











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
World cassava production has increased in the last years, reaching 270.2 million of tons by 2014 1. Starch from sour cassava is a cheap source to produce ethanol. Starch pretreatment process, involve the breaking of amylose and amylopectin by enzymes, being these the costliest step in the ethanol production from starch, around 47% of total value, mainly by the enzymes price 2. One example is the use of saccharolytic enzymes for the hydrolysis of starch in obtaining fermentable syrups for bio-ethanol production, which reduces time and generates fewer residues compared with traditional acid hydrolysis 3. Enzymatic extracts are commercially available for liquefaction and saccharification of corn and sorghum starch, highcost raw materials that compromise food security. However, for cassava starch, which is not suitable for human consumption, no commercial enzymes for hydrolysis exist in the market. In addition, sour cassavas abound greatly in Latin America. Glucose syrups can be obtained via chemical or enzymatic hydrolysis of starch 4. In chemical hydrolysis, the use of acids increases the cost of production, unwanted products are formed, and wastes can pollute the environment. In contrast, enzymatic hydrolysis is a cheaper alternative that produces less pollution. However, due to the complexity of the structure of starch, it is necessary to incorporate heat treatment and the use of different enzymes to ensure the highest conversion of this polymer to glucose 5.
The enzymatic hydrolysis of starch is carried out in two steps: liquefaction and saccharification. In the first step, α-amylase (EC 3.2.1.1) is used to make cuts at random into the internal structure of starch due to endo-enzyme activity, generating many products such as pullulan, dextrin and glucose. The pullulan and dextrin are not viable to be used in the alcoholic fermentation process because they are not absorbed by fermenting microorganisms. Saccharification can occur by two enzymes: pullulanase (EC 3.2.1.41), which only breaks α-1.6 bonds and produces mostly dextrins, disaccharides such as maltose and a very small amount of glucose; and glucoamylase (EC 3.2.1.3 and EC 3.2.1.20), which hydrolyzes α -1,4 glycosidic terminal bonds of dextrins from starch liquefaction, forming mainly glucose. The pullulanases are produced mostly by bacteria, and glucoamylases are produced by fungi 6. Enzymatic saccharification is expensive and thermally unstable. The enzymatic process is performed at high temperatures because this improves solubility, decreases viscosity of the starch, reduces microbial contamination, decreases the process time and reduces production costs 7. The bio-fuel industry seeks to replace the enzymes used in the industrial processes of starch hydrolysis with enzymes having higher thermostability 8. These enzymes can be obtained from wild type microorganisms because in some cases, native enzymes exhibit better catalytic activity than commercial enzymes 9.
For production of enzymes with saccharolytic activity from filamentous fungi, the genera Aspergillus and Rizhopus have been the most studied using solid state fermentation (SSF) 10. Native fungal enzymes also have been shown to viably be used for glucose obtaining on an industrial scale.
The aim of this project was the isolation, identification and selection of native filamentous fungi with saccharolytic activity, and to obtain an enzymatic extract from the optimal isolation to be used in glucose syrup production from maltose and cassava starch.
Materials and methods
For primary isolation of fungi, sour cassava (Manihot sp) was collected with evident fungal colonization from two farms in rural Barrancabermeja, Santander, Colombia. Ten grams of sour cassava was chopped, homogenized and diluted in 90ml of sterile saline solution. Each suspension from cassava homogenized dilutions were taken to obtain fungal colonies by surface culture. Sub-cultures were made to obtain axenic strains from the isolations at 25°C 11. The culture media used contained maltose (5g.L-1) as the only carbon source, rose bengal to restrict microbial growth and chloramphenicol (0.1g.L-1) as a broad-spectrum antibiotic.
Genus of Isolated fungi were identified by macro- and microscopic characteristics using taxonomic keys. The culture media for identification was potato dextrose agar (PDA).
Submerged fermentation (SF) was performed in flasks of 250mL in sterile conditions, each flask contained 50mL of basal media in deionized water with the following per L: maltose (5), KH2PO4 (5), MgSO4.7H2O (5), peptone (2) and yeast extract (1). SF was made to determine maltose consumption and production of glucose, protein and biomass. Each flask was adjusted to a final concentration of 105 propagules mL-1. Each batch of SSF isolations had twenty-five flasks, the fermentation was held with agitation at 150rpm and incubate at 25˚C for 72 hours. Every eight hours, one flask and two replicates were taken to obtain the enzymatic extracts, the biomass was separated from the broth using filtration through a Büchner funnel with Whatman No. 1 filter paper, and the enzymatic extract was obtained by vacuum.
