










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista ION
versão impressa ISSN 0120-100X
Rev. ion vol.25 no.2 Bucaramanga jul./dez. 2012
UTEX 1803 en fotobiorreactores a escala laboratorio
Andrés Fernando Barajas Solano*; Carlos Augusto Godoy Ruiz; Juan David Monroy Davila; Crisóstomo Barajas Ferreira; Viatcheslav Kafarov.
Escuela de Ingeniería Química. Universidad Industrial de Santander (UIS)
Cra 27 Calle 9, Bucaramanga, Colombia
*andresfdobarajas@gmail.com
Fecha Recepción: 6 de febrero de 2012
Fecha Aceptación: 11 de septiembre de 2012
Las microalgas han sido sugeridas como una respuesta adecuada para la producción de combustibles debido a sus ventajas relacionadas con altas tasas de crecimiento, alta eficiencia fotosintética y alta producción de biomasa comparada con otros cultivos terrestres energéticos.
Se estudió el efecto de la inyección continua de CO2 sobre la productividad de biomasa de Chlorella vulgaris UTEX 1803 a escala de laboratorio. Los cultivos se realizaron en condiciones autotróficas a 23±1°C, con ciclos luz-oscuridad 12:12h durante periodos establecidos de 5 días. Los cultivos se efectuaron a concentraciones de NaNO3 (0,59mM; 1,18mM; 1,76mM; 2,35mM) y flujos continuos con 2,3 y 3,8%v de CO2. Todos los experimentos fueron desarrollados por triplicado. Los resultados obtenidos en los diferentes experimentos muestran que las mejores productividades fueron 0,28(g/l.dia) y 0,44(g/l.dia) utilizando 2,3%v de CO2, 0,59mM NaNO3 y 3,8%v de CO2, 1,18mM NaNO3 respectivamente, indicando que la cantidad de CO2 influencia genera la mayor influencia en el crecimiento de C. vulgaris. Sin embargo, al disminuir la concentración de nitrógeno, es posible obtener una mayor productividad de biomasa, esto representa un avance importante en la producción de cultivos fotosintéticos, ya que abre las posibilidades para mejorar la producción total de biomasa mientras se reduce los costos en la producción.
Palabras clave: microalgas, Chlorella vulgaris, cultivo autotrófico, dióxido de carbono, fotobioreactores.
UTEX 1803 on labscale photobioreactors
Microalgae have been suggested as an appropriate response to the production of fuels due to their advantages associated with high growth rates, high photosynthetic efficiency and high production of biomass compared to other terrestrial energy crops.
This work focuses on the effect of continuous inyection of CO2 on biomass productivity of Chlorella vulgaris UTEX 1803 on labscale photobioreactors. Autotrophic cultures were carried out on 23±1°C, with light:dark cicles of 12:12, also different fluxes of CO2 (2.3 and 3.8 v%) and different concentrations of sodium nitrate (0.59mM; 1.18mM; 1.76mM; 2,35mM) were tested.
Results obtained in the different experiments show that the best productivity were 0.28 (g/l.dia) and 0.44(g/l.dia) using 2.3v% de CO2, 0.59mM NaNO3 and 3.8v% de CO2, 1.18mM NaNO3 respectively, Indicating that the amount of CO2 influence generates the greatest influence on the growth of C. vulgaris. However, by lowering the concentration of nitrogen, it is possible to obtain a higher productivity of biomass. This represents an important advance in photosynthetic crop production, as it opens the possibilities to improve the total biomass production while reducing costs production.
Keywords: microalgae, Chlorella vulgaris, autotrophic cultures, carbon dioxide, photobioreactors.
El calentamiento global es un serio problema ambiental, el cual se atribuye principalmente a los gases de efecto invernadero como el metano, ozono troposférico, clorofluorocarbonados y dióxido de carbono (CO2) [1], este último es considerado el principal Gas de Efecto Invernadero (GEI) [2, 3, 4] y se estima que sus emisiones anuales aumentaron un 80% entre 1970 y 2004, afectando la estabilidad económica, social y ambiental del planeta [5]. Por ello su mitigación y utilización se ha estudiado de manera considerable [6].
Las microalgas representan alrededor del 0,5% de la biomasa global, además presentan ventajas tales como la alta tasa de crecimiento, alta eficiencia fotosintética y alta producción de biomasa comparada con otros cultivos terrestres [7, 8], es por esto que pueden llegar a producir alrededor del 70% del oxígeno neto en la tierra, por lo que se ha considerado 10 a 50 veces más eficientes en fijación de CO2 que las plantas terrestres [9]. Emplear este tipo de cultivos para la biofijación de dióxido de carbono, tiene el potencial, no sólo de reducir los costos en la obtención de productos de valor agregado (biodiesel y otros) sino también la compensación de las emisiones de carbono [6, 9, 10, 11].
