










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.59 no.4 Palmira Oct./Dec. 2010
1Ingeniera Agroforestal. Universidad de Nariño, Pasto, Colombia. 2Facultad de Ciencias Agrícolas. Universidad de Nariño, Pasto, Colombia. Autores para correspondencia: tclagosb@udenar.edu.co, nancyta.p@hotmail.com, kathesantacruz@hotmail.com, hectoramiro@hotmail.com
Se realizó un análisis del comportamiento meiótico de las especies de lulo S. hirtum, S. quitoense y S. sessiliflorum, siguiendo la metodología convencional para los estudios de microsporogénesis. Se tomaron botones florales en diferentes estados de desarrollo, fijándolos por 24 horas en una solución de tres partes de etanol por una parte de ácido acético, saturada con trazas de cristales de cloruro férrico. Para la preparación de las placas se siguió la técnica de aplastamiento, se liberaron las células madres del grano de polen y finalmente se hicieron las observaciones bajo microscopía de luz. El análisis mostró que la meiosis se presenta en longitudes de antera que van desde los 2,79 mm hasta los 4,45 mm. La normalidad meiótica fue del 100%, tanto para meiosis I, como para la meiosis II. El índice meiótico en las tres especies fue del 99,98% lo cual indica que son buenos parentales y que pueden utilizarse en programas de cruzamiento. Las tres especies evaluadas tienen igual número de cromosomas (2n=2X=24). La frecuencia de anormalidades durante el proceso meiótico fue baja para S. hirtum, y alta para S. quitoense; sin embargo, la viabilidad polínica fue de gran magnitud (91,2-97,3%).
Palabra clave: Meiosis, variabilidad, microsporogénesis, índice meiótico, normalidad meiótica.
An analysis of meiotic behavior of lulo species S. hirtum, S. quitoense and S. sessiliflorum, following the conventional methodology for studies of microsporogenesis was realized. Flower buds were taken at different stages of development, fixing them for 24 hours in a solution of three parts of ethanol per one part of acetic acid, saturated with traces of ferric chloride crystals. For the preparation of the slides following the technique of squash, releasing pollen mother cells and finally made the observations under light microscopy. The analysis showed that meiosis occurs in anther ranging from 2.79 to 4.45 mm. Meiotic normality was 100% for both meiosis I and II. The meiotic index in all three species was 99,98% indicating that they are good parental for use in crossbreeding programs. The three species have the same number of chromosomes (2n = 2x = 24). The frequency of abnormalities during the meiotic process was lower for S. hirtum, and high for S. quitoense, however, the pollen viability was of great magnitude (91.2-97.3%).
Key words: Meiosis, variability, microsporogenesis, index meiotic, meiotic normality.
Aunque el lulo es considerado como una especie promisoria desde hace setenta años por ser Colombia parte del centro de origen, este cultivo no ha alcanzado el grado de desarrollo esperado, debido a la dificultad de obtener cultivares mejorados con características deseables que puedan competir en el mercado. Además, los esfuerzos de la investigación en fitomejoramiento se han canalizado en cultivos tradicionales relacionados con seguridad alimentaria y a cultivos industriales o generadores de divisas (Bernal et al., 1998).
El lulo es una solanaceae cuyo centro primario de diversidad y variabilidad genética está ubicado entre Colombia, Ecuador y Perú. Al contrario del lulo de Ecuador, el de Colombia presenta espinas y se conoce como S. quitoense Lam. var. septentrionale (Dennis et al., 1985). S. quitoense es una especie diploide con un número cromosómico somático de 2n=2x=24 y un apareamiento cromosómico normal en meiosis (Vivar y Pinchinat, 1970).
Por otro lado, se acepta que el lulo está en proceso de domesticación, de ahí que al cambiar su hábitat natural y cultivarlo a plena exposición solar, se ha observado que reduce su periodo vegetativo y presenta un gran número de problemas fitosanitarios, los cuales han limitado su desarrollo como cultivo, especialmente en aquellas zonas donde la temperatura y la altura no son óptimas (Corpoica, 2004).
