










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.56 no.3 Palmira July/Sept. 2007
Resumen
La investigación se realizó en la vereda Cruz de Amarillo, municipio de Pasto (1º 7' norte y 77º 17´ oeste, 2.850 msnm, 12º C y humedad relativa del 83%). Se seleccionaron tres usos del terreno: praderas de Lollium multiflorum (ryegrass) y Pennisetum clandestinum (kikuyo) y un bosque secundario. En cada uno se trazó una línea recta y se tomaron ocho muestras, separadas 10 m, cada 15 días. El muestreo se hizo siguiendo la metodología del Tropical Soil Biology and Fertility Programe (TSBF). La fauna se recolectó manualmente y posteriormente se hizo el conteo y la valoración de la biomasa; los organismos se identificaron hasta nivel de familia y en algunos casos sólo hasta orden. También se determinó humedad gravimétrica, porosidad y contenido de materia orgánica. Los datos se analizaron de acuerdo con un diseño de bloques completos al azar. La pradera de L. multiflorum y el bosque secundario presentaron los mayores valores de abundancia de lombrices (5.648 y 4.864 ind m-2) y biomasa (141.3 y 670.7 g.p.f.m-2.) En la pradera de P. clandestinum abundaron los ácaros (2.768 ind m-2) con una biomasa de 0.22 g. p. f m-2. En todos los usos del suelo la mayor abundancia de organismos y biomasa se presentó en la profundidad de 0 – 10 cm.
Palabras claves: Typic melanudands; bosque secundario; Pennisetum clandestinum; Lolium multiflorum
ABSTRACT
At the county of Cruz de Amarillo, municipality of Pasto, Colombia, located al 2850 masl, a field study to determine the numbers of organisms and its biomass was carried out. Three land use systems were selected: prairie with the grass Lolium multiflorum (Ryegrass), pasture with Pennisetum clandestinum (kikuyu) and secondary forest. The prairie of .L multiflorum and the secondary forest showed the highest values of abundance of earth worms (5648 and 4864 ind m-2, respectively). Biomass was 141.3 and 670.7 gpfm-2 for L. multiflorum and P. clandestinum, respectively. Mites were abundant (2768 ind m-2) with a biomass of 0.22 gpfm-2. At all land use systems, the highest abundance of organisms and biomass at 2 depth of 0-10 cm was found.
Key words: Typic melanudands; secondary forest; Pennisetum clandestinum; Lolium multiflorum.
El sistema suelo es el resultado de complejas interacciones entre factores físicos, químicos y biológicos. Lavelle y Kohlman (1984) proponen un modelo jerárquico general que describe la importancia potencial de estos factores como determinantes de los procesos del suelo. En la escala más baja del modelo se encuentran los sistemas de regulación biológica. Los tres componentes de estos sistemas son la fuente de energía (materia orgánica fresca), la población de descomponedores (comunidades microbiales) y la población de macroorganismos reguladores (raíces vivas e ingenieros del ecosistema: lombrices y termitas). Cuando se remplaza un bosque por un cultivo, disminuye en forma dramática la diversidad de la fauna, siguiendo la misma tendencia de la diversidad vegetal; un área de bosque de diferentes edades, rastrojos, potreros y cultivo es un mosaico de hábitats, cada uno con su conjunto propio de especies (Higgins, 1994).
La macrofauna del suelo tiene diferentes efectos sobre los procesos que determinan la fertilidad del suelo y el crecimiento de las plantas (Lavelle, 1995). Influye en tres procesos que determinan la fertilidad del suelo: (I) la descomposición y la dinámica de la materia orgánica, (II) la formación y el mantenimiento de la estructura del suelo y, (III) la disponibilidad para las plantas de los nutrientes y del agua. La diversidad y la abundancia de las comunidades de organismos y la importancia relativa de los grupos de mayor interés (lombrices y termitas) se pueden usar como indicadores de calidad de suelo (Decaens et al., 1998).
Por las razones expresadas el trabajo tuvo como objetivo comparar los cambios de la edafofauna y de otras características biológicas por efecto de distintos usos del suelo
La investigación se realizó entre agosto y noviembre de 2005, en la vereda Cruz de Amarillo, municipio de Pasto (1º 7' 51.17 norte y 77º 17' 47.29 oeste, 2.850 msnm, 12º C y humedad relativa del 83% - Corponariño, 2004). Los suelos pertenecen a la Consociación Guadalupe (GA) y se clasifican como Typic melanudand, desarrollados a partir de cenizas volcánicas, muy profundos, bien drenados, de relieve quebrado a fuertemente quebrado; reacción positiva al fluoruro de sodio (NaF), textura franca a franco-arenosa muy friable; pH varía entre 4.0 y 5.7, con tendencia a aumentar con la profundidad; alto contenido de carbono orgánico, alta capacidad de intercambio de cationes y de saturación del complejo de cambio (IGAC, 2005).