Basados en literatura, las microalgas pueden fijar cerca del 9% de la luz solar incidente, la cual pueden aprovechar para generar hasta 280 ton de biomasa seca/ha.año, mientras consume cerca de 513 toneladas de CO2 [3]. Además pueden fijar CO2 del ambiente en lugares con concentraciones 1000 veces más altas que las ambientales, lo cual podría ser útil para controlar los altos niveles de CO2 industriales [12]. El CO2 fijado es utilizado en la síntesis de diferentes biomoleculas de valor agregado como proteínas, carbohidratos, pigmentos y lípidos, los cuales a su vez pueden ser aprovechados para la obtención de diferentes productos tales como alimentos, combustibles y otros [6, 9]. La fijación del CO2 por medio de microalgas es ecológicamente sostenible cuando se combina con otros procesos de protección ambiental tales como el tratamiento de aguas residuales [13] o la remoción de metales pesados [14]. Sin embargo la tasa de inyección y la transferencia eficiente del CO2 al medio de cultivo junto con la intensidad de luz son los mayores factores en la asimilación fotosintética del carbono en plantas y en otros organismos fotosintéticos [15-18]. Según Pulz O [18], el balance entre CO2/O2 es también un factor primordial en el logro de una mayor tasa fotosintética. Así por ejemplo, un exceso de oxígeno (CO2/O2) reducirá significativamente las velocidades de crecimiento celular, sin embargo la relación CO2/ O2 que evita la foto-oxidación varia de organismo en organismo [19], por lo cual los cultivos fototróficos pueden no alcanzar siempre altas tasas de crecimiento y productividad [20].
La cantidad de luz requerida para la fotosíntesis es aproximadamente de un quinto a una decima parte de la cantidad de luz disponible al medio día [19]. La tasa de crecimiento aumenta, de la misma manera que aumenta la irradiación de luz hasta una irradiación máxima, en la cual el crecimiento es inhibido, generando un fenómeno conocido como fotoinhibición [21-25], esta es una de las mayores causas en la reducción de la productividad del alga [26]. A una mayor densidad celular en el cultivo, menor será la cantidad de luz que puede penetrar al mismo sin embargo este aumento en la densidad celular propone otros riesgos para el cultivo, ya que produce efectos de sombra entre las células, el cual decrece la intensidad, lumínica exponencialmente desde la superficie del cultivo hasta el centro de éste, generando así una gran zona oscura en el centro del cultivo y una pequeña zona iluminada cerca de la superficie donde la fotosíntesis puede ocurrir [27-29].
La fuente de carbono es el mayor contribuyente en la producción de biomasa para la microalga. El carbono puede ser agregado en compuestos orgánicos como la peptona o ácido acético, o en compuestos inorgánicos como el dióxido de carbono [19]. Durante la fotosíntesis, las microalgas pueden fijar el dióxido de carbono de diferentes fuentes, incluida la atmósfera, gases industriales de chimenea y soluciones salinas de carbonato (NaHCO3 y Na2CO3) [8].
La fijación del CO2 por parte de las microalgas implica un crecimiento fotoautotrófico de las células; la capacidad de fijación del CO2 para algunas especies, puede estar relacionada con el crecimiento de las células y el adecuado aprovechamiento de la luz [15, 30]. Además, la eficiencia de la fotosíntesis en microalgas decrece con un incremento de la temperatura, ya que la solubilidad del CO2 decrece de manera significativa [18].
La disolución del dióxido de carbono en el agua es compleja debido a las reacciones reversibles en fase líquida, las cuales generan 4 especies diferentes en el medio (dióxido de carbono, CO2, ácido carbónico, H2CO3, ion bicarbonato, HCO3-, e ion carbonato, CO3=) cuyas concentraciones en equilibrio dependen del pH [31]. Además, el crecimiento celular depende de la velocidad de asimilación del CO2, su velocidad de flujo, el tamaño de burbuja y el tiempo de residencia de la burbuja [31, 32]. En general, obtener una amplia área de interface gas/líquido es el primer paso vital para aumentar la transferencia de masa del CO2 [32, 17]. Sólo una baja fracción del dióxido de carbono inyectado se incorpora a la biomasa de las microalgas. Utilizando únicamente el CO2 presente en el aire (0,036%) se obtiene una baja productividad. Por lo tanto la solución a este déficit es mezclar CO2 con aire en bajas proporciones del total del flujo de gas para obtener un buen crecimiento [21, 29, 33, 34, 35].
Dentro de la gran diversidad de microalgas, se ha encontrado que C. vulgaris es un organismo altamente eficiente en la fijación del CO2 debido a su habilidad de crecer en medios enriquecidos con altas concentraciones de este gas [36, 37]. lo cual le confiere una gran ventaja sobre otras especies del mismo género [38, 39]. Según estudios realizados por Jeong ML et al. [6] y Lv JM et al. [40], la fijación de CO2 por C. vulgaris requiere de ciertos parámetros como pH 5,7, además de un balance entre las diferentes especies de carbono. Para este caso, se encontró que el medio cultivo contenía 4,61x10-3M de CO2 acuoso (H2CO3) y 8,92x10-4M del ion bicarbonato (HCO3-), lo cual indica para la fijación del CO2, que C. vulgaris puede utilizar estas dos especies químicas. Sin embargo, para lograr un secuestro eficiente es necesario tener en cuenta ciertos factores abióticos del medio de cultivo, dentro de estos el más influyente es el nitrógeno [41, 42, 43, 44, 45], el cual mediante diferentes rutas metabólicas regula significativamente la producción y composición de la biomasa microalgal.
