










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.57 no.1 Palmira Jan./Mar. 2008
RESUMEN
Se realizó la evaluación de poblaciones segregantes producidas a partir de cruzamientos entre tomate S. lycopersicum L. cv. maravilla y la accesión silvestre PI134418 de S. habrochaites var. glabratum resistente al pasador del fruto N. elegantalis. Se encontró que hay dominancia de los alelos que disminuyen la expresión del caraacute;cter peso promedio de fruto. El método del retrocruzamiento resultó maacute;s eficiente que el de la autofecundación, para contrarrestar la dominancia de los alelos que codifican por el tamaño pequeño del fruto. Los tricomas y el número promedio de frutos por racimo no ejercieron ningún efecto sobre el ataque de los frutos por parte del pasador; mientras que el peso promedio de fruto sí tuvo efecto importante sobre dicho ataque, de tal manera que el insecto tiende a producir mayor daño a medida que se incrementa el peso promedio de fruto. Las poblaciones RC1 y RC2 se comportaron como las típicas segregantes para los caracteres evaluados y exhibieron amplia variabilidad en los mismos. El método del retrocruzamiento fue efectivo para romper la asociación negativa entre el peso promedio de fruto y la resistencia al insecto. El peso promedio de algunos de los frutos en los recombinantes RC2 fluctuó entre 45.1 y 68.6 g y mostraron resistencia al pasador.
Palabras claves: Solanum lycopersicum, Neoleucinodes elegantalis, autofecundación, tricomas, dominancia.
ABSTRACT
Evaluation of segregating populations from crosses between cultivated tomato S. lycopersicum L. cv. maravilla and PI134418 wild accession of Solanum habrochaites var. glabratum with resistance to the fruit borer N. elegantalis was made. There was dominance of alleles that decrease the expression of the character mean fruit weight. The backcrossing method was more efficient than selfing in order to counteract the dominance of alleles that encode by small fruit size. The thrichomes and the mean number of fruit per bunch did not produce any effect on the fruit attack by the fruit borer; while that mean fruit weight had important effect on this attack, therefore, the insect tends to produce more damage with the increasing in the mean fruit weight. The RC1 and RC2 populations showed segregating typical behavior for evaluated characters, presenting high variability in all of them. The backcrossing method was effective to break the negative association between mean fruit weight and the resistance to the insect. The mean fruit weight in the RC2 recombinant was between 45.1 and 68.6 g and showed resistance to the fruit borer.
Key words: Solanum lycopersicum, Neoleucinodes elegantalis, selfing, thrichomes, dominance.
El pasador del fruto Neoleucinodes elegantalis es uno de los insectos plaga que limitan severamente la producción de tomate S. lycopersicum en Colombia, Venezuela, Ecuador y Brasil (Vallejo, 1999; Marcano, 1991; Eiras y Blackmer, 2003).
Luckwill (1943) describió en el género Solanum sección Lycopersicum siete tipos de tricomas, los no glandulares (tipos II, III y V) y los glandulares (I, IV, VI y VII). El tipo VI presente en la especie S. habrochaites tiene cabeza multicelular y tiene importancia enorme como factor de resistencia a insectos. La densidad de ese tipo de tricomas es una característica valiosa para los programas de resistencia varietal a insectos.
Williams et al. (1980) reportaron que la 2-tridecanona (2-TD) es el principio activo de la resistencia a insectos en la accesión PI 134417 de S. habrochaites var. glabratum. La 2-TD estaacute; ausente en el interior de las hojas y se encuentra concentrada en la superficie de las mismas en los tricomas glandulares. Aragao et al. (2000) encontraron alta correlación positiva entre la cantidad de 2-TD y la densidad de tricomas tipo VI y alta correlación negativa entre el daño causado por la arañita roja Tetranycus urticae y las variables densidad de tricomas tipo VI y contenido de 2-TD.
Neves et al. (2003), al evaluar cinco progenies F3 de cruzamientos entre el cultivar de tomate IPA-6 y la accesión PI134418 de S. habrochaites var. glabratum, reportaron correlación negativa entre el daño causado por el cogollero Tuta absoluta y la densidad de tricomas tipo VI, y sugirieron que la resistencia al cogollero estaacute; relacionada con la alta cantidad de ese tipo de tricomas.
