











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCION
La raza criolla Hartón del Valle se originó de bovinos introducidos por los españoles alrededor de 1539; se explota principalmente en sistemas de doble propósito (leche y carne) y puede generar hasta el 65% de los ingresos por concepto de venta de leche y quesos (Casas y Valderrama, 1998).
El contenido total de proteínas en leche bovina es de alrededor de 35 g L-1, con el 80% representado por las caseínas αs1-, αs2-, β- y κ-CN, 10% por β- lactoglobulina, 2% por α-lactoalbúmina y pequeñas cantidades de enzimas e inmunoglobulinas (Muysson y Verrinder, 1989; Braunschweig et al., 2000). Las caseínas αs1- y αs2- constituyen aproximadamente el 48% de las caseínas totales, las β-caseínas el 35% y κ-caseínas el 13%. Las α y β caseínas contienen varios grupos serina fosfato, la k-caseína presenta muy pocos, razón por la cual tiene baja capacidad para unirse al calcio, lo que la hace insensible a la precipitación inducida de las proteínas lácteas (Muysson y Verrinder, 1989).
Las caseínas se precipitan a un pH bajo (4.5- 5.0), temperatura de 20ºC, seguida de una proteólisis en la porción hidrofílica de la κ-CN ocasionada por 44 quimosina o pepsina, lo cual permite la formación del cuajo, materia prima para la manufacturación del queso (Ginger y Grigor, 1999).
Las caseínas están codificadas por genes autosómicos estrechamente ligados (Chessa et al., 2003), lo que implica que la unidad de transmisión genética sea el haplotipo. Del gen de la κ-CN se han descrito seis variantes alélicas: A, B, C, E, F y G. La κ-CN está conformada por 169 aminoácidos, con regiones variables en los codones 136 y 148 del tercer exón; la variante A contiene treonina en el codón 136 (ACC) y ácido aspártico en el 148 (GAT), la variante B contiene isoleucina (ATC) y alanina (GCT). La leche bovina que contiene κ-CN del tipo B presenta contenido proteico más alto, mayor estabilidad al calor y a la congelación, menor tiempo de coagulación, cuajo más consistente y 5-10% más de rendimiento quesero (Barroso et al., 1998).En la detección de los genotipos AA, AB y BB se han utilizado muestras de leche, en hembras lactantes y en el caso de evaluación de machos se requieren 5 a 6 años para una prueba de progenie. Las técnicas moleculares han permitido identificar genotipos de interés en machos jóvenes.
La sustitución nucleotídica permite usar el exón como marcador de selección (Prinzenberg et al., 1996), con técnicas como PCR-RFLP (reacción en cadena de la polimerasa-polimorfismo en la longitud de los fragmentos de restricción) o PCR-SSCP (reacción en cadena de la polimerasa-polimorfismo en la conformación de ADN de cadena única). (Barroso et al., 1998). En la técnica PCR-RFLP se amplifica una región del gen de κ-CN y para la detección de las variantes alélicas se digiere con enzimas de restricción que son de alto costo. El método de detección de mutaciones PCR-SSCP (Orita et al., 1989) es una técnica rápida, sencilla y de bajo costo que se basa en la migración diferencial de cadenas desnaturalizadas de ADN, debido a que las cadenas simples adquieren estructuras secundarias complejas que dependen de la secuencia de nucleótidos (Hoelzel, 1998). Con los métodos PCR-RFLP y PCR-SSCP pueden identificarse variantes alélicas de forma rápida en muestras de semen o sangre, en animales de cualquier sexo y edad, lo que permite a los productores de hatos lecheros aumentar en corto tiempo la frecuencia del alelo B, mediante la utilización de reproductores homocigotos.
En esta investigación se estandarizaron y compararon las técnicas PCR-RFLP y PCR-SSCP para la identificación de los alelos de la κ-CN, en bovinos Hartón del Valle.
MATERIALES Y METODOS
Se utilizó sangre de 2 machos reproductores y 34 hembras de la raza de ganado criollo Hartón del Valle en dos hatos lecheros situados en Roldanillo (04º 24' 53'' latitud norte, 76º 09' 17'' longitud oeste) y dos unidades de producción animal pertenecientes a la Universidad Nacional de Colombia, en Palmira (03º 32' 22'' latitud norte, 76º 18' 13'' longitud oeste) y Candelaria (03º 24' 27'' latitud norte, 76º 21' 04'' longitud oeste) (Valle del Cauca, Colombia). Los individuos se seleccionaron al azar; las muestras de sangre se tomaron en tubos vacutainer® de 5 ml con anticoagulante EDTA, se conservaron refrigeradas y se extrajo el ADN mediante el protocolo de “Salting Out” (Miller et al., 1998); la valoración del ADN se realizó en geles de agarosa al 1.2% por comparación con concentraciones conocidas de ADN del bacteriófago lambda.