El estrés generado por déficit de nitrógeno, en comparación con el recurso de carbono, puede afectar la respuesta de las células a la disponibilidad de nutrientes y por tanto afectar las tasas de crecimiento y el consumo de la fuente de carbono, [41]. Para el caso del CO2, su metabolismo está fuertemente relacionado con la asimilación del nitrógeno (tanto de NO3 - como de NH4+) debido al costo energético celular [46, 47], ya que hasta el 55% del carbono asimilado está comprometido para la asimilación y metabolismo del nitrógeno [45], por lo cual es posible mejorar la tasas de fijación de CO2 mediante una regulación en el contenido de carbono y nitrógeno presente en el medio. Sin embargo, aun conociendo la existencia de esta dependencia metabólica, no es posible generalizar la relación carbono/ nitrógeno en microalgas, dado que existe un potencial para perturbar los organismos mediante la modificación de otros factores abióticos como: intensidad lumínica, pH y otros [48]. Además existen diferencias entre las especies y grupos de microalgas, tanto en la rapidez y magnitud de las respuestas como en el umbral que hace significativas estas respuestas [41].
Para el caso de Chlorella vulgaris la regulación en la concentración inicial de nitrógeno se ha enfocado principalmente en el aumento en el contenido lipídico [49, 50, 51]. Sin embargo, no existen estudios en los cuales se evalué el efecto de la relación entre la fuente de carbono (ya sea orgánico o inorgánico) y de nitrógeno, por lo cual el objetivo de este trabajo fue mejorar el secuestro de CO2 por Chlorella vulgaris UTEX 1803 en fotobiorreactores a escala laboratorio mediante un ajuste en la relación Carbono/Nitrógeno.
Métodos de cultivo
La microalga Chlorella vulgaris UTEX 1803, fue adquirida de la colección de cepas proveniente de la Universidad de Texas (Austin, Texas, USA); se cultivó en medio Bold Basal, cuya composición en g/l es: NaNO3 (2,94 X 10-3), MgSO4.7H2O (3,04 X 10-4) NaCl (4,28 X 10-4), K2HPO4 (4,31 X 10-4), KH2PO4 (1,29 X 10-3), CaCl2.2H2O (1,70 X 10-4) y micronutrientes (g/l) ZnSO4.7H2O (3,07 X 10- 5), MnCl2.4H2O (7,28 X 10-6), MoO3 (4,93 X 10-6), CuSO4.5H2O (6,29 X 10-6), Co(NO3)2.6H2O (1,68 X 10-6), H3BO3 (1,85 X 10-4), EDTA (1,71 X 10-4), KOH (5,53 X 10-4), FeSO4.7H2O (1,79 X 10-5). Como reactores se usaron botellas plásticas transparentes de 2,5l con diámetro interno de 14cm y 35cm altura con un volumen del cultivo de 2l. Los reactores se acoplaron a un sistema de aireación por burbujeo para la inyección de aire y CO2. Con este sistema se buscó mejorar la homogeneidad de todo el cultivo.
Relación carbono/nitrógeno
Con el fin de mejorar la productividad de biomasa, se probaron diferentes relaciones carbono/nitrógeno en condiciones autotróficas. Se utilizaron flujos continuos de CO2 y diferentes concentraciones de nitrato de sodio (Tabla 1). La inyección continua de CO2 fue proporcionada por una bala de CO2.
Cada uno de los tratamiento se realizó por triplicado sometidos a ciclos de 12:12 horas de luz: oscuridad y a una temperatura de 23±1°C; todos los experimentos tuvieron un tiempo de cultivo de 5 días.
Cuantificación de biomasa
Para las mediciones de concentración de biomasa, se tomó una muestra de 6 ml de cada cultivo. Posteriormente se diluyó a una relación muestra:agua destilada de 1:8. Luego se utilizó un espectrofotómetro (Spectroquant® Pharo 300) con el cual se midió la densidad óptica de cada muestra a una longitud de onda de 500nm (los valores típicos de medición están entre 0,1 - 0,9). La densidad óptica (DO) fue correlacionada con la cantidad de biomasa (g/l, CB) de acuerdo con:

Concentración de clorofila total
Para la cuantificación de clorofila total, se tomaron alícuotas de 10ml de cada fotobioreactor, la cual fue centrifugada a 3400rpm durante 7min (Centrifuga PowerSpin™ MX). Luego se extrajo el sobrenadante con el fin de eliminar el medio presente. El pellet (biomasa resultante) fue resuspendido en 3ml de etanol al 99%v. La mezcla se llevó a calentamiento en baño maría durante 5min. Luego de un posterior enfriamiento se agregaron 2ml adicionales de etanol y se centrifugó por 1min a 2000rpm. La concentración de clorofila en el sobrenadante se calculó leyendo la absorbancia (A) del pigmento en el espectrofotómetro a 650 y 665nm. La relación de la cantidad de clorofila (a+b) en el sobrenadante Ca+b (mg/l) con la densidad óptica fue correlacionada de acuerdo con la ecuación de Becker [52].

