










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.59 no.1 Palmira Jan./Mar. 2010
1Department of Soil Science and Agriculture Chemistry, Institute of Agricultural Science, BHU, Varanasi, UP. amitavabhu@gmail.com 2Department of Agricultural and Food Engineering, Indian Institute of Technology Kharagpur 721 302, India pbsb@iitkgp.ac.in
The effect of vesicular arbuscular mycorrhizal (VAM) infection on growth and P nutrition in Maize (Zea mays, cv.DDH hybrid) were assessed in Oxisol pot experiment. Maize was grown inoculated with spores of VAM fungi Glomus mosseae or non-inoculated. Low soluble ferrous phosphate (FePO4.4H2O) was added to the mycorrhized and non-micrrohized maized. The dry weight of mycorrhized plants with added phosphate (P) were higher than in mycorrhized plants without added P or non-mycorrhized plants with added P. The amount of P in the soil samples from pots with mycorrhizal plants fertilized with P was evidently smaller than those in samples also fertilized non-mycorrhizal plants. The percentage of P was higher in tissues of fertilized mycorrhial plants than in those mycorrihzed plants without or nonmycorrhized plants with added low-soluble P. These results indicated that plants in VAM symbiosis mobilize P better from low-soluble P than non-mycorrhized plants.
Key words: Mycoryhyzae, micorrizas arbusculares, Glomus mosseae, phosphorus, maize, Oxisol.
En condiciones de casa de malla se evaluó el efecto de la infestación con micorrizas vesículo arbusculares (VAM) en la asimilación de P por plantas de maíz (Zea mays, cv.DDH hybrid) cultivadas en un Oxisol. Como micorriza se utilizó el hongo Glomus mosseae. En ambos tratamientos (con micorriza y sin ella) se aplicó fosfato ferroso (FePO4.4H2O). La producción de MS de maíz fue mayor cuando se aplicaron el hongo + la fuente de P. La cantidad de P en el suelo con este tratamiento fue menor que en el suelo fertilizado pero sin aplicación del hongo. En las hojas de las plantas las mayores concentraciones de P se observaron igualmente en el tratamiento micorriza + aplicación de fertilizante.
Palabra clave: Mycoryhyzae, micorrizas arbusculares, Glomus mosseae, fósforo, maiz, Oxisol.
Most of the soils that are now available in East Midnapore districts of West Bengal are acidic and low in fertility. These soils are classified as Oxisols and occupy about 10% of the land area of West Bengal (Anonymous, 2005). One of the factors that allow plants to grow on this poor soils is their association with vesicular arbuscular-mycorrhizal (VAM) that enable uptake of immobile phosphorus and other nutrients (Nielsen and Jensen, 1983). Plants in the VAM symbiosis are better supplied with P than non-mycorrhizal plants. VAM fungi increase growth of plants in soil where P deficiency limits the growth of non-mycorrhizal plants. Perhaps VAM fungi synthesize and secrete organic substances (siderophores) which increase the desoption of P in labile soil P pools. With organic acids, VAM fungi can solubilize low-soluble and insoluble P sources which is a part of the crystalline structure of minerals in the soil (Anonymous, (2005). Citrate exudation into the rhizosphere is thought to increase the availability of P to plants by mobilizing sparingly soluble Fe and Al phosphates or Ca-P. It has been demonstrated that some dicotyledonous plant roots, and specially non-mycorrhizal plants such as Lupinus albus, but not L. angustifolia, and Brassica napus are capable of releasing large amounts of organic acids into the rhizosphere in response to P deficiency. Synergism of VAM fungi with soil bacteria such as Pseudomonas and Agrobacterium, fungi that will secrete organic acids and solubilize lowsoluble P. In VAM symbiosis in the rhizosphere of plants more of these bacteria and fungi can be found (Mosse, 1973).
Some authors have observed that plants in VAM symbiosis were better supplied with P from low-soluble P sources or rock phosphate. Leopold and Hoefner (Bowen et al., 1975) reported that VAM fungi were using rock phosphate and thus improved the shoot dry weight of clover. The same was observed by Antunus and Cardoso (Owusu-Bennoah and Wild, 1980) in citrus plants. One the other hand, Sreenivasa et al. (Bagyaraj, 1984) observed that VAM fungi did not mobilize more P from rock phosphate by the chilli plant. It seems that the mobilization of P from lowsoluble P sources depends on the species of VAM fungi and sybiotic plants.
The objective of this study was to investigate and explore the hypothesis that the VAM fungi G. mosseae improved the mobilization of P from low-soluble FePO4. 4H2O as well as the shoot growth of maize.
