










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.57 no.2 Palmira Apr./June 2008
RESUMEN
Los objetivos del trabajo fueron determinar la época de maduración del polen y del estigma y establecer el tipo de polinización de la uchuva en condiciones de invernadero con cinco genotipos de uchuva. La investigación se realizó en la Granja Botana de la Universidad de Nariño (2.820 msnm, 13°C, precipitación de 800 mm/año y humedad relativa de 82%). P. peruviana tomó 37 días para la apertura floral, la cual se efectuó entre las 7:00 a.m. y las 10:00 a.m. En el 85% de las flores la primera antera fue dehiscente al día siguiente de la apertura floral. Los granos de polen tuvieron 97% de viabilidad a los 35 días. El polen maduró antes de la antesis y el estigma fue receptivo antes de la apertura de la flor. Tanto el polen como el pistilo maduraron a los 35 días de desarrollo, dos días antes de la apertura floral; la receptividad del pistilo se presentó dos días antes de la antesis, fenómeno que restringe la autopolinización. En invernadero, la ausencia de vectores influyó en la polinización de P. peruviana. En flores emasculadas y sometidas a libre polinización se presentó baja formación de frutos y semillas, existió respuesta diferencial a la polinización dentro del invernadero entre las muestras evaluadas. P. peruviana presentó polinización mixta con 54% de polinización cruzada.
Palabras claves: Physalis peruviana; antesis; apertura floral; autopolinización; polinización cruzada.
ABSTRACT
The objectives of the study were to determine the time of pollen and stigma ripening and to establish the kind of pollination in Physalis. This research was conducted in the Botana Farm at the University of Nariño, Colombia (2820 m a s l, 13 ° C, pluvial precipitation of 800 mm / year and relative humidity of 82%). P. peruviana took 37 days to anthesis, which took place between 7:00 and 10:00 am. In 85% of the flowers the first anther dehiscent was the day after the opening of the flower. The pollen grains showed 97% of viability to 35 days. Pollen matured before anthesis and stigma was receptive before the opening of the flower. Both, pollen and the pistil matured to 35 days of development, two days before anthesis; receptivity of the pistil was filed two days prior to anthesis, a phenomenon that restricts the self. The absence of vectors influenced pollination of P. peruviana. In emasculate flowers and subjected to self-pollination is introduced low fruit and seed formation, there was a differential response in pollination among samples inside the greenhouse. P. peruviana showed mixed pollination with 54% of cross-pollination.
Key words: Physalis peruviana; anthesis; self-pollination; cross-pollination.
Las flores de Physalis peruviana son pentámeras, hermafroditas, solitarias y pedunculadas, se ubican en las axilas de las ramas; la corola es glabra por dentro, con una línea de pelos por fuera y con bordes ciliados (Ibarra y Jurado, l989; Fisher, 2000).
Según el tipo de polinización las especies vegetales se distribuyen entre la autogamia y la alogamia, las especies de polinización intermedia se denominan a menudo alógamas o especies de reproducción mixta (Briggs y Knowles, 1967; Chávez, 1993; Poehlman y Sleper, 2003). Muchas especies tienen mecanismos que evitan o restringen la autopolinización, y la incompatibilidad y la androesterilidad son las más comunes. Existen muchas causas de incompatibilidad como la protoginia, la protandría y la heterostilia (Vallejo y Estrada, 2002).
En la uchuva prevalece la alogamia (Santana y Angarita, 1999; Fisher, 2000), pero son escasos los estudios relacionados con la fenología, fisiología y biología floral, los cuales son básicos en investigaciones de evolución, planificación de estrategias de conservación y en la escogencia de los métodos de mejoramiento genético. Las flores son fácilmente polinizadas por insectos y el viento, y se presenta también autopolinización (CRFG, 2003).
Con base en las consideraciones anteriores se emprendió el estudio para determinar la época de maduración del polen y el pistilo, y evaluar el grado de alogamia o autogamia en cinco genotipos de uchuva.