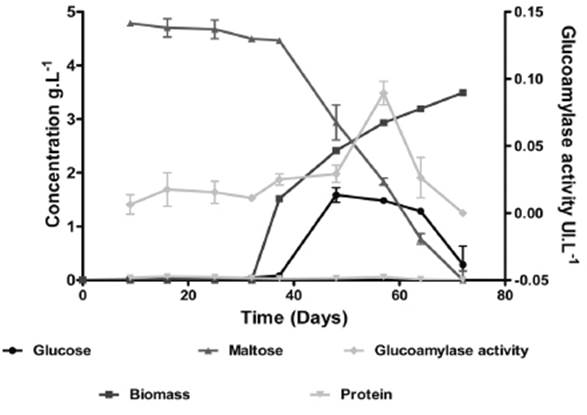
A first screening of the enzymatic extract was performed by evaluating maltose hydrolysis and glucose production using 3,5-dinitrosalicylic acid (DNS) 12 and glucose oxidase (Biosystems®) 13, respectively. Volumetric productivity was calculated based on glucose production. The maltose concentration was calculated via subtraction of the concentration of reducing sugars and glucose. The production of glucose, protein by Bradford method 14, and biomass by dry weight and the hydrolysis of maltose were determined in a second screening. The isolation with the best results was named Aspergillus A1. The enzymatic volumetric activity (UI.L-1) of the enzymatic extract from this isolation was quantified (Figure 1). The enzymatic volumetric activity (UI.L-1) was defined as micromoles of glucose produced per ml in one minute at 50°C and 200rpm.
To increase the protein concentration from the enzymatic extract, SSF of Aspergillus A1 was performed in a column bioreactor with a lignocellulosic support sterilized at 121°C for 15min. The column bioreactor was filled with 8cm of support and 150ml of broth. The crude enzymatic extract was obtained by pressing the lignocellulosic polymer, followed by centrifugation at 10000 × g for 15 minutes and finally filtering (Millipore, cellulose 0.45µm).
Electrophoresis SDS-PAGE was performed on enzymatic extracts obtained by SSF of Aspergillus A1 to determine the molecular weight of the proteins present in the extract. A Fermentas PageRuler of 10 to 200kDa was used as a molecular weight standard. Samples were run at 120 volts in 12% polyacrylamide gels using Coomassie blue as the dye.
Analysis of variance was performed to establish the highest values of enzymatic activity of the enzymatic extract from Aspergillus A1 at a pH 3-9, a temperature of 30-90°C and an agitation of 0-600rpm. The results of this analysis were used to determine the best experimental conditions for maltose hydrolysis. The independent variables were pH 4.0; 4.5 and 5.0; 16mmol.L-1 buffer sodium acetate and 0,1molL-1 citric acid; temperatures of 55, 60 and 65°C; and agitation at 0, 200 and 400rpm. The experiments were performed in tubes containing 1mL of enzymatic extract and 1mL of maltose at a concentration of 5mgmL-1.
The response variable was the glucose concentration versus time. Each assay was performed in triplicate, and the results were analyzed using Statgraphics software.
The effect of 1mmolL-1 aliquots of cofactors Cu2+, K+, Mg2+ and Na+ on enzymatic extract activity was determined per the methodology of Bhatti et al. 15. This was performed based on the optimized conditions found in the experimental design. The thermal stability of the enzymatic extract was also evaluated at 55°C through glucose quantification every 15 minutes for 3 hours.
Finally, the saccharolytic activity of the selected enzymatic extract was evaluated using the optimized parameters for the best cofactor on fermentable syrups obtained from liquefied cassava starch. Liquefied cassava starch was obtained according to the methodology described by Ruiz et al. 16.
Results and Discussion
From samples of sour cassava, it was possible to obtain 13 fungal isolations belonging to the genera Fusarium, Penicillum, Geotrichum, Scopularipsis and Aspergillus, and all the isolations showed saccharolytic activity. The two isolations with the best volumetric productivity were Aspergillus A1 and Aspergillus A12 with 0.07 and 0.05gL-1, respectively. The isolation that showed the best production of glucose from maltose was Aspergillus A1 (1.3gL-1) between 50 and 60 hours of fermentation. This agrees with the highest volumetric enzymatic activity from extract obtained at the same time, despite a low protein concentration, as shown in Figure 1. Based on the volumetric activity, glucose production from maltose hydrolysis was confirmed in all fungal isolations, with the fungi of genera Aspergillus having the best productivity. The previous results confirm that the Aspergillus genus had the highest glucose syrup production by submerged fermentation 17,18. Authors such as Riaz et al19. and O’Brien et al20, have suggested several species of Aspergillus, including A. awamori, A. foetidus, A. niger, A. oryzae, A. terreus, for glucose syrups obtaining from polysaccharides.