El muestreo se hizo siguiendo la metodología del Tropical Soil Biology and Fertility Programe (TSBF) (Anderson e Ingram, 1993). Los usos del terreno seleccionado fueron: pradera con Pennisetum clandestinum (kikuyo, colchón cespitoso de 15 a 30 cm, Crowdwer, 1960), pradera con Lollium multiflorum (Ryegrass) y bosque secundario. En cada uno se trazó una línea recta y se tomaron ocho muestras, separadas 10 m, cada 15 días. Se tomó un volumen de suelo de 25x25x30 cm,dividiendo el monolito en cuatro estratos: mantillo (cuando estuvo presente) 0 – 10 cm, 10 – 20 cm y 20 – 30 cm. La fauna se recolectó manualmente y se preservaron las lombrices de tierra en formaldehído al 5% y los otros organismos en alcohol al 70%. Posteriormente se hizo el conteo y la valoración de la biomasa. Los organismos se identificaron hasta nivel de familia y en algunos casos solo hasta orden. En submuestras se determinó la humedad gravimétrica, porosidad y contenido de materia orgánica.
Los datos se analizaron de acuerdo con un diseño de bloques completos al azar, con dos factores: tipo de uso del terreno y profundidad de muestreo. Para las variables que presentaron diferencias estadísticas significativas se utilizó la prueba de Tukey (Little y Jackson, 1987). Se graficaron las interacciones entre variables y se hizo un análisis de correlación entre la variables estudiadas. Los datos de abundancia y biomasa de lombrices y artrópodos se transformaron por medio de la expresión
Abundancia y biomasa de organismos del suelo
El terreno cubierto por el bosque secundario presentó la mayor abundancia de organismos (Tabla 1) y la biomasa (Tabla 2) estuvo dominada por las lombrices, quizá porque ofrece alimento y condiciones favorables de habitación como sombra, alta humedad y materiales vegetales en distinto grado de descomposición (mantillo). La pradera de Ryegrass ocupó el segundo lugar en lombrices y biomasa de lombrices. En la pradera de kikuyo predominó el orden acarina y en la biomasa también dominaron las lombrices.
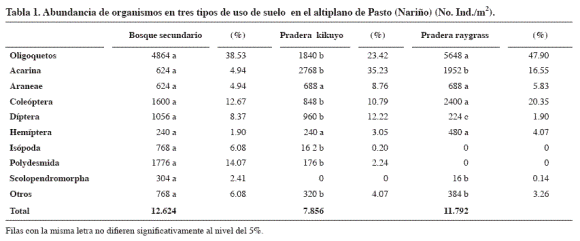
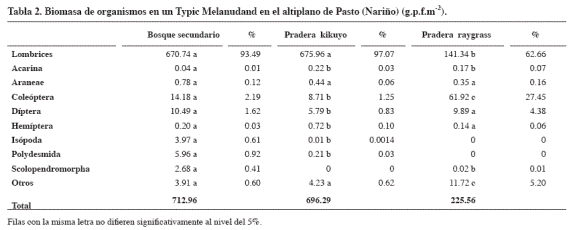
En los tres usos del terreno el segundo lugar en cuanto a biomasa lo ocupó el grupo de los coleópteros.
En el bosque secundario el 38,5% de la comunidad estuvo representado por oligoquetos (lombrices), seguido por el orden Polydesmia (14.07% ) con cuatro familias: Cryptodesmidae, Cryptopidae, Chelodesmidae y Paradoxomathidae. Los Coleópteros se agruparon en 10 familias: Carabidae, Cucujidae, Curculionidae, Elateridae, Lampyridae, Melolonthidae, Ptillidae, Scolytidae, Scydmaenidae, Staphylinidae; los Dípteros en tres familias (Bibionidae, Chironomidae, Tipulidae); los Isopodos en dos familias (Oniscidae, Porcellionidae). Otros órdenes que contribuyeron fueron Diplura, Geophilomorpha, Glomeridesmida, Hymenoptera, Lepidoptera, Neuroptera, Opilionida, Psocoptera, Scolopendromorpha y Thysanoptera.
En la pradera de kikuyo el 35.2% de la comunidad estuvo representado por el orden acarina, seguido por oligoquetos (23.42%). En Aranae se determinaron tres familias: Dipluridae, Salticidae, Tetragnathidae.