Spores of VAM fungi were isolated from the soil of a corn fi eld. They were concentrated in this soil and in maize roots accordingly to the method of Ferguson and Woodhead (8). The crude substrate from the soil, pieces of maize roots infected with VAM fungi and concentrated spores VAM fungi were used as the inoculum for plants in pot experiments. Spores of VAM fungi Glomus mosseae and Glomus multicaulis predominate in this substrate.
In pot experiments, maize (Zea mys L. cv. DDH Hybrid) was grown in sterilized soil with low P fertility (P2O5 = 6.2 mg/100 g soil) in a greenhouse for 92 days. The soil was autoclaved twice for two hours at 123 °C at 1.3 atmospheres (Antunes and Cardoso, 1991). Three plant were grown in each pot.
In May, sterilized maize seeds were sown in 7.5 kg of sterile soil in pots. The maize seeds were sterilized for 15 minutes with 33% hydrogen peroxide (H2O5) (Sreenivasa et al., 1993). Plant were grown as non-mycorrhized plants, non-mycorrhized plants with 1.8 g/pot of added FePO4.4H2O, mycorrhized plants, non-mycorrhized plant with added FePO4.4H2O. Mycorrhizal plants were inoculated with VAM mycorrhizal fungi accordingly to the procedure described by Nielsen and Jensen (Nielsen and Jensen,1983). Maize seeds were sown above the layer of the inoculum, 5 cm below the soil surface. At the end of the growing season, all plants were taken from the pots. The percentage of infection in their roots was evaluated after dying with Trypan blue (Krishna and Bagyaraj, 1982) by estimating the percent root colonization. Per cent root infection was obtained as follows:

The dry weights of whole plants, root length and fruits were measured. Soil solution P in the bulk soil was determined using Adams (1974) procedure. Plants with roots were dug out and they were shaken carefully to avoid a damage to the roots. After removing the bulk soil from the roots they were put into the cylinders with the adhering soil and soil solution (CLi) was collected in a similar manner as for the bulk soil. Phosphorus concentration was determined by colorimetry (Nielsen and Jensen, 1983). The level of P in plant tissues was also analyzed (Phillips and Hayman, 1970).
To estimate total hyphae length modified gridline intersect method was employed. Soil cores of known volume were collected at random, thoroughly mixed, sub samples (10 g) removed and suspended in water and passed through a sieve with 250 µM openings. The filtrate is blended for 15 seg and a portion (25 mL) passed through a membrane with pores < 5 µm diam. The membrane is briefl y flooded with a tryptan blue solution (0.5 g of tryptan blue + 500 mL of deionised water + 170 mL of Lactic acid + 330 mL of glycerine) and rinsed with deionised water. The membrane cut to fit on a microscope slide and observed at 100X through an eyepiece whipple disc that has a 10 by 10 lined intersect grid. The total length of the hyphae was estimated by gridline intersect method using the following equation:
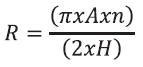
where, R = hyphal length, A total area in which roots are distributed, n = no of intersections between roots and scribed lines, and H = total length of scribed lines.
Hyphae were counted only if showing characterstics described as typical for AM fungal hypahe, such as dichotomous branching, abundant angular projections, and absence of septa.
The root of VAM symbotic plants showed greater infection when a low-soluble P source was added (Figure 1). Perhaps this was the result of synergism of VAM fungi with some bacteria such as Pesudomonas and Agrobacterium and other fungi which can be solubilize P in the soil with organic acids. The rhizosphere of VAM plants often contains many of these bacteria and fungi. These microorganisms also sythesize organic substances, such as hormones and vitamins, which stimulate the growth of VAM dungi (Mosse, 1973. Vejsadova et al. (1993) also reported that P-solubilizing bacteria increased the percentage of root infection with VAM fungi. The stimulation of growth of VAM fungi with the added ion (Fe) might also be possible.
Total length of hyphae in the pot soil did vary substantially (Figure 1). The amount of total hyphal length in soil AM plants was seven times higher than in the non-mycorrhizal plants. The relationship between total hyphae length in the soil and root collonization indicates positive correlation.
The shoot and fruit dry weights of nonmycorrhizal maize with added FePO4. 4H2O was greater than in non-mycorrhizal maize without the added FePO4. 4H2O (Figure 2) These results indicated that non-mycorrhizal plants were also able to use low-soluble P to some extent. In all mycorrhizal plants, the dry weight was higher than in the non-mycorrhizal, indicating that VAM fungi improved the mineral nutrition of maize.
