capacidad de fijar nitrógeno (de Angelis et al., 2021).
En este estudio, los individuos de B. salicifolia y L. leucocephala mostraron su capacidad de regeneración natural en el sitio intervenido. Estas dos especies revelaron una germinación, un establecimiento y un desarrollo bajo condiciones de baja cobertura características de las etapas iniciales de la sucesión, con ausencia de dosel desde el inicio de la reforestación. Tanto B. salicifolia como L. leucocephala son especies pioneras demandantes de luz y de rápido crecimiento, características de las etapas iniciales en procesos de sucesión (Ferri et al., 2009; Marod et al., 2012), por lo que las condiciones iniciales de baja cobertura del dosel les fueron favorables. Otros autores (Patiño-Flores et al., 2022a, 2022b) también registraron la presencia de B. salicifolia en las etapas iniciales de la sucesión ecológica en el matorral espinoso tamaulipeco. En un estudio de regeneración de sitios degradados por erosión en la zona semiárida de la mixteca, Lira-Caballero et al. (2018) observaron la presencia de B. salicifolia en las etapas iniciales de la sucesión secundaria. De manera similar Colón y Lugo (2006) observaron la dominancia de L. leucocephala durante los primeros y hasta los 45 años en condiciones de dosel abierto.
Cobertura, altura y diámetro normal
Las especies con mayor cobertura, diámetro normal, altura total y volumen fueron P. aculeata, y A. farnesiana (Figura 2). Este comportamiento puede deberse a la estrategia ecológica de estas especies, que es característica de las especies pioneras de rápido crecimiento. Dichas características pueden ser favorecidas en condiciones de plantación a través de estrategias de riego y eliminación de herbáceas. Algunos estudios (R. Foroughbakhch, 1992; R. Foroughbakhch et al., 2012) observaron, en condiciones de plantación, que estas especies tienen un rápido crecimiento de copas, así como de altura y diámetro basal. Este crecimiento puede verse favorecido cuando el recurso luz no es limitante (R. Foroughbakhch, 1992), y es ahí que las pioneras B. salicifolia y E. ebano también pueden encontrar las condiciones para un buen crecimiento en altura (Figura 2c). A diferencia de P. aculeata y A. farnesiana, la mayor cobertura observada en C. boissieri no correspondió a una tendencia similar en los otros dos parámetros (altura y diámetro normal). Esto puede deberse a que C. boissieri es una especie pionera, con rápido crecimiento al inicio de su establecimiento (R. Foroughbakhch, 1992), pero moderado en los años siguientes (Navar et al., 2014), y con mayor longevidad que las dos primeras. Esto puede representar una habilidad distinta de dominancia del dosel, no por medio de la altura sino de su cobertura.
Tanto las especies reforestadas como las que aparecieron de forma natural son propias de valles y planicies. Si bien es cierto que algunas especies como Cordia boissieri pueden además desarrollarse en los matorrales submontanos de las laderas montañosas, otras como Ebenopsis ebano, Parkinsonia aculeata y Prosopis glandulosa prefieren una orografía plana y con mayor humedad (Molina-Guerra et al., 2019).
La mayor cobertura, diámetro y altura observadas para P. aculeata y A. farnesiana sugiere que el tratamiento aplicado tuvo mejores resultados respecto al resto de especies. Esta situación es en parte reforzada por la similitud en las alturas promedio alcanzadas por estas dos especies en el presente estudio (a los seis años de ser plantadas en el sitio, como resultado de las actividades de restauración) y las registradas en sitios de referencia del MET con vegetación madura (Domínguez-Gómez et al., 2013). La igualdad estadística de la altura de A. farneciana y B. salicifolia denota su naturaleza oportunista. A su vez, la segunda es caracterizada por los atributos ecofisiológicos que explican su dispersión, germinación establecimiento y rápido desarrollo, aun sin los tratamientos y cuidados emprendidos con las otras especies.
CONCLUSIONES
Mediante la evaluación de la composición y de la respuesta de las especies medida a través de variables dasométricas, se logró determinar la importancia de la dispersión y reclutamiento de las especies que no participaron en la revegetación emprendida como parte de las actividades de restauración. Así, en el caso de restauración activa, en la que se plantan especies particulares, el volumen y la altura pueden ser más importantes, ya que pueden dar una mejor idea del desarrollo y la condición de las mismas en el proceso de seguimiento y evaluación de la restauración. Así, la mayor cobertura de copa, diámetro normal y volumen de las pioneras P. aculeata y A. farnesiana se debe a su estrategia ecológica: estas especies son de rápido crecimiento en todo su desarrollo, y se establecen y crecen de mejor manera en sitios abiertos (en este caso, en condiciones de deforestación por agricultura).
Una restauración activa exitosa debe considerar la incorporación de las especies que caractericen a la vegetación de referencia, así como la composición de especies que intervienen en las distintas fases de la dinámica de la vegetación. También debe considerar la vegetación circunvecina como fuente potencial de propágulos. Las dos especies que se regeneraron a partir de semillas disponibles en el sitio de manera natural (banco de semillas y lluvia de semillas de L. leucocephala y B. salicifolia), que fueron dispersadas a partir de fuentes de semilla aledañas, fueron especies pioneras, consideradas también como especies oportunistas, que intervienen en las fases iniciales de la sucesión. Se recomienda el uso de P. aculeata, A. farnesiana y Cordia boissieri en futuros proyectos de restauración en el matorral espinoso tamaulipeco, ya que estas son las especies que presentaron mejores resultados en las variables dasométricas. Finalmente, el presente trabajo muestra que la revegetación asistida puede favorecer el rápido crecimiento de algunas de las especies incorporadas con las actividades de restauración activa.