La investigación se llevó a cabo en la Granja Experimental Botana de la Universidad de Nariño, situada en el altiplano de Pasto a 2.820 msnm, 01°09'12" N y 77°18'31" W (Lagos et al., 2001), con cinco genotipos de la colección de la Universidad de Nariño (Tabla 1) cultivados en condiciones de invernadero. Se utilizaron 10 plantas por genotipo, sembradas en materas de 10 kg. Las observaciones de apertura floral y antesis se realizaron entre las 07:00 y las 18:00 horas. Durante la evaluación se presentó una temperatura mensual mínima de 9.9°C y máxima de 16°C, humedad relativa de 84.22% (IDEAM, 2005).
Veinticuatro horas antes de la apertura se marcaron 20 botones florales y cada media hora se registraron los cambios en las diferentes partes de la flor hasta la caída de los pétalos. En la apertura de la flor se registró la hora de iniciación, duración entre el comienzo y la máxima apertura y la hora del cierre de los pétalos. Se evaluaron los días al cierre permanente de los pétalos, caída de los mismos, hora de la antesis, inicio de la antesis a partir de la apertura floral, fin de la antesis y el tiempo transcurrido entre la antesis de la primera y la última antera.
La maduración del grano de polen se monitoreó a los 20, 30, 35 y 40 días de desarrollo en botones florales de 2 mm de longitud; para medir la viabilidad polínica (VP) se utilizó la tinción con acetocarmín de 2 %.
Para evaluar la viabilidad del polen se marcaron 20 primordios florales por cada estado de maduración, se polinizaron artificialmente diez flores emasculadas, se protegieron con bolsas de glacyne durante cinco días y se observó el cuajado del fruto. Se seleccionaron los estados que presentaron valores de VP mayores al 87% (30, 35 y 40 días). A partir de los 30 días se recolectó polen de 30, 32, 35, 37 y 39 días de desarrollo, se hicieron polinizaciones artificiales en flores emasculadas y se las comparó con el testigo de polinización natural. Se evaluó el número de frutos formados y el número de semillas por fruto (NSF).
La receptividad de los pistilos a los 20, 30, 35 y 40 días se evaluó utilizando como indicador la formación de burbujas al adicionar una gota de peróxido de hidrógeno en los estigmas.
Para confirmar la receptividad del pistilo se marcaron 20 primordios florales para cada estado de maduración, se polinizaron artificialmente diez flores emasculadas y se protegieron con bolsas de glacyne. La fertilidad de los óvulos y la receptividad de los pistilos se estimó por el número de frutos y semillas por fruto (NSF).
En cada genotipo se evaluaron tres formas de polinización: autopolinización (AUTO, 50 botones florales), polinización natural o libre polinización (LP, 20 botones florales) y libre polinización de flores emasculadas (EMAS, 50 flores). En los frutos maduros se determinó el peso (PS) y número de semillas (NS) para estimar el porcentaje de polinización cruzada natural y autocompatibilidad. Con el fin de determinar la existencia de mecanismos que favorecen la autogamia se emascularon 20 botones antes de la apertura floral.
La madurez del polen, receptividad del pistilo y los mecanismos de polinización se analizaron mediante el diseño completamente al azar, en donde cada estructura floral marcada se consideró como una repetición. Se utilizó la prueba de diferencia mínima significativa para comparación de medias.