Por medio de PCR se amplificó un fragmento de 453 pb, situado en el tercer exón del gen de κ-CN del cromosoma 6 (Barroso et al., 1998). Se utilizaron 120 ng de ADN en una solución buffer de PCR que contenía 20 nmoles de cada primer (sentido, 5´ -TGT GCT GAG TAG GTA TCC TAG TTA TGG-3´; antisentido, 5´-GCG TTG TCT TCT TTG ATG TCT CCT TAG-3´); 2 mM de MgCl2, 200 μM de mezcla de dNTP (MBI fermentas) y 2 U de Taq polimerasa (InvitrogenTM) en un total de 100 μl. Las muestras se sometieron a un ciclo de desnaturalización (5 minutos a 94˚C) y luego a 35 ciclos (1 minuto 94˚C; 1 minuto 65˚C; 2 minutos 72˚C) con una extensión final de 72˚C por cinco minutos.
Para la PCR-RFLP se digirieron 15 μl de producto de PCR con 2U de cada una de enzimas de restricción HinfI, HaeIII y Tail (estereoisómero de MaeII) (Tabla 1). Las muestras se incubaron a 37˚C por 3 horas para las enzimas HinfI y HaeIII y a 65˚C para la enzima TaiI. La visualización de los fragmentos se realizó en geles de poliacrilamida al 12% (relación de acrilamida: N,N´-metilene-bisacrilamida de 29:1) que contenía 0.5XTBE (0.045 M tris-borato, 0.001 M EDTA, pH 8.0). La tinción de los geles se hizo con nitrato de plata (Sambrook y Russell, 1991) Para PCR-SSCP se mezclaron 2 μl de los productos de PCR con 8 μl de buffer desnaturalizante (0.05% Xylene-Cianole, 0.05% azul de Bromofenol, 5.5 mM de EDTA pH 8.0), se desnaturalizaron a 95oC por dos minutos y luego se enfriaron a 4˚C para evitar que el ADN se renaturalice. La electroforesis se realizó en una cámara Biometra (12 x 8 cm), en geles al 12% de poliacrilamida (relación de acrilamida: N,N´-metilenebis- acrilamida de 100:1), TBE 0.5X (0.045 M trisborato, 0.001 M EDTA, pH 8.0) y 5% de glicerol. Las muestras se corrieron durante 8 horas a 200 voltios en TBE 0.5X con temperatura regulada a 15oC. La tinción de los geles se hizo con nitrato de plata.
Se calcularon las frecuencias alélicas para evaluar si la muestra poblacional se encontraba en equilibrio Hardy-Weinberg (Klug y Cumming, 1999), usando la distribución chi-cuadrado.
RESULTADOS Y DISCUSION
PCR-RFLP: La enzima HinfI permitió diferenciar los alelos A y E de B y C (Tabla 1 y Figura 1). Con la enzima HaeIII se descartó la presencia del alelo E y con TaiI se descartó la presencia del alelo C. En el patrón de bandas generado, solo se encontraron alelos A y B y se identificaron 17 individuos homocigotos AA, 15 heterocigotos AB y 4 homocigotos BB.
Tabla 1 Longitud (pb) de los fragmentos obtenidos por digestión de los productos de PCR, con las enzimas de restricción Hinfl, Haelll y Tail.
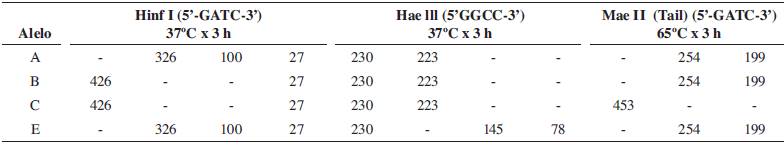
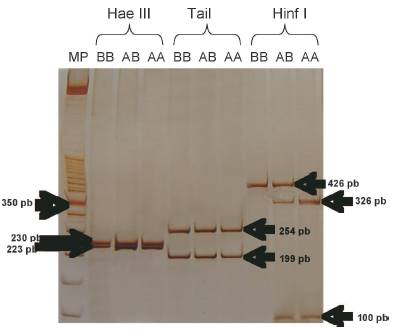
Figura 1 Patrón de bandas obtenido por amplificación del fragmento de 453 pb del gen de la k-CN, digerido con las enzimas HinfI, HaeIII y Tail.
En la digestión con la enzima HinfI se observó una banda inespecífica alrededor de los 250-300 pb (Figura 2), cuando se utilizó 5U de enzima de restricción por 15 μl de producto de PCR, de acuerdo con la metodología descrita por Barroso et al . (1998); esta banda desapareció al reducir a 2U de enzima.