En la pradera de Ryegrass, el 47.9% de la comunidad estuvo representada por oligoquetos (lombrices), seguido por Coleópteros (20.3%). Los Hemiptera pertenecieron a las familias Cercopidae y Cicadellidae. La categoría otros agrupó Hymenoptera, Delphacidae, Enicocephalidae, Pseudococcidae Lepidoptera, Neuroptera, Scolopendromorpha y Thysanoptera.
El análisis de varianza mostró diferencias significativas entre los usos de suelo para la variable abundancia de lombrices. Según la prueba de Tukey el número de individuos fue significativamente diferente entre el bosque secundario y la pradera de kikuyo. No hubo diferencias estadísticas entre la pradera natural (kikuyo) y la pradera establecida (Ryegrass). Esta variable presentó una correlación altamente significativa con la humedad gravimétrica (r2: 0.63), porosidad total (r2: 0.54), materia orgánica (r2: 0.50) y es inversa con la densidad aparente (r2: -0.62). Cortés et al. (1990) no encontraron lombrices en andisoles de la sabana de Bogotá cubiertos por plantaciones forestales pero sí estuvieron presentes en bosque nativo y praderas.
Igualmente se presentaron diferencias altamente significativas para la abundancia de artrópodos en las distintas profundidades del suelo. El mayor valor correspondió al mantillo respecto de la profundidades 0 - 10 cm y 20 - 30 cm. También se encontraron diferencias entre las profundidades 0 - 10 y 20 - 30 cm.
La biomasa de artrópodos varió significativamente con la profundidad del perfil. Se presentó mayor peso en el mantillo que en la profundidad 10 - 20 cm. También hubo diferencias entre el mantillo y la profundidad 20 - 30 cm. De acuerdo con el análisis de varianza para la biomasa de oligoquetos hubo diferencias significativas entre los usos de suelo. Según la prueba de Tukey la pradera natural (kikuyo) presentó mayor valor que la pradera establecida (Ryegrass). La contribución de las lombrices a la biomasa puede atribuirse posiblemente a la profusión y renovación de raíces en los pastos y, en el bosque, a los buenos contenidos de materia orgánica que permiten baja densidad aparente y por ende buena circulación de aire y humedad para el desarrollo.
Los resultados ponen de manifiesto que el cambio en el uso del terreno puede generar variación en las poblaciones edáficas como respuesta a modificaciones en la cobertura vegetal, radiación solar, lluvia y propiedades físicas y químicas del suelo (Peñaranda y Naranjo, 1988). En el bosque secundario con zonas intervenidas se evidenció mayor densidad de organismos, lo que lleva a pensar que las condiciones edáficas y de microclima son adecuadas para la macrofauna existente, así como que la capa de materia vegetal en diversos grados de descomposición ofrece alimento, protección y hábitat para la macrofauna. La ausencia, disminución o cambio en los sitios intervenidos se debe al efecto antrópico.
El cambio de uso de la tierra de bosque a pradera ejerció un efecto negativo sobre la macrofauna del suelo al disminuir la riqueza de especies (25, 16 y 13 unidades taxonómicas) y aumentar la abundancia de algunos grupos tales como acarina, capaces de adaptarse a las nuevas condiciones.
En lombrices de la familia Glossoscolecidae de una región de los Andes del Valle del Cauca, en sitios intervenidos drásticamente las especies endémicas retrocedieron y los espacios quedaron disponibles para la colonización de las cosmopolitas; aunque algunas especies de mayor plasticidad ambiental se sobrepusieron a la alteración (Feijoo y Quintero, 1998).
Los resultados indicaron que la macrofauna del suelo se desenvuelve en el mantillo y en la profundidad de 0 - 10 cm, ya que a medida que se profundiza en el perfil del suelo disminuye el contenido de oxígeno y de materia orgánica que proporciona el hábitat y alimento para desarrollarse, por otra parte el desarrollo se limita en condiciones anóxicas.
Henrot y Brussard (1997) encontraron mayor peso de las lombrices en pastos que en cultivos y selvas tropicales; probablemente por la alta biomasa de raíces en las praderas.
Remy y Dainar (1982); Nuutinen (1992), Vikram et al. (1994) y Hendrix et al. (1994) afirman que la biomasa de artrópodos se favorece cuando la labranza no se realiza o es menos intensa, y atribuyen lo encontrado a la perturbación causada en el hábitat y a otras operaciones de campo como la aplicación de agroquímicos. Resultados que concordaron con lo encontrado en este estudio donde la mayor biomasa se obtuvo en el bosque, seguido de la pradera natural (kikuyo) y por último la pradera establecida (RayGrass).