The enzymatic extracts obtained from submerged fermentation of Aspergillus A1 did not reach a higher concentration of protein (0.05g.L-1). This contrasted with SSF, where it was possible to obtain concentrations greater than 0.05g.L-1.
The increase in protein concentration in the enzymatic extract was possible with the change from submerged fermentation to solid fermentation with a lignocellulose support. With this variation, the protein concentration reached 0.06g.L -1. This is consistent with the work of Rodríguez et al21, and Singhania et al22, who concluded that use of SSF for obtaining enzymatic extracts is better than the concentrations achieved by submerged fermentation. The electrophoresis SDS-PAGE results of enzymatic extracts obtained from SSF of Aspergillus A1 showed 11 bands with different molecular weights. The band with a molecular weight between 60 and 70kDa had the highest intensity, as shown in Figure 2.
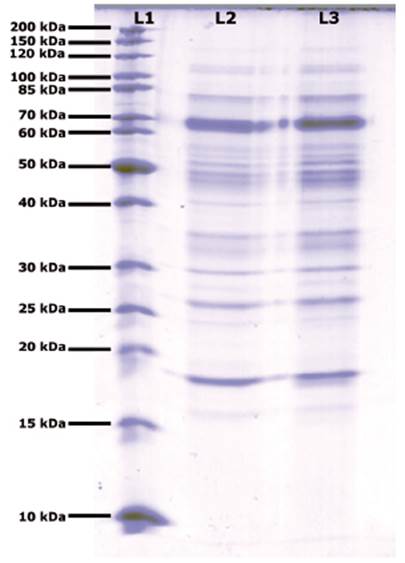
Figure 2 Polyacrylamide gel electrophoresis of enzymatic extract from Aspergillus A1 obtained by SSF. Line 1 Molecular weight marker; Lines 2 and 3 enzymatic extract from Aspergillus A1 from two different bioreactors. Coomassie brilliant blue G-250 staining.
The bands observed in gels from SDS-PAGE obtained from different SSF of enzymatic extracts from Aspergillus A1 were not different from each other, guaranteeing the reproducibility of SSF, as shown in Figure 2. The band with the highest intensity occurred between 60 and 70kDa. These results corroborate those of Norouzian et al.,8 who established the molecular weights of fungal glucoamylases to be within a range of 48 to 90kDa. da Silva et al23 and Bagheri et al24 reported the molecular weight of glucoamylase from Aspergillus niveus to be 76kDa.
Analysis of variance showed that the best saccharolytic activity of the extract at pH values between 4 and 5, agitation between 200 and 600rpm and a temperature of 60°C. By using these values, it was possible to define the variables for the experimental design matrix.
As can be seen in Figure 3, the best enzymatic extract activity from Aspergillus A1 with respect to pH occurs at 20 minutes, (4.4UI.mg-1) at a pH between 4 and 5 and at 60°C. Under these conditions, the activity reached 3.5µmol.L-1. These results are comparable to those of Bagheri et al24, who reported that the highest activity of glucoamylase occurs at a pH between 4.5 and 6.5 and at temperatures between 50 and 70°C.
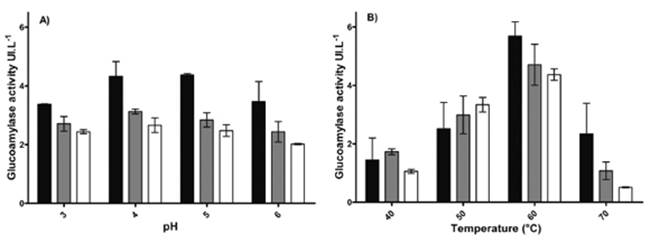
Figure 3 Glucoamylase activity under the effect of different values of pH 3 - 6 (A) and temperatura 40-70°C (B), using maltose to concentration 5mg.mL-1 like a substrate. (black bars) glucoamylase activity at 20min; (grey bars) glucoamylase activity at 40min and (white bars) glucoamylase activity at 60min.
The highest values of enzymatic activity and glucose obtained from maltose were at pH 4 (4.4UI.mg-1 and 6,8µmolmL-1, respectively) and a temperature of 60°C (5.6UI.mg-1 and 3.5µmol.mL-1, respectively), as shown in Figure 3.
From these experimental design results, seven effects were found with a P value lower than 0,05 and confidence level of 95%. The results indicate that there was little or no interaction between the temperature and agitation. This is contrary to what is normally observed regarding interactions of pH/ temperature and pH/agitation, with the latter typically being the most significant.
The polynomial obtained for glucoamylase activity (Equation 1) showed an R-squared value of 89.1% and an adjusted R-squared value of 87.7%, which could explain the variability in enzymatic activity with a standard error of 0.4% and an absolute error of 0.3%.