Morfología de las flores de P. peruviana
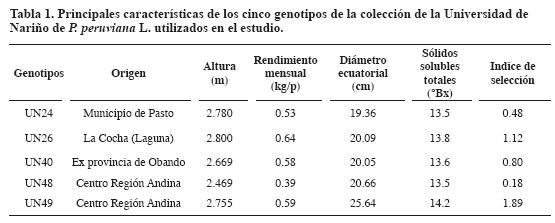
La Figura 1a muestra la estructura de una flor típica; la corola (Figura 1b) es gamopétala, generalmente rotácea con simetría radiada, regular o actinomorfa, amarilla y con manchas en la parte intermedia de los pétalos. El cáliz (Figura 1c) es gamosépalo, verde oscuro, con abundante pubescencia y venas moradas; durante los primeros 40 a 45 días del desarrollo es verde, pero con la maduración toma color dorado (Fisher, 2000). El ovario (Figura 1d) es súpero, amarillo verdoso a verde. El estilo es morado con estigma amarillo verdoso. Cuando la flor ha sido fecundada el estigma se torna café oscuro a medida que se desarrolla. Las anteras (Figura 1e y Figura 1f) son oblongas, biloculadas con dehiscencia lateral y los filamentos glabros y adnados a la base de la corola. La antera es verde oscura a morada, aunque en los primeros días es amarilla cremosa.
La unión del filamento con la antera es basifija. El pistilo es más largo que el androceo (Figura 1f), indicador de incompatibilidad heteromórfica conocida como heterostilia (Fehr, 1987; Borojevic, 1990).
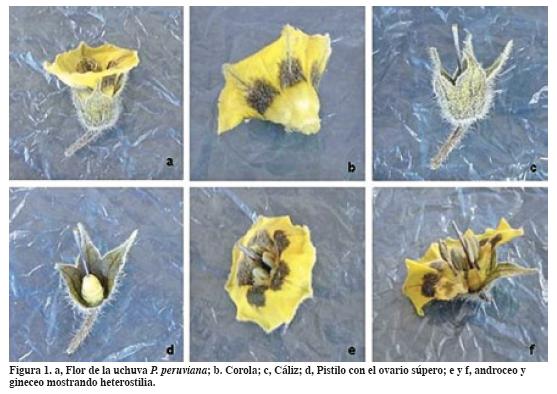
En la Tabla 2 se reportan los tamaños para las partes de una flor de P. peruviana. En muy baja frecuencia se encontraron flores atípicas que presentaron más de un pistilo y flores con seis estambres. Esta variación obedece a fallas en la división celular y a la fusión de los carpelos (Stevenson y Mertens, 1986). Una flor presenta varios carpelos. Cada carpelo puede convertirse en un pistilo simple, de modo que la flor posee varios pistilos; sin embargo, casi siempre los carpelos se funden en diversos grados para formar un solo pistilo compuesto.
Inicio de apertura floral (IAF)
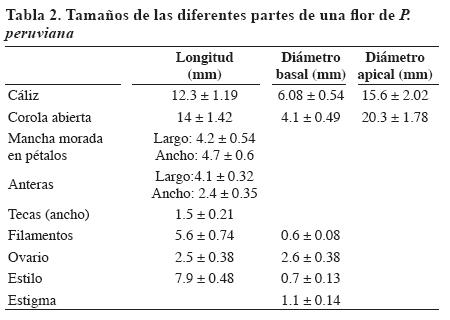
El 98% de las flores iniciaron apertura entre las 07:00 y las 10:00 horas, el 2% lo hizo al día siguiente en el mismo horario (Tabla 3). La duración de la apertura (DA) osciló entre 2.5 hasta 5 horas después del IAF, las flores que iniciaron su apertura a las 7:00 a.m. completaron el proceso entre 3 a 4.5 horas. La menor DA correspondiente a 2.5 horas se dio en las flores que tuvieron un IAF entre 9:30 y 10:00 a.m. y las que presentaron una DA de 5 horas fueron las que iniciaron la apertura a las 8:00 a.m. Esta DA puede estar influenciada por la temperatura, ya que las flores que presentaron DA menor están en un horario en que la temperatura fue superior a las de las primeras horas de la mañana.