Análisis estadístico
Se estudió la influencia del la relación C/N en la productividad de biomasa y cantidad de clorofila (a+b) de C. vulgaris en cultivos autotróficos, mediante el análisis de 2 diseños factoriales de 2 factores utilizando el software Demo-MINITAB. Los factores a estudiar fueron: %CO2 con 2 niveles (2,3 y 3,8%v) y concentración de NaNO3 con 4 niveles (0,59mM; 1,18mM; 1,77mM; 2,35mM).
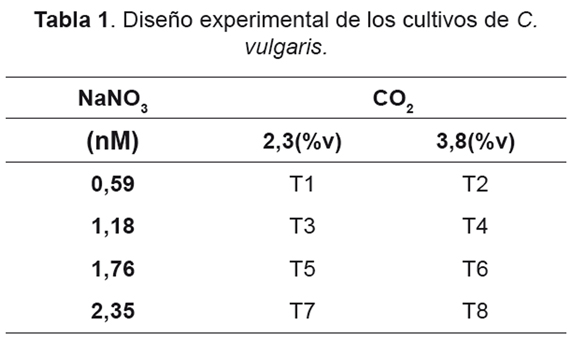
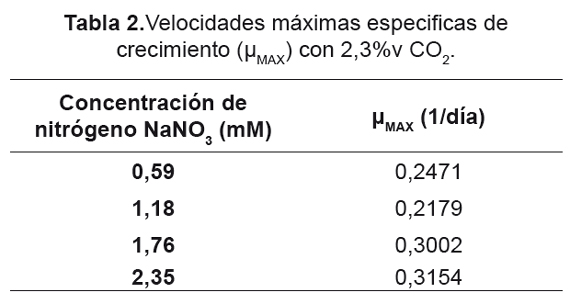
Para cada uno de los diferentes tratamientos, no se encontró inhibición aparente tanto por las concentraciones de CO2 utilizadas como las de NaNO3 (Figuras 1 y 2), Para el caso de 2,3%v de CO2 (Figura 1), no se presentaron etapas de adaptación, lo cual permitó un rápido crecimiento desde el primer día de cultivo, estos resultados son consistentes con los obtenidos por Fan et al. [53], donde un cambio en las condiciones del cultivo microalgal acarreará cambios en las fases de adaptación y crecimiento exponencial. Dentro de los resultados de cantidad de biomasa para la concentración de 2,3%v, el tratamiento con 0,59mM de NaNO3 produjo 1,416g/l; cultivos con concentraciones de 0,59 y 1,18mM de NaNO3 tuvieron un comportamiento similar, aunque sus velocidades específicas de crecimiento máximo (μMAX) fueron diferentes (Tabla 2). Los cultivos con concentraciones de 1,76 y 2,35mM de NaNO3 presentaron fases de adaptación de 2 y 3 respectivamente, después de los cuales iniciaron su fase de crecimiento exponencial con mayores μMAX.
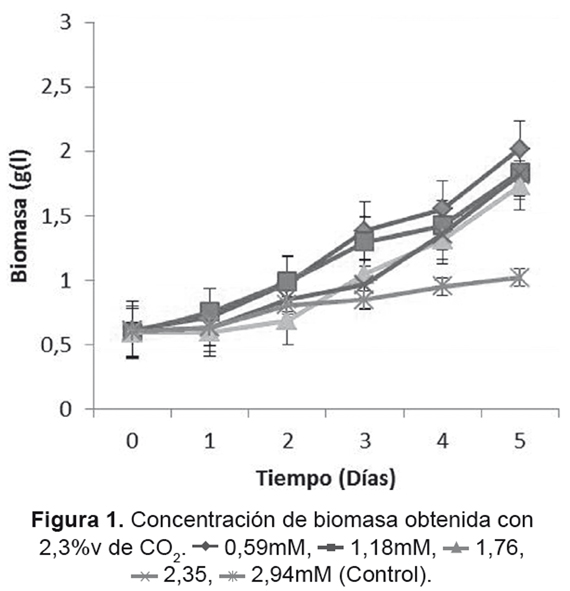
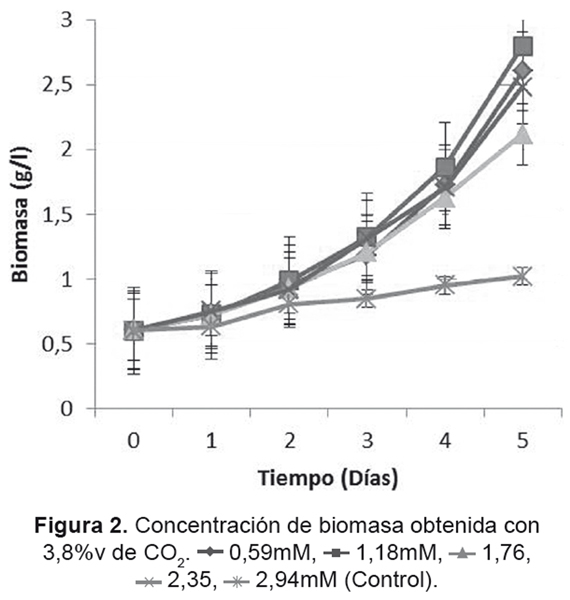
En la Figura 2 se observa el comportamiento para la concentración de 3,8%v de CO2, el cual obtuvo en general una mayor velocidad de crecimiento en comparación con los de 2,3%v; la mayor μMAX se encontró para el tratamiento con una concentración inicial de 1,18mM de NaNO3 (Tabla 3), generando 2,199g/L de biomasa. Cada cultivo tuvo un determinado día donde su crecimiento comenzó a ser exponencial, siendo un factor en la producción final de cada cultivo.