The root length is an important parameter which influences nutrient uptake as the nutrient absorbing root surface area depends up on the length of the roots (Figure 1). Root length is more for non mycorrhizal plants with added insoluble phosphorus source followed by mycorrhizal plants with added insoluble P source, mycorrhizal plants and non mycorrhizal plants.
Soil solution P from pots with mycorrhizal plants (V+: 0.65 µmole/L) contained significantly lower amounts of P than those with non-mycorrhizal plants(C+: 0.79 µmole/ L) indicating that plants in VAM symbiosis absorbed more P from the soil. Low-soluble P which was added to the soil resulted in a higher level of P in this soil. But in the soil from pots with fertilized VAM plants, the amount of P(V+: 2.04 µmole/L) was clearly smaller than in the soil of non-mycorrhizal plants fertilized with the same P source(C+: 2.36 µmole/L). Plants in VAM symbiosis better mobilized P from low-soluble P.
The percentage of P in the fruits of fertilized VAM plants was significantly higher than in VAM plants without added P and in nonmycorrhizal plants with added P (Figure2), showing that plants in VAM sybiosis better use P from low-soluble P than non-mycorrhizal plants.
The efficiency with which each plant root segment absorbs P is given by the uptake per unit length and per unit time, the influx. The P influx is shown in Figure 2. At harvest, the influx is more for mycorrhizal plants with added insoluble P followed by mycorrhizal plants, non mycorrhizal maize and non mycorrhizal maize with added in soluble source of P. It means that VAM fungi are more capable in enhancing plant growth and nutrient uptake.
Adams, F. 1974. Soil solution. In: The Plant Root and its Environment. Carson E.W. (ed.). University of Virginia Press Charlottesville VA p. 441 - 481. [ Links ]
Anonymous. 2005. Department of Agriculture & Co-operation: Statistics at a Glance, Government of West Bengal, Writers Building, Kolkata http://www.wbgov.nic.in [ Links ]
Antunes, V. and Cardoso, E. J. 1991. Growth and nutrient status of citrus plants as influenced by mycorrhiza and phosphorus application. Plant and Soil 131(1):11-20. [ Links ]
Bagyaraj, D. J. 1984. Biological interactions with mycorrhizal fungi. Powell, D. J. Bagyaraj (eds.). ii. Interactions between VAM and benefi cial soil organisms. VA Mycorrhiza. CRC Press, Inc., Boca Raton, FL. p . 135 -136. [ Links ]
Bowen, G. D.; Bevege, D. I.; and Mosse, B. 1975. Phosphate phydiology of vesicular-arbuscular mycorrhizas,. In: Sanders, F. E.; Mosse, B.; and Tinker, P. B. (eds.). Endomycorrhizas. Academic Press, London, United Kingdom. p. 241-243 [ Links ]
Ferguson, J. J. and Woodhead, S. H. 1984. Production of Endomycorrhizal Inoculum. A Increase and Maintenance of Vesicular-arbuscular Mycorrhizal Fungi. In.: Schenck, N. C. (ed.). Methods and principles of mycorrhizal research. The American Phytopathological Society, St. Paul, MN. p. 47 - 48. [ Links ]
Krishna, K. R. and Bagyaraj, D. J. 1982. Effect of vesicular-arbuscular mycorriha and soluble phosphate on Abelmoscus esculentus (L.) Moensch. Plant and Soil 64:209-213. [ Links ]
Leopold, H. J. and Hoefner, W. 2002. Improvement of clover yield and quality by inoculation with VAM fungi and Rhizobium bacteria. Angewandte Botanik 56(1-2):23-34. [ Links ]
Mosse, B. 1973. Advances in the study of vesicular- abrucular mycorrhiza Ann. Rev. Phytopathol. 11:171 - 196. [ Links ]
Nielsen, J. I.and Jensen, A. 1983. Influence of vesicular -arbuscular mycorrhiza fungi on growth and uptake of various nutrients as well as uptake ratio of fertilizer P for lucerne (Medicago sativa). Plant and Soil 70:165-172. [ Links ]
Owusu-Bennoah, E. and Wild, A. 1980. Effects of vesicular-arbuscular mycorrhiza on the size of the labile pool of soil phosphate. Plant and Soil 54:233-240. [ Links ]
Phillips, J. M. and Hayman, D. S. 1970. Improved procedures for clearing rotts and staining parastic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55:15 & 161. [ Links ]
Sreenivasa, M. N.; Krishnaray, P. U.; Gangadhara, G. M.; and Manjunathaiah, H. M. 1993. Response of chilli (Capsicum annuum L.) to the inoculation of an efficient vesicular-arbuscular mycorrhizal fungus. Sci. Hortic. 53(1-2):45- 52. [ Links ]