Restrepo, Vallejo y Lobo (2006) al evaluar germoplasma silvestre de Solanum sección Lycopersicum, identificaron cinco accesiones de la especie S. habrochaites y una de S. peruvianum, con alta resistencia al pasador N. elegantalis.
Del cruzamiento entre plantas de tomate cultivado y accesiones silvestres de S. habrochaites resistentes al pasador se obtuvieron seis poblaciones híbridas. Se produjeron semillas de cuatro poblaciones RC1 y cuatro poblaciones RC2, las cuales presentaron baja eficiencia en el cruzamiento y poca cantidad de semilla por fruto; igualmente, se obtuvieron semillas de tres poblaciones F2 y dos poblaciones F3 (Restrepo, Vallejo y Lobo, 2007).
El objetivo de esta investigación fue la evaluación de poblaciones segregantes producidas a partir de cruzamientos entre tomate cultivado cv. maravilla y la accesión silvestre PI134418 de S. habrochaites var. glabratum resistente al pasador del fruto.
El trabajo se realizó en el Centro Experimental de la Universidad Nacional sede Palmira. Se utilizó el diseño de bloques completos al azar con seis tratamientos y cinco repeticiones. Los tratamientos correspondieron a las poblaciones P1, P2, F1, F2, RC1 y RC2 (Tabla 1). La semilla de los parentales fueron cedidas por la Red de Recursos Genéticos y Biotecnología de Embrapa (Brasil), por el Centro de Recursos Genéticos de tomate de la Universidad de California (USA), por Corpoica La Selva (Colombia) y por el Grupo de Investigación de Hortalizas de la Universidad Nacional de Colombia Sede Palmira.

Cada parcela experimental estuvo conformada por ocho plantas. Las dos plantas de los extremos se utilizaron para eliminar el efecto de bordes, las seis plantas centrales correspondieron a la parcela útil. Para disminuir el efecto de bordes en cada uno de los lados de cada bloque se sembró un surco adicional con plaacute;ntulas de tomate del cv. maravilla. Con el fin de obtener un nivel suficiente de pasador en todas las parcelas experimentales se sembraron surcos intercalados con el mismo tomate cv. maravilla (muy susceptible al insecto). La distancia entre plantas fue de 0.60 m, entre surcos de 1.2 m y entre bloques de 2 m. Se realizó el manejo agronómico que se usa convencionalmente en cultivos comerciales de tomate, con la única diferencia de que una vez empezada la floración no se volvieron a aplicar insecticidas.
Se seleccionaron al azar ocho inflorescencias en cada una de las plantas de las seis poblaciones descritas (Tabla 1). A partir del anaacute;lisis de todos los frutos producidos por dichas inflorescencias se evaluaron por planta las variables: número promedio de frutos por racimo, peso promedio de fruto, formato del fruto, color del fruto maduro, frutos afectados por pasador (%), número promedio de orificios de entrada de pasador por fruto y densidad promedio de tricomas por fruto. La variable frutos afectados por pasador (%) se transformó usando arcoseno √x y la variable número promedio de orificios de entrada por fruto se transformó usando √x+0.5.
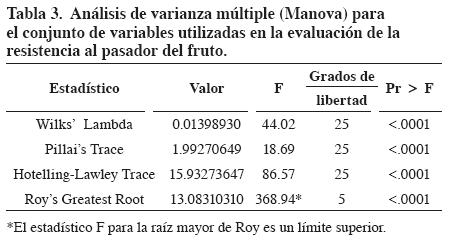
Se realizó un anaacute;lisis de varianza múltiple mediante el uso del procedimiento GLM Manova de SAS, con miras a determinar si había o no diferencias significativas entre poblaciones para el conjunto de variables cuantitativas evaluadas. Se hizo comparación de medias a través de la prueba de DMS (diferencias mínimas significativas).
Se hallaron las correlaciones simples de Pearson entre las variables número promedio de frutos por racimo y peso promedio de fruto; y entre peso promedio de fruto y densidad promedio de tricomas por fruto, a través del procedimiento Corr. de SAS. Se determinaron las correlaciones parciales entre la variable dependiente frutos afectados por pasador (%), y cada una de las siguientes variables independientes número promedio de frutos por racimo, peso promedio de fruto y densidad promedio de tricomas por fruto (procedimiento corr. partial de SAS).