La producción de clorofila para los cultivos con una concentración de 2,3% v de CO2 presentó un estado de adaptación (Figura 3), siendo más largo para los cultivos con concentraciones iniciales de 1,18mM y 2,3 de NaNO3. Sin embargo, todos los cultivos aumentaron su cantidad de clorofila hasta el quinto día, esto se debe a que en el medio hay suficiente nitrógeno para suplir las necesidades fotosintéticas del alga. La mayor producción de clorofila se alcanzó en el cultivo con una concentración inicial de nitrógeno de 1,18mM.

A diferencia de los cultivos con 2,3%v de CO2, los cultivos con 3,8% tuvieron una fase de baja productividad durante los tres primeros días, esto ocurre debido al aumento en la cantidad de CO2 suministrado; sin embargo, el alga necesita estabilizarse a sus nuevas condiciones para que su metabolismo funciones correctamente [19]. Después del tercer día se observó un aumento en las tasas fotosintéticas de todos los experimentos con respecto a los días anteriores, indicando la adaptación a las condiciones (Figura 4). El cultivo con una concentración inicial de nitrógeno de 0,59mM de NaNO3 presentó una caída en su concentración de clorofila en el último día de experimentación, esto puede deberse a la falta de nitrógeno en el medio, ya que según Rodríguez-González AM & Serrano-Luna FA [54], hasta el 90% del nitrógeno del medio de cultivo es consumido durante los cinco primeros días de cultivo, por lo cual el alga aprovecha las reservas de este nutriente (principalmente en forma de proteínas y clorofila) para así mantener sus actividades metabólicas [19]. En este caso específico, el cultivo pasará de un estado autotrófico (donde la principal fuente de carbono proviene del CO2), a un estado heterotrófico, en el cual no dependerá de la clorofila, sino consumirá otras fuentes de carbono (en este caso carbohidratos extracelulares que la misma alga hubiese secretado en las primeras etapas de su crecimiento [55].

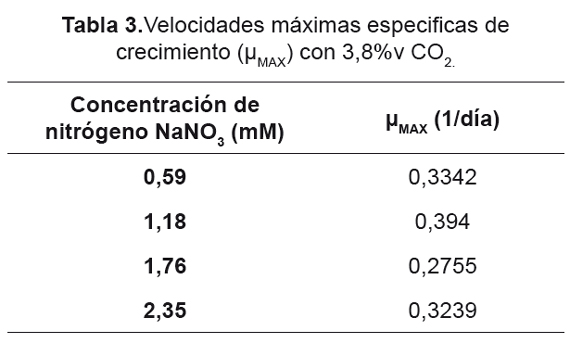
En la Figura 5 se presenta la productividad para los dos tratamientos de CO2, donde es posible destacar el aumento significativo en la productividad para cada uno de estos en comparación con el control, aunque el nitrógeno se considera como un factor determinante en la producción de biomasa [56] según Flynn [41]. El estrés generado por déficit de este nutriente afecta significativamente el crecimiento celular y el consumo de la fuente de carbono; esto se ve soportado con los datos obtenidos, donde al regular la relación entre el carbono y nitrógeno suministrado en el medio de cultivo es posible incrementar no sólo el consumo de la fuente de carbono (en este caso mejorar el secuestro de CO2 por parte de la microalga), sino a la vez aumentar la cantidad de biomasa. Otra explicación es debido a la fuerte relación entre la asimilación del nitrógeno y el metabolismo del CO2 [46,47].
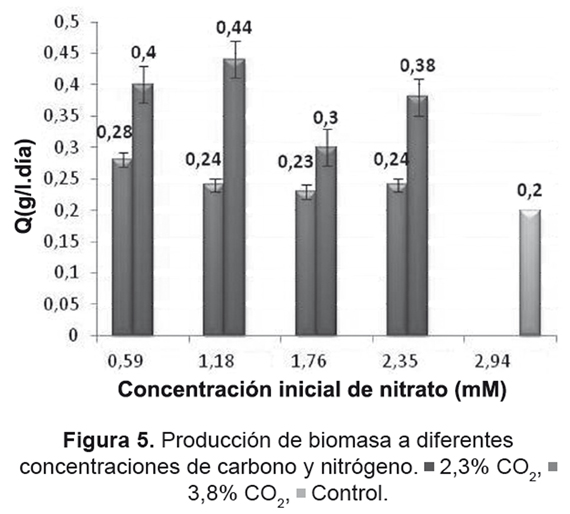
Debido a la obtención de resultados similares en la productividad de biomasa (0,4 y 0,44g/l.día) para los experimentos con las menores concentraciones de nitrato (0,59 y 1,18mM) y 3,8%v de CO2, se realizó un nuevo experimento con una concentración intermedia de nitrato (0,88mM); la productividad obtenida de este nuevo tratamiento fue de 0,43g/l.día (Figura 6), el cual es un valor intermedio entre los dos tratamientos originales. Este resultado confirma la existencia de un rango en la concentración inicial de nitrógeno que maximiza la producción de biomasa (entre 0,4 a 0,44g/l.día).