A pesar de la importancia de la especie como un cultivo potencial, las investigaciones en torno a su mejoramiento son escasas. Por lo tanto, es necesario aprovechar los recursos genéticos de una manera adecuada. En todo programa de mejoramiento genético, ya sea para la selección de genotipos élite o para la reproducción de híbridos, es necesario conocer la base genética y la estabilidad meiótica del germoplasma disponible (Huertas et al., 1996).
En ese sentido la citogenética, definida como una ciencia de investigación (citología + genética), que tiene por objeto el estudio de los aspectos genéticos observables en la célula mediante la microscopía, Caetano (2003), puede contribuir a un conocimiento más amplio de las colecciones utilizadas para el mejoramiento de las especies vegetales. De esta manera, la caracterización y el establecimiento de las propiedades de los genotipos, desde su estructura y comportamiento meiótico y mitótico, permiten no solo utilizar mejor los recursos genéticos en programas de mejoramiento, sino que también llevan a aclarar una serie de problemas evolutivos, tanto generales como referidos al origen de determinados grupos (Talledo y Escobar, 1994).
Desde la perspectiva genética, la meiosis puede considerarse como una diferenciación del núcleo, que tiene como metas producir células germinativas o sexuales (gametos), con un número haploide de cromosomas y nuevas combinaciones genconotípicas, dos características de vital importancia en la existencia de la diploidía y en los procesos evolutivos de la reproducción sexual en los eucariotos (Singh, 1993; Goicoechea y Giraldes, 1996).
En Colombia no se han realizado trabajos de citogenética de las especies de lulo. El estudio del comportamiento meiótico es muy importante para establecer el grado de homología cromosómica entre especies y definir algunos cruzamientos de tipo interespecífico, especialmente con algunas especies que poseen genes de utilidad para el lulo cultivado. Igualmente, el índice meiótico es una variable clave para definir los parentales que podrían hacer parte de un esquema de cruzamientos, sin tener que gastar recursos en la siembra y establecimiento del cultivo para probar si este es apto o no para producir suficiente semilla híbrida (Lagos et al., 2001).
Este trabajo se orientó a obtener conocimiento acerca de la citogenética de algunas especies del género Solanum, la cual es importante para resolver problemas de hibridación, reproducción y macho esterilidad y por supuesto, una herramienta útil en la caracterización de estos recursos genéticos.
Este trabajo se realizó en el Laboratorio de Cultivo de Tejidos de la Universidad de Nariño, Sede Pasto, ubicado a una altura de 2.488 msnm, 1°14'3" LN, 77°17'7" L O, con una temperatura promedio de 18° C y humedad relativa de 75%. Las especies de lulo evaluadas fueron S. hirtum Vahl., S. sessiliflorum Dunal, S. quitoense Lam., las cuales se encuentran propagadas en el Invernadero de Investigación de la Facultad de Ciencias Agrícolas de la Universidad de Nariño.
Para evaluar el comportamiento meiótico se siguió la metodología convencional para los estudios de microsporogénesis (meiosis masculina) descrita por Belling (1926) y McClintock (1929) y adaptada por Dempsey (1993). Para ello se tomaron entre las 9 a 11 am, botones florales en diferentes estados de desarrollo. Estos se fijaron por 24 horas en una solución fresca (solución Farmer) de tres partes de etanol por una de ácido acético, saturada con trazas de cristales de cloruro férrico. El fijador se cambió, y el material se guardó bajo refrigeración.
Para la preparación de las placas se siguió la técnica de aplastamiento (squash), que consistió en remover las anteras de los botones florales, en portaobjetos con dos gotas de acetocarmin al 1%. Estas se cortaron con bisturí para liberar las células madres del grano de polen (CMPs). El material fue cubierto con una laminilla. El conjunto se presionó suavemente entre dos hojas de papel filtro y finalmente se hicieron las observaciones bajo microscopía de luz (Caetano, 2003).