From the mathematical model obtained to predict enzymatic activity, pH and pH2 were the most influential factors. From the response surface and surface contour (Figure 4) it can be concluded that the best saccharolytic activity obtained by the enzymatic extract was at an agitation of 0rpm. Enzymatic activity was increased when the agitation increased to 4rpm at an average temperature of 55°C 25.
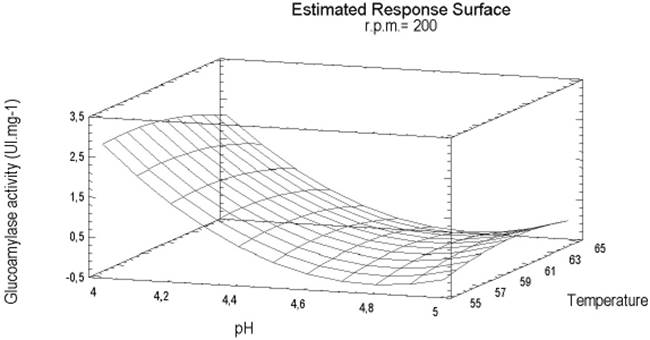
Figure 4 Response surface estimated of the combined effects of pH and temperature of enzymatic extract from Aspergillus A1, without agitation, 5mgmL-1 of maltose concentration at 15 minutes of reaction
Enzymatic cofactor evaluation showed that all cofactors evaluated had a positive influence on enzymatic activity. The Mg2+ ion was found to increase enzymatic activity by 32%, resulting in a value of 4.15UI.mg-1. Figure 5 shows that the use of metallic ions such as enzymatic cofactors increases enzymatic activity compared to the control. Mg+ was the most influential factor, resulting in a 32.4% increase in enzymatic activity. This result agrees with that reported by Bhatti et al 15 and Benassi et al 26, who found a 176% increase in enzymatic activity using manganese ions in glucoamylase from Fusarium phoenicis.
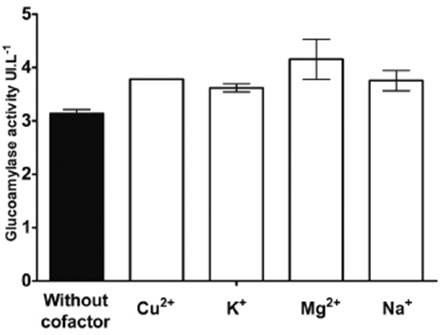
Figure 5 Effects of various metal ions at 1mmol concentration on glucoamylase activity, (black bar) control.
The thermal stability assay showed three periods of variation. The first 30 minutes of enzymatic activity peaked at 5.6UI.mg-1, followed by a decrease by 22%, and then the activity remained stable until 120 minutes. The enzymatic activity finally decreased to an average of 56% at the completion of the study.
The half-life determined for the enzymatic extract from Aspergillus A1 was 60 minutes of incubation at 60°C, which was a similar time to that obtained by Sutthirak et al27, who determined that the greatest decrease in enzyme activity occurred after two hours (Figure 6).
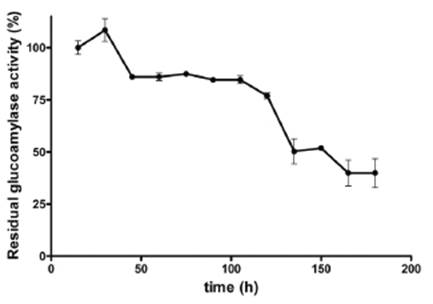
Figure 6 Thermal stability of enzymatic extract from Aspergillus A1, 5mg.ml-1 maltose concentration.
In saccharification assays of liquefied cassava starch, it has been found that saccharolytic activity has a glucose/enzymatic activity ratio of 0.9 for enzymatic extracts from Aspergillus A1. This is lower than that obtained by Ruiz et al16, who found a value of 3.7 for purified commercial glucoamylase Spirizyme ® Fuel.
From 13 fungal isolations identified from decomposed cassava, the Aspergillus genera had the highest saccharolytic activity. From this group, it was possible to obtain an enzymatic extract with glucoamylase enzyme activity.
The SSF make it possible to obtain a higher protein concentration in enzymatic extracts from Aspergillus A1 for quantification and observation of the proteins by Bradford assay and electrophoresis, respectively.
The best parameters for maltose hydrolysis by enzymatic extracts from Aspergillus A1 were a pH of 4.0; a temperature of 55°C without agitation and 1 mmol.L -1 of Mg2+ cofactor.
It was confirmed that the enzymatic extract from native Aspergillus sp. could undergo saccharification of liquefied cassava starch.