Para el análisis estadístico se realizaron 2 diseños factoriales de 2 factores con el fin de analizar el efecto que tienen los factores %CO2, concentración de NaNO3 y su interacción en las variables de respuesta Productividad (Q) y Clorofila (Ca+b).
Primero se analizó la influencia de los 2 factores en cada uno de los valores de productividad (Q) de biomasa de C. vulgaris.
Se aplicó una serie de pruebas de normalidad y un test de homogeneidad de varianza del efecto carbono/nitrógeno sobre la productividad para evaluar la normalidad del conjunto de datos a analizar. Utilizando la prueba de Kolmogorov- Smirnov, se puedo concluir que los datos de productividad presentan una distribución normal y homogeneidad en las varianzas (p>0,05). Por otro lado, los resultados de la prueba de análisis de varianza (ANOVA) permitieron inferir que las variaciones del %CO2 y de la concentración NaNO3 tienen efectos significativos en la Q (p<0,05) obtenida en los cultivos modificados de C. vulgaris, esto nos permite inferir que la relación entre el concentración de nitrógeno y carbono influye positivamente en el mejoramiento de la biomasa.
La regulación de la cantidad tanto de carbono como de nitrógeno suministrado afecta positivamente la producción de biomasa, donde bajas concentraciones de la fuente de nitrógeno (menos de 1,18mM) y altas concentraciones de carbono (3,8%v de CO2) mejoran hasta dos veces la productividad total de biomasa (de 0,2 a 0,44g/l.día). Otro fenómeno a resaltar es la no existencia de óptimos locales en la producción de biomasa, ya que con valores de hasta 0,59mM de NaNO3, es posible obtener altos valores de productividad. Estos resultados representan un avance importante en la producción de cultivos microalgales, ya que abre las posibilidades para mejorar la producción total de biomasa mientras se reducen costos operacionales y tiempos de producción, además permite desarrollar estrategias para acoplar la producción de diferentes metabolitos (ya sea para la obtención de biocombustibles como biodiesel o otros elementos) con el secuestro de CO2.
[1] Chae SR, Hwang EJ, Shin HS. Single cell protein production of Euglena gracilis and carbon dioxide fixation in an innovative photo-bioreactor. Bioresour. Technol. 2006;97(2):322-9. [ Links ]
[2] Kondili EM, Kaldellis JK. Biofuel implementation in east Europe: current status and future prospects. Renew. Sustainable. Energy. Rev. 2007;11(9):2137-51. [ Links ]
[3] Bilanovic D, Andargatchew A, Kroeger T, Shelef G. Freshwater and marine microalgae sequestering of CO2 at different C and N Concentrations â Response surface methodology analysis. Energy. Convers. Manage. 2008;50:262-7. [ Links ]
[4] Chinnasamy S, Ramakrishnan B, Bhatnagar A, Das KC. Biomass production potential of a Wastewater alga Chlorella vulgaris ARC 1 under elevated levels of CO2 and temperature. Int. J. Mol. Sci. 2009;10(2):518-32 [ Links ]
[5] Loera-Quesada MM, Olguín EJ. Las microalgas oleaginosas como fuente de biodiesel: retos y oportunidades. Rev. Latinoam. Biotecnol. Amb. Algal. 2010; 1(1):91-116. [ Links ]
[6] Jeong ML, Gillis JM, Hwang JY. Carbon dioxide mitigation by microalgal photosynthesis. Bull. Korean. Chem. Soc. 2003;24(12):1763-6. [ Links ]
[7] Um BH, Kim YS. Review: A change for Korea to advance algal-biodiesel technology. J. Ind. Eng. Chem. 2009;15(1):1-7. [ Links ]
[8] Sostarie M, Golob J, Bricelj M, Klinar D, Pivec A. Studies on the growth of chlorella vulgaris in culture media with different carbon sources. Chem. Biochem. Eng. 2009;23(4):471-7. [ Links ]
[9] Costa JAV, Linde GA, Atala DIP, Mibielli GM. Modelling of growth conditions for cyanobacterium Spirulina platensis in microcosms. World J. Microbiol Biotechnol. 2000;16(1):15â8. [ Links ]
[10] Crutzen PJ, Mosier AR, Smith KA, Winiwarter W. N2O release from agro-biofuel production negates global warming reduction by replacing fossil fuels. Atmos. Chem. Phys. 2008;8:389â95. [ Links ]
[11] Hughes E, Benemann JR. Biological fossil CO2 mitigation. Energy. Convers. Manage. 1997;38:467-73. [ Links ]