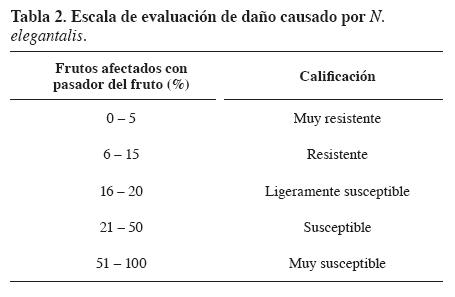
Se realizó un anaacute;lisis de regresión simple a través del procedimiento Reg. de SAS, entre la variable de respuesta frutos afectados por pasador (%) y la variable independiente peso promedio de fruto. De acuerdo con los frutos afectados con pasador (%) que presentó cada planta, éstas recibieron una calificación (de acuerdo con la escala de evaluación de daño propuesta en la Tabla 2).
Anaacute;lisis conjunto de las poblaciones
Se presentaron diferencias altamente significativas entre poblaciones para el conjunto de variables utilizadas en la evaluación de la resistencia al pasador (número promedio de frutos por racimo, peso promedio de fruto, frutos afectados por pasador (%), número promedio de orificios de entrada por fruto y densidad promedio de tricomas por fruto), que sugieren que existe al menos una población con un promedio del conjunto de descriptores que es significativamente diferente de las demaacute;s (Tabla 3).
La población P1 (padre muy resistente al pasador) presentó número promedio de frutos por racimo significativamente diferente del promedio de la población P2 (padre muy susceptible al pasador) (Tabla 4). La población F1 (híbrido interespecífico) registró promedio de frutos por racimo, que estaacute; por encima del rango comprendido entre los dos parentales que le dieron origen, y por consiguiente, se puede inferir que esto se debe posiblemente a la manifestación de vigor híbrido. Por otro lado, las poblaciones segregantes F2, RC1 y RC2, presentaron promedios similares de frutos por racimo, y significativamente diferentes de los mismos promedios de las poblaciones P2 y F1 (Tabla 4).
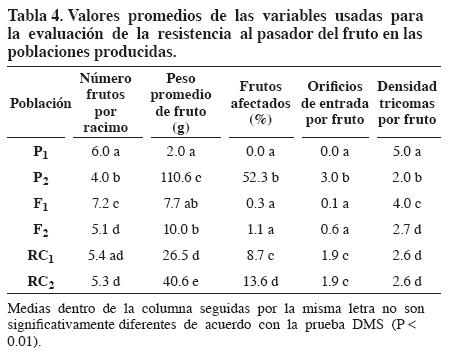
La población P1 registró peso promedio de fruto significativamente menor que el mismo promedio de la población P2 (Tabla 4). Las poblaciones F1 y F2 presentaron promedios de fruto por racimo, similares al promedio de la población P1 y significativamente diferentes del promedio de la población P2, y por tanto se puede inferir que posiblemente hay dominancia de los alelos que disminuyen la expresión de dicho caraacute;cter, es decir, que hay dominancia del tamaño pequeño de fruto sobre el tamaño grande del mismo (Tabla 4).
La población segregante RC1 presentó peso promedio de fruto mayor y significativamente diferente con respecto al promedio de la población F2, lo cual indica que el método del retrocruzamiento es maacute;s eficiente que el de la autofecundación, para contrarrestar la dominancia de los alelos que codifican por el tamaño pequeño del fruto (Tabla 4). Por otro lado, al comparar los pesos promedios de frutos por racimo de las poblaciones RC1 y RC2 se observó cómo este último fue mayor y significativamente diferente del primero, y por ende se pudo confirmar que al ir haciendo retrocruzamientos sucesivos hacia el padre recurrente (parental susceptible) es posible la recuperación paulatina del tamaño grande del fruto (Tabla 4).
El 52.3% de los frutos fueron afectados por pasador en la población P2; mientras que en la población P1 ningún fruto fue afectado por el insecto (Tabla 4). Las poblaciones F1 y F2 presentaron promedios de frutos afectados similares al promedio de la población P1. Por otro lado, la población RC2 registró promedio de frutos afectados significativamente mayor que el mismo promedio de la población RC1, de lo cual, se puede deducir que a medida que se hacen retrocruzamientos sucesivos hacia el padre recurrente (parental susceptible) la resistencia se va diluyendo (Tabla 4). El alto porcentaje de frutos afectados en la población P2 (padre susceptible) indicó que el nivel de infestación en condiciones de campo fue adecuado para evaluar la resistencia al pasador en las poblaciones evaluadas.