- El ecosistema que presentó la mayor abundancia y biodiversidad fue el bosque secundario.
- El cambio de uso de suelo de bosque a pradera disminuyó la riqueza de especies y aumentó la abundancia.
A la Universidad de Nariño y Universidad Nacional de Colombia por el apoyo en la realización de la tesis de Maestría del primer autor, de donde se derivó la información para la redacción del artículo.
1. Anderson J.; Ingranm, J. 1993. Tropical soil biology and fertility: A Handbook of methods. Wallingford, CAB. pp. 44 – 46. [ Links ]
2. Corponariño. 2004. Actualización del diagnóstico físico-biótico y socioeconómico y formulación del plan de ordenamiento y manejo integral del río Bobo en el departamento de Nariño. Pasto. 120 p. [ Links ]
3. Cortés, A.; Chamorro, C.; Vega A. 1990. Cambios en el suelo por la implantación de praderas, coníferas y eucaliptos en un área aledaña al embalse del Neusa (páramo de Guerrero). Biol Suelo (IGAC). 2 (1):101-114. [ Links ]
4. Crowder, L.V. 1960. Gramíneas y leguminosas forrajeras de Colombia. Bogotá: División de Investigación Agropecuaria. 111 p. (Boletín Técnico No. 8). [ Links ]
5. Decaens, T.; Jimenes, J.; Schneidmald, J.; Lavelle, P. 1998. La macrofauna del suelo en sistemas de producción agrícola: Respuesta a las perturbaciones y perspectivas de manejo. Un caso de estudio en los Llanos Orientales de Colombia. Suelos Ecuatoriales (Colombia) 28: 262 – 268. [ Links ]
6. Feijoo, A.; Quintero, H. 1998. Glossoscolecidae de una región de los Andes del departamento del Valle, Colombia. Suelos Ecuatoriales (Colombia) 28: 249 – 253. [ Links ]
7. Hendrix, P.; Mueller, B.; Bruce, R.; Langdale, G.; Parmelee, R. 1992. Abundance and distribution of earthworms in relation to landscape factors on the Georgia piedmont, U.S.A. Soil Biol Biochem 24 (12): 1357 – 1361. [ Links ]
8. Henrot, J.; Brussaard, L. 1997. Abundance, and cast quality of earthworms in an acid ultisol under aller-cropping in the humid tropics. Appl Soil Ecol 6: 169 – 179. [ Links ]
9. Higgins, M. 1994. La entomofauna como indicador de la diversidad biológica. Diversidad biológica y diálogo de saberes. Memorias del curso de campo sobre biodiversidad y recursos genéticos indígenas y campesinos. Cali, Colombia. pp 49 – 56. [ Links ]
10. Instituto Geográfico Agustín Codazzi (IGAC). 2005. Estudio general de suelos y zonificación de tierras, departamento de Nariño. Bogotá, IGAC. 733 p. [ Links ]
11. Lavelle, P. y Kohlman, B. 1984. Etude quantitative de la macrofaune do sol dans une forest tropicale. Pedobiologia, 24: 834 – 845. [ Links ]
12. Lavelle, P. 1995. Faunal activities and soil processes: adaptative strategies that determine ecosystem function ISSS Congress, 15th, Acapulco, México. [ Links ]
13. Little, T.; Jackson, H. 1987. Métodos estadísticos para la investigación en la agricultura. México, Trillas. 265 p. [ Links ]
14. Nuutinen, V. 1992. Earthworm community response to tillage and residue management of different soil types in southern Finland. Soil Till Res, 23: 221 – 239. [ Links ]
15. Peñaranda, M.R.; Naranjo, G.M. 1998. Composición y variación de la edafofauna de un oxisol (Petroférrico acroperox) del complejo migmático de Mitú bajo tres usos diferentes del suelo. Suelos Ecuatoriales (Colombia) 28: 273 – 277. [ Links ]
16. Remy, E.A.D.; Dainar, T.B. 1982. Effects of tillage methods on earthworm population in monoculture corns. Canadian J Soil Sci, 62: 699 – 703. [ Links ]
17. Vikram, M.; Kiran, V.P.; Ravinder, V.; Balashouri, P.; Yule, D.; Cogle, A.; Jangawad, L.S. 1994. Earthworms biomass response to soil management in semi-arid tropical Alfisol agroecosystems. Biol Fertil Soils, 19: 317 – 321. [ Links ]