[12] Papazi A, Makridis P, Divanach P, Kotzabasis K. Bioenergetic changes in the microalgal photosynthetic apparatus by extremely high CO2 concentrations induce an intense biomass production. Physiol. Plant. 2008;132(3):338-49. [ Links ]
[13] Mallick N. Biotechnological potential of immobilized algae for wastewater N, P and metal removal: A review. Biometals. 2002;15(4):377â90. [ Links ]
[14] Jacome-Pilco CR, Cristiani-Urbina E, Flores- Cotera LB, Velasco-Garcia R, Ponce-Noyola T, Canizares-Villanueva RO. Continuous Cr(VI) removal by Scenedesmus incrassatulus in an airlift photobioreactor. Bioresour. Technol. 2009;100(8):2388â91. [ Links ]
[15] Hodaifa G, Martinez ME, Sanchez S. Daily doses of light in relation to the growth of Scenedesmus obliquus in diluted threephase olive mill wastewater. J Chem Technol Biotechnol. 2009;84:1550â8. [ Links ]
[16] Jacob-Lopes E, Revah S, Hernandez S, Shirai K, Franco TT. Development of operational strategies to remove carbon dioxide in photobioreactors. Chem. Eng. J. 2009;153(1):120â6. [ Links ]
[17] Ryu HJ, Oh KK, Kim YS. Optimization of the influential factors for the improvement of CO2 utilization efficiency and CO2 mass transfer rate. J. Ind. Eng. Chem. 2009;15(4):471â5. [ Links ]
[18] Pulz O. Photobioreactors: production systems for phototrophic microorganisms. Appl Microbiol Biotechnol. 2001;57(3):287â93. [ Links ]
[19] Richmond A. Handbook of microalgal culture-biotechnology and applied phycology. 4 ed. Vol. 4. Estados Unidos.: Wiley-Blackwell. 2004. [ Links ]
[20] Xiong WG. Double CO2 fixation in photosynthesisâfermentation model enhances algal lipid synthesis for biodiesel production. Bioresour Technol. 2010;101(7):2287-93. [ Links ]
[21] Lee YK, Pirt SJ. CO2 absorption rate in an algal culture: effect of pH. J. Chem. Tech. Biotechnol. 1984;34(1):28â32. [ Links ]
[22] Blanchemain A, Grizeau D. Eicosapentaenoic acid content of Skeletonema costatum as a function of growth and irradiance; relation with chlorophyll a content and photosynthetic capacity. J. Exp. Mar. Biol. Ecol. 1996;196:177â88. [ Links ]
[23] Molina Grima E, Fernández Sevilla JM, Sánchez Pérez JA, García Camacho F. A study on simultaneous photolimitation and photoinhibition in dense microalgal cultures taking into account incident and averaged irradiances. J. Biotechnol. 1996;45(1):59â69. [ Links ]
[24] Lee C. Calculation of light penetration depth in photobioreactors. Biotechnol. Bioprocess Eng. 1999;4:78-81. [ Links ]
[25] Melis A. Photosystem-II damage and repair cycle in chloroplasts: what modulates the rate of photodamage in vivo?. Trends. Plant. Sci. 1999;4(4):130â5. [ Links ]
[26] Ono E, Cuello JL. Selection of optimal microalgae species for CO2 sequestration. Disponible en: http://www.netl.doe.gov/publications/proceedings/03/carbon-seq/PDFs/158.pdf. Acceso en 5 de Enero de 2012. [ Links ]
[27] Luo HP, Al-Dahhan MH. Analysing and modeling of photobioreactors by combining first principles of physiology and hydrodynamics. Biotech Bioeng. 2004;85(4):382â93. [ Links ]
[28] Barber J. Andersson B. Too much of a good thing: Light can be bad for photosynthesis. Trends. Biochem. Sci. 1992;17:61â6. [ Links ]
[29] Merchuk JC, Gluz M, Mukmenev I. Comparison of photobioreactors for cultivation of the red microalga Porphyridium sp. J. Chem. Technol. Biotechnol. 2000;75(12):1119â26. [ Links ]
[30] Jacob-Lopes E, Scoparo CHG, Lacerda LMC, Franco TT. Effect of light cycles (night/day) on CO2 fixation and biomass production by microalgae in photobioreactors. Chem. Eng. Process. 2009;48(1):306â10. [ Links ]
[31] Hsueh HT, Chu H, Yu ST. A batch study on the bio-fixation of carbon dioxide in the absorbed solution from a chemical wet scrubber by hot spring and marine algae. Chemosphere. 2009;66(5):878â86. [ Links ]
[32] Carvalho AP, Malcata FX. Transfer of carbon dioxide within cultures of microalgae: plain bubbling versus hollow-fiber modules. Biotechnol. Prog. 2001;17:265â72. [ Links ]
[33] Morita M, Watanabe Y, Okawa T, Saiki H. Photosynthetic productivity of conical helical tubular photobioreactors incorporating Chlorella sp. under various culture medium flow conditions. Biotechnol. Bioeng. 2001;74(2):136â44. [ Links ]