En la población P1 ningún fruto presentó orificios de entrada de larvas de pasador; mientras que en la población P2 se registró un promedio de tres orificios de entrada por fruto (Tabla 4). Las poblaciones F1 y F2 registraron promedios de orificios de entrada por fruto similares al promedio de la población P1. Por otro lado, las poblaciones RC1 y RC2 presentaron promedios de orificios de entrada similares, pero significativamente diferentes de los promedios de las otras poblaciones (Tabla 4).
La población P1 presentó densidad promedio de tricomas significativamente mayor que la densidad promedio de la población P2 (Tabla 4). La población F1 registró densidad promedio de tricomas que tiende a ser maacute;s parecida a la densidad de la población P1. Por otro lado, las poblaciones segregantes F2, RC1 y RC2 presentaron promedios de densidad de tricomas similares y significativamente diferentes de los mismos promedios de las poblaciones P1, P2 y F1 (Tabla 4).
La variable peso promedio de fruto (PPF) registró alta asociación negativa con la densidad promedio de tricomas por fruto (DTF) y sugirió que a mayor peso promedio de fruto ellos presentan menor densidad de tricomas (Tabla 5). Como se observa en la Tabla 4, la población P1 presentó el menor peso promedio de fruto y la maacute;s alta densidad promedio de tricomas por fruto; mientras que la población P2 registró el mayor peso promedio de fruto y la densidad de tricomas por fruto maacute;s baja. Por otro lado, se presentó alta asociación negativa entre las variables número promedio de frutos por racimo (NFR) y peso promedio de fruto (PPF), lo cual era de esperarse (Tabla 5).

Se presentó alta asociación positiva entre las variables frutos afectados por pasador (FAP) y peso promedio de fruto (PPF) (Tabla 6). Este resultado es similar al encontrado por Salinas, Vallejo y Estrada (1993) que reportaron correlación simple positiva (r = 0.4870; p = 0.01) entre el número de frutos dañados por pasador y el peso promedio de fruto. Por otro lado, no se presentó asociación entre la variable frutos afectados por pasador (FAP) y las variables número promedio de frutos por racimo (NFR) y densidad promedio de tricomas por fruto (DTF) (Tabla 6). De lo anterior se infiere que solo la variable peso promedio de fruto por racimo tuvo efecto importante sobre el ataque de los frutos por parte del insecto.

Al realizar el anaacute;lisis de regresión simple entre la variable independiente peso promedio de fruto (PPF) y la variable dependiente frutos afectados por pasador (FAP), se encontró que el modelo que establece la mejor relación entre las variables es el modelo de una línea recta, cuya ecuación fue:

La ecuación se cumple rigurosamente para valores de peso promedio de fruto superiores a 19.0 g e inferiores a 150 g. Un porcentaje muy bajo de frutos con peso promedio inferior a 19 g fueron ocasionalmente atacados por el insecto, y por ende se puede inferir que el insecto no tiene preferencia por frutos muy pequeños.
De la ecuación se deduce que por un incremento de un gramo en el peso promedio de fruto (a partir de frutos con un peso promedio de 19 g), el porcentaje de frutos afectados por pasador se incrementa en 0.47%. El valor del coeficiente de determinación indica que este modelo explica el 82.30% de la variabilidad observada en el porcentaje de frutos afectados por pasador por planta. El nivel de significancia calculado para esta variable fue menor al 0.0001, el cual es inferior al nivel de significancia crítico 0.01, y por ende la variable fue altamente significativa en el modelo.
Del anaacute;lisis conjunto de los resultados obtenidos al realizar las correlaciones parciales y la regresión simple se pudo inferir que los tricomas y el número promedio de frutos por racimo no ejercieron ningún efecto sobre el ataque de los frutos por parte del pasador; mientras que el peso promedio de fruto sí tuvo un efecto importante sobre dicho ataque, de tal manera que el insecto tiende a producir un mayor daño a medida que se incrementa el peso promedio de fruto.
El hecho de que los tricomas de la variedad glabratum de S. habrochaites no sean un factor que confiera resistencia al pasador del fruto N. elegantalis, es muy interesante, pues dicho factor al actuar como barrera física o como productor de aleloquímicos tipo 2-tridecanona y 2-undecanona es el que se ha encontrado asociado con la resistencia de varios insectos plaga de tomate diferentes a N. elegantalis (Williams et al., 1980; Zamir et al., 1984; Kennedy, Farrar y Kashyap, 1991; Eigenbrode et al., 1993; Barbosa y Maluf, 1996; Pocoví et al, 1998; y Aragao et al, 2000).