En el mismo día el cierre parcial de pétalos (CPP) se presentó entre las 16:00 y 18:00 horas mostrando en la parte apical una forma de estrella. El inicio del CPP se registró en 40% a las 16:00, el 10% a las 17:00 y un 40% a las 18:00 horas. Durante los cuatro a seis días siguientes los pétalos se abrieron en la mañana y se cerraron en la tarde. El cierre final de pétalos (CFP) ocurrió entre los cinco a siete días después de la apertura floral. El CFP fue del 55%, 25% y 20% a los cinco, seis y cuatro días, en su orden. Este comportamiento también se encontró en la India, donde el CFP se presentó entre los tres a cuatro días después de la apertura floral (Fischer, 2000). A los cinco (15%), seis (20%) y siete días (65%) después de la apertura floral los pétalos se desprenden. Con respecto al CFP, 45% de los pétalos cayeron al día siguiente y 55% a los dos días.
Antesis.
La línea de ruptura para la liberación de los granos de polen se localizó a lo largo de la antera entre los dos lóculos de cada teca. Esta apertura es de tipo longitudinal (Afah,1974). En el 56.87% de las flores evaluadas ocurrió la antesis entre las 10:00 y las 13:00 horas. Para las primeras horas de la mañana (8:00 a 9:00) se registró el 23.53% de flores en antesis. Después de las 14:00 horas se observó significativa disminución de la antesis, presentándose el menor porcentaje desde las 14:00 horas (11.76%).
Estos registros coinciden con lo descrito por Ordóñez y Ruano (2002), quienes encontraron que las flores liberan el polen con mayor facilidad entre las 12:00 y las 16:00 horas, periodo donde se registra la mayor temperatura del día. Mosquera (2002) encontró que en Botana la antesis se llevó a cabo entre las 9:00 y las 16:00 horas.
Las horas de inicio de la antesis desde el IAF presentaron variación alta, la menor en el rango de 2 a 5 horas con 15% en el primer día de apertura floral y el 85% ocurrió al día siguiente hasta los tres días después del IAF. El inicio de la antesis fue más frecuente entre los 22 y 29 días después de la apertura floral con 60%, temperaturas de 10.4°C y 17.4°C y ausencia de lluvias. Para los otros días de evaluación se registraron lloviznas con cambios en la temperatura, retardando la apertura en la primera antera en 72 horas después de la apertura floral, lo cual fue observado en el 5% de las flores evaluadas. Mosquera (2002) encontró en el primer día de antesis efectiva temperatura cercana a los 16 °C, y por debajo de los 15 °C se limitó.
El final de la antesis (FA) ocurrió al segundo día después de la apertura floral con 25%, y en mayor porcentaje al tercer día (55%) y 20% al cuarto día, cuando la dehiscencia llegó a la parte basal de la última antera. El tiempo transcurrido entre la antesis de la primera y la última antera fue de uno a tres días. El 75% de las anteras tomó dos días (46 a 51 horas), el 10% tardó un día (12 a 17 horas), el 10% día y medio (25 a 30 horas) y el 5% más de tres días (75 horas).
La diferencia marcada entre el inicio y el final de la antesis puede darse por la continua variación de los factores climáticos y por la restricción que pueden tener las flores a la autopolinización, pues éstas se hacen receptivas a los 33 días de desarrollo aún antes del IAF, y la dehiscencia de las anteras se presentó en 85% a partir de los 38 días de desarrollo, un día después de la apertura floral.
Maduración del polen y viabilidad polínica (VP) en P. peruviana.
La VP se incrementó desde los 20 hasta los 35 días. Los granos de polen de anteras de 35 días (dos días pre-antesis) y 40 días (tres días post-antesis) presentaron los mayores porcentajes de VP (Tabla 4). La edad de 20 días fue diferente estadísticamente de los demás tratamientos. Los resultados permiten enunciar que a partir de los 30 días se dispone de 87.56% de polen apto para la polinización y fecundación para programas de fitomejoramiento.