[34] Babcock RW, Malda J, Radway JC. Hydrodynamics and mass transfer in a tubular air-lift photobioreactor. J. Appl. Phycol. 2002;14:169â14. [ Links ]
[35] Chiu SY, Kao CY, Chen CH, Kuan TC, Ong SC, Lin CS. Reduction of CO2 by a high-density culture of Chlorella sp. In a semicontinuous photobioreactor. Bioresour. Technol. 2008;99(9):3389â96. [ Links ]
[36] Keffer JE, Kleinheinz GT. Use of Chlorella vulgaris for CO2 mitigation in a photobioreactor J. Microbiol. Biotechnol. 2002;29(5):275-80. [ Links ]
[37] Yoo C, Jun SY, Lee JY, Ahn CY, Oh HM. Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour Technol. 2010; 01(1):71â 4. [ Links ]
[38] Yun YS, Lee SB, Park JM, Lee CI, Yang JW. Carbon dioxide fixation by algal cultivation using waste water nutrients. J. of Chem. Technol. Biotechnol. 1997;69(4):451-5. [ Links ]
[39] Chisti Y. Biodiesel from microalgae beats bioethanol. Trends. Biotechnol. 2008;26(3):126-31. [ Links ]
[40] Lv JM, Cheng LH, Xu XH, Zhang L, Chen HL. Enhanced lipid production of Chlorella vulgaris by adjustment of cultivation conditions. Bioresour. Technol. 2010;101(17):6797â804. [ Links ]
[41] Flynn K. Algal carbon-nitrogen metabolism: a biochemical basis for modelling the interactions between nitrate and ammonium uptake. J. of Plankton Research. 1991;13(1):373-87. [ Links ]
[42] Solovchenko A, Khozin-Goldberg I, Didi- Cohen, S, Cohen Z, Merzlyak M. Effects of light intensity and nitrogen starvation on growth, total fatty acids and arachidonic acid in the green microalga Parietochloris incisa. J of Appl Phycol. 2008;20(3):245-51. [ Links ]
[43] Takagi M, Watanabe K, Yamaberi K, Yoshida T. Limited feeding of potassium nitrate for intracellular lipid and triglyceride accumulation of Nannochloris sp UTEX LB1999. Appl Microbiol Biotechnol 2000;54(1):112-7. [ Links ]
[44] Flynn K. & Cunningham, D. Relations between Carbon and Nitrogen during Growth of Nannochloropsis oculata (Droop) Hibberd Under Continuous Illumination. New Phytologist. 1993;125(4):717-22. [ Links ]
[45] Huppe, H, Turpin D. Integration of carbon and nitrogen metabolism in plant and algal cells. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1994;45:577-607. [ Links ]
[46] Turpin D, Elriffi I, Birk D, Wegner H, Holmes J. Interactions between photosynthesis, respiration and nitrogen assimilation in microalgae. Canadian J of Botany, 1988;66(10):2083-97. [ Links ]
[47] Turpin D. Effects of inorganic N availability on algal photosynthesis and carbon metabolism. J. Phycol, 1991;27(1):14-20. [ Links ]
[48] Hitchcock G, Goldman C, Dennett M. Photosynthate partitioning in cultured marine phytoplankton metabolic patterns in a marine diatom under constant and variable light intensities. Marine Ecology Progress Series, 1986;30:77-84. [ Links ]
[49] Illman A, Scragg A, Shales S. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enz Microb Technol. 2000;27(8):631-5. [ Links ]
[50] Scragg A, Illman A, Carden A, Shales S. Growth of microalgae with increased calorific values in a tubular bioreactor. Biomass Bioenergy. 2002;23(1):67-73. [ Links ]
[51] Heredia-Arroyo T, Wei W, Hu B. Oil Accumulation via Heterotrophic/Mixotrophic Chlorella protothecoides. App Biochem Biotechnol. 2010;162(7):1978â95. [ Links ]
[52] Becker EW. Microalgae, Biotechnology and microbiology. Estados Unidos: Cambridge University Press. 1994. [ Links ]
[53] Fan LH, Zhang YT, Zhang L, Chen HL. Evaluation of a membrane-sparged helical tubular photobioreactor for carbon dioxide biofixation by Chlorella vulgaris. J. Membr. Sci. 2008;325(1):336-45. [ Links ]
[54] Rodríguez González AM, Serrano Luna FA. Efecto de la relación carbono/nitrógeno en la productividad de biomasa y lípidos en cultivos de Chlorella vulgaris UTEX 1803 en fotobiorreactores a escala de laboratorio. (Tesis de grado) Bucaramanga, Colombia: Universidad Industrial de Santander; 2012. [ Links ]
[55] Flemming HC, Wingender J. The biofilm matrix. Nature Rev. 2010;8:623-33. [ Links ]
[56] Xiong W, Li XF, Xiang JY, Wu Q. Highdensity fermentation of microalga Chlorella protothecoides in bioreactor for microbiodiesel production. Applied Microbiology and Biotechnol. 2008;78:29â36. [ Links ]