Análisis por población
Todas las plantas evaluadas de la población P1 (accesión PI134418 de la variedad glabratum de Shabrochaites) presentaron 0.0% de frutos afectados por pasador, y por ende fueron catalogadas como muy resistentes. Ademaacute;s, todas presentaron color del fruto maduro verde claro, formato de fruto ligeramente achatado y muy alta densidad de tricomas en los frutos (Tabla 7). Algunas plantas de esta población presentaron frutos que tenían posturas de pasador; sin embargo, no se presentó daño causado por el insecto.

Las plantas de la población P2 (cultivar maravilla de S. lycopersicum) presentaron entre 37.0% y 76.9% de frutos afectados por pasador, y fueron clasificadas como susceptibles o muy susceptibles al insecto; igualmente, presentaron frutos maduros de color rojo, formato de fruto tipo chonto y baja densidad de tricomas en los frutos (Tabla 7).
En la población F1 (híbrido interespecífico) solo dos plantas presentaron porcentajes muy bajos de frutos afectados (2.4% y 5.1%, respectivamente) y furon catalogadas como muy resistente y resistente respectivamente; el resto de plantas presentaron 0.0% de frutos afectados y se clasificaron como muy resistentes. Todas las plantas exhibieron frutos maduros de color amarillo claro, alta densidad de tricomas en frutos y formatos redondo o ligeramente achatado (Tabla 7).
En la población F2 solo tres plantas registraron porcentaje muy bajo de frutos afectados (2.0, 3.4 y 11.5%, respectivamente) y fueron catalogadas las dos primeras como muy resistentes y la última como resistente; las demaacute;s plantas presentaron 0.0% de frutos afectados y se clasificaron como muy resistentes. Las plantas exhibieron frutos maduros de colores diferentes (naranja, verde claro o diversas tonalidades de amarillo), densidades de tricomas en frutos (muy baja, baja, media o alta), y formatos de fruto redondo, ligeramente achatado o achatado (Tabla 7). Como se aprecia, la población F2 se comportó como la típica segregante para los caracteres color del fruto maduro, densidad de tricomas y formato de fruto; sin embargo, no se comportó de la misma manera para el caraacute;cter frutos afectados por pasador. Lo anterior se debió posiblemente a que los frutos de las plantas de esta población presentaron peso promedio muy bajo (10.0 g/planta), y por consiguiente no fueron atractivos para el insecto.
Las plantas de la población RC1 presentaron entre 0.0% y 40.9% de frutos afectados por pasador y fueron clasificadas como muy resistentes, resistentes, ligeramente susceptibles o susceptibles al insecto; igualmente, presentaron frutos maduros de diversos colores (rojo claro, naranja o diferentes tonalidades de amarillo), formato de fruto tipo (redondo, chonto o ligeramente achatado) y densidades de tricomas en los frutos baja, media o alta (Tabla 8). De los resultados se infiere que la población RC1 se comportó como la típica segregante para todos los caracteres evaluados.

Las plantas de la población RC2 presentaron entre 0.0% y 34.6% de frutos afectados por pasador y fueron clasificadas como muy resistentes, resistentes, ligeramente susceptibles o susceptibles al insecto; igualmente, presentaron frutos maduros de diversos colores (amarillo, naranja o diferentes tonalidades de rojo), formato de fruto tipo chonto o redondo, y densidades de tricomas en los frutos baja, media o alta (Tabla 9). Como se aprecia, la población RC2 también se comportó como la típica segregante para todos los caracteres evaluados.