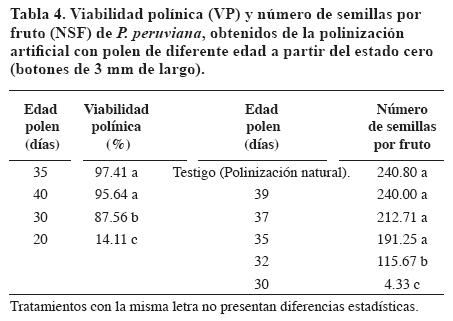
El número de semillas por fruto (NSF) (Tabla 4) se incrementó con la edad del polen. Los estados de desarrollo 35, 37, 39 días y el testigo no mostraron diferencias estadísticas entre ellos y fueron similares a las 207 - 267 semillas/fruto reportadas por Hejeile e Ibarra (2001), Ordóñez y Ruano (2002) en condiciones de Botana. El estado de 30 días presentó el más bajo valor, probablemente por la falta de maduración del polen.
Pueden considerarse como maduros los granos de polen de 35 días, que corresponden a dos días antes de la antesis. En este estado los granos de polen se encuentran en una solución nutritiva de consistencia gelatinosa que a medida que transcurre la apertura floral se vuelve menos adherente y adquieren consistencia polvorienta en el momento de la antesis.
Con polinizaciones adecuadas, es decir, con granos de polen maduros que presentaron una VP superior al 95% se obtuvieron frutos con un diámetro ecuatorial de 15 a 20 mm, un eje polar de 14.5 a 19.3 mm y un NSF de 145 a 302. Esto sucede en otras solanáceas como tomate (Solanum lycopersicum), en donde el número de óvulos fecundados afectó el crecimiento del fruto (Mosquera, 2002).
Madurez del pistilo
Los pistilos de 20, 30, 35 y 40 días reaccionaron positivamente en 100% a la prueba del peróxido de hidrógeno, indicando probablemente que la receptividad del ovario se presenta antes de la antesis y permite la polinización por otras flores.
El NSF se asoció con la edad del ovario hasta los 35 días. La prueba de comparación de medias (Tabla 5) para el NSF indicó que las polinizaciones de flores emasculadas a los 35 y 37 días y el testigo no presentaron diferencias significativas. El estado de 33 días mostró diferencias significativas con 30 días. Estos resultados permiten establecer que a partir de los 33 días el pistilo está receptivo y se puede polinizar.
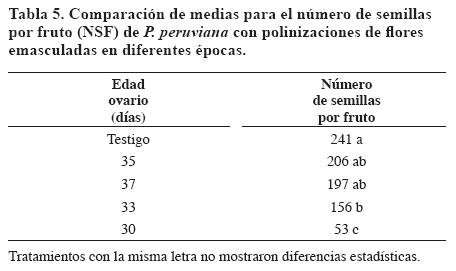
La receptividad del pistilo se presenta dos días antes de la dehiscencia de las anteras y la apertura de la flor. A partir de los 33 hasta los 37 días se obtuvieron frutos con DEF de 12.7 a 20.1 mm, EP de 12.3 mm a 19.6 mm y NSF de 76 a 336.
La flor de P. peruviana restringe la autopolinización, ya que el pistilo se hace receptivo antes que los estambres entraran en antesis. Los estambres se abrieron después de la apertura floral en forma alterna y en 85% al día siguiente y permitieron que otro polen la fecunde.
Mecanismos de polinización.
El mayor porcentaje de frutos formados ocurrió con la autofecundación (AUTO), seguido de libre polinización (LP) y de la libre polinización de flores previamente emasculadas (EMAS) (Tabla 6). En el caso de emasculación antes de apertura floral (AA) no hubo formación de frutos que indicaran ausencia de mecanismos que aseguran la autogamia. Frankel y Galun (1977) mencionan la falta de vectores de polinización como insectos, la falta de movimiento de las ramas, flores y polen debido a la no presencia de lluvias y a la baja frecuencia de corrientes de aire como causas de la baja eficiencia de la polinización obtenida en invernadero en LP y EMAS.