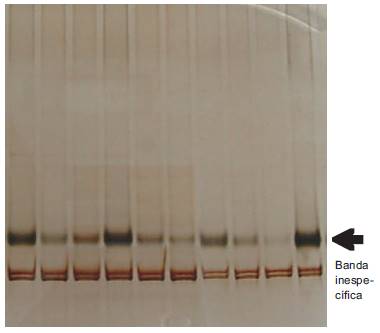
Figura 2 Digestión del fragmento de 453 pb la k-CN, con 5U de la enzima HinfI, donde se muestra la presencia de una banda inespecífica.
PCR-SSCP: Se detectaron dos patrones de movilidad que corresponden a los alelos A y B (Figura 3). En los geles PCR-SSCP cada cadena de ADN migra de manera independiente. El alelo A de k-caseína genera dos bandas que se diferencian por su ubicación en el gel, de las dos del alelo B; los genotipos homocigotos, por lo tanto presentan dos bandas y los heterocigotos cuatro bandas. Se detectaron 17 individuos homocigotos AA, 15 heterocigotos AB y 4 homocigotos BB, lo que coincidió perfectamente con la técnica PCR-RFLP.
Figura 3 Patrones de corrida de las variantes alélicas de k-CN obtenidas por PCR-SSCP en ganado Hartón del Valle donde se presentan los genotipos homocigotos AA t BB y el heterocigoto AB.
Se detectaron claramente las variantes alélicas en fragmentos de 453 pares de bases, cuando la literatura menciona que la PCR-SSCP es más eficiente para fragmentos entre 176 y 354 pb y enfatizan en la importancia de mantener la temperatura entre 4˚C y 10˚C con cámaras de electroforesis con sistema de enfriamiento. La PCR-SSCP se simplificó en aspectos técnicos pues se utilizó una cámara Biometra pequeña con dimensiones 12cm x 8 cm, sin sistema de refrigeración, lo cual permitió reducir los costos asociados al tamaño de los geles de poliacrilamida como el de los reactivos para la elaboración del gel, fijación, tinción y revelado, además de los costos de refrigeración, así como un equipo más económico. La detección de las variantes se logró con electroforesis a 200 v durante cuatro horas, por lo cual se disminuyó el tiempo de 14 horas recomendado por otros autores (Barroso et al, 1998). Para una detección óptima se puede aumentar a ocho horas, pero para efectos diagnósticos solo se requirieron cuatro horas. Aunque algunos autores (Fan et al., 1993; Cai y Touitou, 1993; Barroso et al., 1998) recomiendan la purificación de los productos de PCR para PCR-SSCP con el fin de evitar exceso de primers o iniciadores que puedan alterar los patrones de bandas, no fue necesario realizarlo, lo que redujo los costos de la técnica.
La detección de los alelos de la k-caseína utilizando PCR-SSCP tiene la ventaja con relación a la técnica PCR-RFLP que se evita la digestión del fragmento amplificado con tres enzimas de restricción HinfI, HaeIII y Tail proceso costoso por el alto precio de las enzimas y porque cada digestión debe hacerse de manera independiente.
Equilibrio Hardy-Weimberg: Las frecuencias alélicas fueron: f(A) = 0.681, f(B) = 0.319. La población muestreada resultó en equilibrio (P ≤ 0.05), aunque el tamaño de muestra es pequeño para hacer inferencias poblacionales. La frecuencia del alelo B (0.319) en Ganado Hartón del Valle fue mayor que en Holstein en EEUU (0.18) (Van Eenennaan y Medrano (1991) y Colombia (0.24) (López et al., 1999). Similar a Bovinos Criollos de Argentina (0.30) (Poli et al., 2002) y menor a los Bovinos Criollos de Uruguay (0.47) Postiglioni et al, (2002), Bovinos Criollos de Argentina (Liron et al., 2002) y a Criollo de la Patagonia (0.395) y Bolivia (0.645) (Liron et al., 2002).
Mediante técnicas moleculares es posible identificar animales portadores del genotipo de interés de cualquier sexo y edad, en forma rápida, lo cual representa una ventaja para los propietarios de hatos lecheros. El incremento de la frecuencia alélica de la variante B es una de la vías para mejorar la eficiencia lechera para su transformación en queso. Para una raza que presente una frecuencia alélica de 0.25 para el alelo B de la kappa-caseína, es posible aumentar la frecuencia alélica si solo se utilizaran toros con el genotipo BB; la frecuencia de este alelo ascendería al 60% en la primera generación y se situaría alrededor del 100% en pocas generaciones (Prinzenberg et al., 1996).
Se recomienda la realización del estudio poblacional completo, utilizando la técnica SSCP, ya que se conocen los patrones de bandas que se obtienen para los alelos A y B y utilizar enzimas de restricción únicamente en el caso de que se obtenga un patrón de bandas diferente del reportado, para reducir drásticamente los costos.