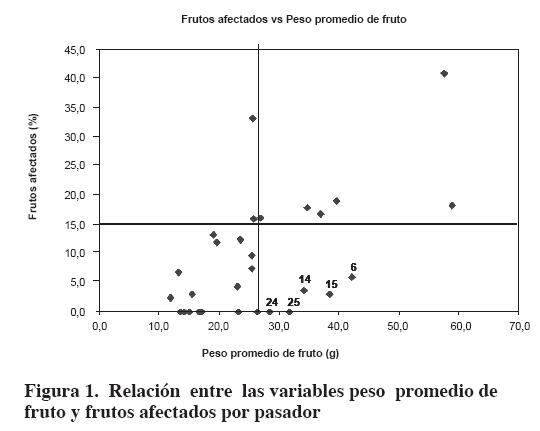
Al analizar la relación entre las variables peso promedio de fruto y frutos afectados por pasador en las plantas de la población RC1, se observa en la Figura 1 que es posible obtener plantas recombinantes con resistencia al pasador y pesos promedios de fruto superiores a la media de la población (plantas 6, 14, 15, 24 y 25). La mayoría de esos recombinantes presentaron baja densidad de tricomas y confirmaron que ese factor no es el responsable de la resistencia al pasador
Del anaacute;lisis de la relación entre las variables peso promedio de fruto y frutos afectados por pasador en las plantas de la población RC2, se observó que también es posible obtener plantas recombinantes con resistencia al pasador y pesos promedios de fruto superiores a la media de la población (plantas 1, 4, 8, 10, 15 y 22). La mayoría de esos recombinantes presentaron densidad baja o media de tricomas y confirmaron nuevamente que ese factor no es el responsable de la resistencia al pasador (Figura 2).
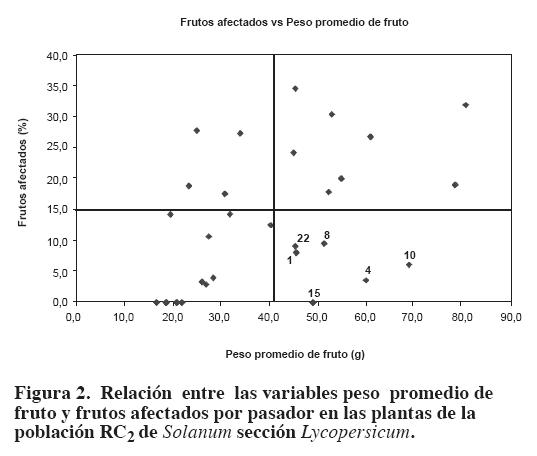
La identificación de recombinantes RC2 resistentes al pasador y con pesos promedios de fruto que fluctuaron entre 45.1g y 68.6 g, es muy valiosa, pues el insecto tiende a producir mayor daño a medida que se incrementa el peso promedio de fruto. De lo anterior se infiere que el método del retrocruzamiento fue efectivo en romper gradualmente la asociación negativa entre elpeso promedio de fruto y la resistencia al insecto, y por consiguiente es posible que al continuar realizando retrocruzamientos hacia el tomate chonto cultivado se pueda seguir incrementando paulatinamente el peso promedio de fruto y conservar la resistencia al pasador del fruto.
Al comparar las diversas poblaciones provenientes de autofecundación y retrocruzamiento se encontró dominancia de los alelos que disminuyen la expresión del caraacute;cter peso promedio de fruto.
El método del retrocruzamiento resultó maacute;s eficiente que el de la autofecundación para contrarrestar la dominancia de los alelos que codifican por el tamaño pequeño de fruto.
Los tricomas y el número promedio de frutos por racimo no ejercieron ningún efecto sobre el ataque de los frutos por parte del pasador; mientras que el peso promedio de fruto tuvo efecto importante sobre dicho ataque, de tal manera que el insecto tiende a producir un mayor daño en la medida en que se incrementa el peso promedio de fruto.
Las poblaciones RC1 y RC2 se comportaron como típicas segregantes para los caracteres evaluados y exhibieron amplia variabilidad en los mismos.
El método del retrocruzamiento permitió romper gradualmente la asociación negativa entre el peso promedio de fruto y la resistencia al insecto.
Se obtuvieron plantas recombinantes RC2 con resistencia al pasador y pesos promedios de fruto que fluctuaron entre 45.1g y 68.6 g. La mayoría de esos recombinantes presentaron baja densidad de tricomas y confirmaron que ese factor no fue el responsable de la resistencia al insecto.
El artículo se derivó de la tesis doctoral de E. F. Restrepo S. adelantada con recursos del Programa de Investigación Mejoramiento Genético, Agronomía y Producción de Semillas de Hortalizas de la sede Palmira, con apoyo de Colciencias y la Corporación Colombiana de Investigación Agropecuaria (Corpoica La Selva).