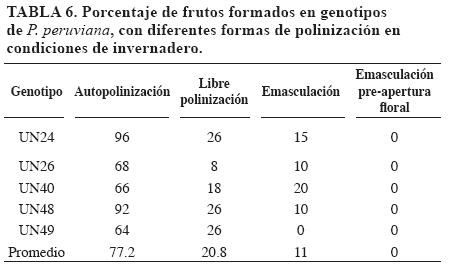
En AUTO y LP se obtuvieron mayor porcentaje de frutos que en EMAS, que carece de anteras. En AUTO el estigma de una flor protegida estuvo rodeado por un ambiente con mayor cantidad de polen que en LP y EMAS, además el estigma quedó sometido al poco polen que le llegó del ambiente.
Los genotipos UN24 y UN48 fueron los más prolíficos; UN26 fue el de peor desempeño. Dentro de LP el máximo porcentaje de frutos obtenidos fue de 26 para los genotipos UN24, UN48 y UN49. En EMAS UN49 no formó frutos y el máximo porcentaje fue para UN40 (20%).
Es probable que este comportamiento diferencial obedezca a la interacción de mecanismos genéticos y el ambiente limitante para la polinización que ofrece el invernadero. Puede ser que en estas condiciones los genotipos de baja formación de fruto en AUTO, tales como UN26, UN40 y UN49 manifiesten fenómenos de autoincompatibilidad. Se estimó el porcentaje de polinización cruzada natural en 52.88%.
Según el número de semillas por fruto (NSF) el genotipo UN48 fue estadísticamente diferente del resto de genotipos. Los mejores fueron UN24 y UN48 y los peores UN49 y UN40 (Tabla 7). El análisis de varianza permitió establecer que la interacción genotipos por FP no fue significativa.
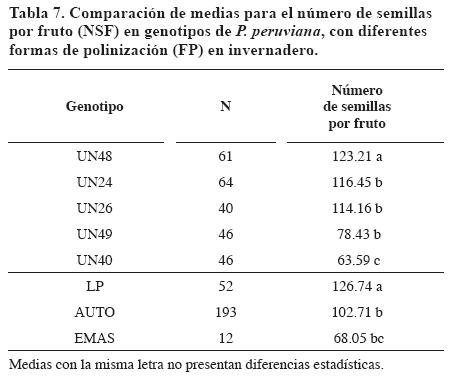
Las formas de polinización presentaron diferencias estadísticas. La LP (NSF=126.74) mostró diferencias estadísticas con las demás. Las razones expuestas en la formación de frutos en invernadero también son válidas para explicar el bajo NSF obtenido por el EMAS: La polinización cruzada natural estimada con base en la relación del NSF del EMAS y la LP fue de 53.69%. Con base en el alto promedio de número de frutos y semillas obtenidos en AUTO también se puede concluir que no existieron fenómenos de autoincompatibilidad genética.
Por las evidencias registradas se puede considerar a P. peruviana como una especie de polinización mixta, que se debe a fallas en el reemplazo de la exogamia por un mecanismo definido de autogamia (Frankel y Galun, 1977). El fenómeno se presenta en aquellas especies que han evolucionado de la alogamia hacia la autogamia.
En el cultivar regional de P. peruviana la apertura de las flores ocurrió a los 37 días (primordios de tres milímetros) y se llevó a cabo entre las 07:00 y las 10:30 horas. La apertura floral duró entre 5 a 7 días. La antesis se llevó a cabo el día siguiente de la apertura floral y duró tres días.
Los granos de polen alcanzaron la madurez dos días antes de la antesis, la receptividad estigmática también se presentó a los 35 días de desarrollo.
La receptividad del pistilo se presentó aproximadamente dos días antes de la dehiscencia de las anteras y la apertura de la flor. Los estambres se abrieron en forma alterna.
En P. peruviana la polinización cruzada natural fue del 53% y 54% en condiciones de invernadero.
Los autores expresan sus agradecimientos a la Universidad de Nariño por los recursos aportados para la financiación de la investigación doctoral de T. C. Lagos B., de la cual se derivó este artículo.
1.Afah, N. 1974. Plant anatomy. 2nd ed. New York: Pergamon Press. p. 425-500. [ Links ]
2. Borojevic, S. 1990. Principles and methods of plant breeding. Amsterdam: Elsevier. 368p. [ Links ]
3.Briggs, F.N.; knowles, P.F. 1967. Introduction to plant breeding. New Cork: Reinhold Publishing. 426p. [ Links ]
4.Chávez, J. L. 1993. Mejoramiento de plantas 2a ed. México: Trillas - Universidad Autonoma Agraria Antonio Narro. 163p. [ Links ]
5.CRFG. California Rare Fruit Growers. 2003. Cape gooseberry: Physalis peruviana L. http://www.ctfg.org/pubs/ff/cape-gooseberrv.html 10/09/2003. [ Links ]
6.Fehr, W. 1987. Principles of cultivar development: theory and technique. New York: Iowa University-Macmillan. Vol 1 536p. [ Links ]
7.Fischer, G. 2000. Crecimiento y desarrollo. En: Flórez, V. J., Fisher, G.y Sora ángel, D. (Eds). Producción, poscosecha y exportación de la uchuva Physalis peruviana L. Bogotá, Universidad Nacional de Colombia, Ministerio de Agricultura y Desarrollo Rural y Asociación Hortifrutícola de Colombia. 175p. [ Links ]
8.Frankel, R.; Galun, E. 1977. Pollination mechanism, reproduction and plant breeding. Berlin: Springer-Verlag. 281p. [ Links ]
9.Hejeile, H.; Ibarra, A. 2001. Colección y caracterización de recursos genéticos de uviIIa (Physalis peruviana L.) en algunos municipios del sur del departamento de Nariño. Trabajo de Grado (Ing Agr). Pasto: Universidad de Nariño, Facultad de Ciencias Agrícolas. 123p. [ Links ]
10.Ibarra, V.; Jurado, H. 1989. Evaluación de diferentes distancias de siembra en el cultivo de la uvilla (Physalis peruviana L.). Trabajo de Grado (Ing Agr). Pasto: Universidad de Nariño, Facultad de Ciencias Agrícolas. 79 p. [ Links ]
11.IDEAM. Instituto de Hidrologia, Meteorología y Estudios Ambientales. 2005. Datos meteorológicos. Pasto, Estación Meteorológica Botana. [ Links ]
12.Lagos, T. C.; Criollo, H.; Mosquera, C. 2001. Evaluación preliminar de cultivares de uvilla o uchuva (Physalis peruviana L.) para escoger materiales con base en la calidad del fruto. Rev. Ciencias Agric (Colombia). 18(2): 82-94. [ Links ]
13.Mosquera, C. 2002. Polinización entomófila de la uvilla (Physalis peruviana L). Rev Ciencias Agríc. 19(1/2):140-1 56. [ Links ]
14.Ordóñez, M.; Ruano, L. 2002. Evaluación de diferentes horarios de polinización artificial en uvilla (Physalis peruviana L.) bajo condiciones de campo de la granja de Botana. Trabajo de Grado (Ing. Agr.). Pasto: Universidad de Nariño, Facultad de Ciencias Agrícolas. 85p. [ Links ]
15.Poehlman, J.M.; Sleper, D.A. 2003. Mejoramiento genético de las cosechas. 2. ed. Trad. M. Guzmán y M. A. Hernández. México: Limusa, 511p. [ Links ]
16.Santana, G.; Angarita, A. 1999. Regeneración adventicia de somaclones de uchuva. http://www.agronomia.unal.edu.co/biblioteca/public/revista/artic/Fuchuv.htm [ Links ]
17.Stevenson, F.; Mertens, T. 1986. Anatomía vegetal. México: Limusa. 169-182 pp. [ Links ]
18.Vallejo, F. A.; Estrada E. I. 2002. Mejoramiento genético de plantas. Palmira: Universidad Nacional de Colombia. 401p. [ Links ]