1. Aragao, C. A.; Maluf, W. R.; Dantas, B. F.; Gavilanes, M. L.; Cardoso, M. D. 2000. Tricomas foliares associados a resistencia ao acaro rajado (Tetranychus urticae Koch) em linhagens de tomateiro com alto teor de 2-tridecanona nos foliolos. Cienc. Agrotec. 24: 81-93 [ Links ]
2. Barbosa, L. V.; Maluf, W. R. 1996. Heritability of 2-tridecanone- mediated arthropod resistance in an interspecific segregating generation of tomato. Rev Bras Genet. 19 (3) : 465-468. [ Links ]
3. Eigenbrode, S. D.; Trumble, J. T. 1993. Antibiosis to bett armyworn (Spodoptera exigua) in Lycopersicon accesions. Hortic Sci. 28 (9): 932-934. [ Links ]
4. Eiras, A.; Blackmer, J. 2003. Time of eclosion and larval behavoir of the tomato fruit borer, N. elegantalis (Guenée) (Lepidóptera: Crambridae) on tomato, Lycopersicon esculentum. Sci Agric. 60 (1): 195 -197. [ Links ]
5. Kennedy, G.; Farrar, R.; Kashyap, R. 1991. 2-tridecanone glandular trichome mediated insect resistance in tomato. Effects on parasitoids and predators of Heliothis zea. American Chemical Society Symposium. No. 449. p. 150-165. [ Links ]
6. Luckwill, L. C. 1943. The genus lycopersicon : an historical, biological, and taxonomic survey of the wild and cultivated tomatoes. Aberdeen University Studies. 120: 1-44. [ Links ]
7. Marcano, R. V. 1991. Estudio de la biología y algunos aspectos del comportamiento del perforador del fruto del tomate Neoleucinodes elegantalis (Lepidóptera: Pyralidae) en tomate. Agron Trop (Maracay). 41 (5): 257-263. [ Links ]
8. Neves, L. G.; Leal, N. R.; Rodrigues, R.; Pereira, N. E. 2003. Estimativa de paraacute;metros genéticos e correlacao entre componentes de resistencia a traca do tomateiro em progenies de Lycopersicon esculentum x L. hirsutum f. glabratum. Hortic Bras. 21 (3): 458-461. [ Links ]
9. Pocoví, M.; Gilardón, E.; Gorustovich, M.; Olsen, A.; Gray, L.; Hernández, C.; Petrinich, C.; Collavino, G. 1998. 2-tridecanona y su asociación con la resistencia a la polilla del tomate (Tuta absoluta Meyrick) y a la arañuela roja (Tetranycus urticae Koch). Rev. Fac. Agron Univ Nac La Plata. 103 (2): 165-171. [ Links ]
10. Restrepo, E. F.; Vallejo, F. A.; Lobo, M. 2006. Evaluación de la resistencia al pasador del fruto Neoleucinodes elegantalis y caracterización morfoagronómica de germoplasma silvestre de Lycopersicon spp. Acta Agron. (Palmira). 55 (1): 15-21. [ Links ]
11. Restrepo, E. F.; Vallejo, F. A.; Lobo, M. 2007. Producción de poblaciones segregantes resistentes al pasador del fruto a partir de cruzamientos entre tomate y accesiones silvestres de Lycopersicon spp. Acta Agron. (Palmira). 56 (1): 1-6 . [ Links ]
12. Salinas, A. H.; Estrada, E. I.; Vallejo, F. A. 1993. Evaluación de la resistencia al pasador del fruto del tomate Neoleucinodes elegantalis (Guenée) en materiales de Lycopersicon hirsutum Hum y Bonpl y Lycopersicon pimpinellifolium (Just) Mill y su transferencia a materiales cultivados de tomate L. esculentum Mill. Acta Agron (Palmira) 43(1): 44-56. [ Links ]
13. Vallejo C.; Franco A. 1999. Mejoramiento genético y producción de tomate en Colombia. Palmira: Universidad Nacional de Colombia. 216 p. [ Links ]
14. Williams, W. G.; Kennedy, G. G.; Yamamoto, E. T.; Thacker, J. D.; Bordner, J. 1980. 2-tridecanone: a naturally occurring insecticide from the wild tomato Lycopersicon hirsutum f. glabratum. Science. 207: 888-889. [ Links ]
15. Zamir, D.; Ben-David, T.; Rudich, J.; Juvik, J. 1984. Frecuency distributions and linkage relationships of 2-tridecanone in interspecific segregrating generations in tomato. Euphytica. 33: 481-48. [ Links ]