Se determinó además el índice meiótico que corresponde al porcentaje de tétradas de microsporas viables. En S. sessiliflorum, se analizaron 43 placas, mientras que en S. hirtum y S. quitoense, se estudiaron 73 placas en cada una de ellas. En cada placa se observaron cien campos visuales. Se registraron las longitudes de anteras en milímetros y se realizaron los respectivos conteos de células en meiosis y microsporas.
Mediante microscopía de luz, en cada una de las especies se evaluaron todas las fases meióticas. También se determinó la normalidad meiótica (NM), la cual corresponde a la relación de células normales en cada fase sobre el total del número de células observadas (células normales + células anormales). Se consideran como células anormales aquellas que presentan ascensiones precoces, células retardadas y microcitos, entre otros. Además, se observaron el número de tétradas de microsporas viables para calcular el índice meiótico y la viabilidad del polen (VP) mediante la tinción con acetocarmín al 2%. Los granos de polen viables son aquellos que se tiñen de rojo (Dempsey, 1993).
En el Cuadro 1 se presenta la relación entre la longitud de antera versus el estadio de desarrollo meiótico. Se observa que en la clase dos (2,7-4,4 mm) se encuentra el mayor número de células en meiosis, mientras que en las clases cuatro y cinco (6,09-9,4 mm) se presentaron granos de polen viable. Esto indica que longitudes de antera por encima de 6,1 mm contienen microsporocitos que han terminado la meiosis y entrado en maduración y formación de grano polen. Entre los tamaños 1,14 y 6,098 mm se observan las diferentes fases de la meiosis, mostrando una relación entre estas clases de longitud de antera y la actividad meiótica de las CMPS (microsporocitos). Después que las anteras han alcanzado una longitud mayor a 6,1 mm, la actividad de la meiosis es nula y las tétradas liberan los granos de polen maduros.
En las clases dos y tres con longitudes de antera que oscilan entre 2,79 y 6,10 mm se encuentra la mayor actividad meiótica caracterizada por una frecuencia más elevada de células en diferentes estados de meiosis, lo que se traduce en un mayor número de granos de polen formados (Cuadro 1).
El Cuadro 2 describe el número de células halladas en cada fase de la meiosis de las tres especies de lulo evaluadas. En S. hirtum se encontraron un total de 4519, seguido por S. sessiliflorum con 1484 células y S. quitoense con 734 células. La profase I presentó un mayor número de células, debido a que es la fase más larga y compleja, conformada por las etapas de leptoteno, cigoteno, paquiteno, diploteno y diacinesis (Gardner et al., 2000).
La normalidad meiótica del 100% expresada por S. hirtum, S. quitoense y S. sessiliflorum (Cuadro 2), les permite formar un gran número de gametos masculinos que admiten la polinización y fecundación que garantizan la supervivencia de la especie en la naturaleza, sobresaliendo la especie S. hirtum. No se presentaron anormalidades como ascensiones precoces de cromosomas y cromosomas retardados, como las que encontró Lagos (2006) en Physalis peruviana y P. philadelphica. La Meiosis I microsporogenica mostrada por S. hirtum, comprende la profase I con sus etapas de cigoteno (Figura 1a), diploteno (Figura 1b), diacinesis (Figura 1c), la metafase I (Figura 1d), la anafase I (Figura 1e) y la telofase I, donde existe una reorganización de cromosomas en los polos para continuar con la Meiosis II (Figura 1f).
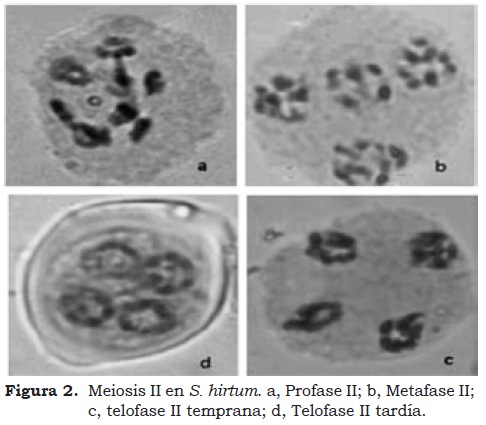
En la Figura 2 se presentan las diferentes fases de la Meiosis II, las cuales corresponden a profase II (Figura 2a); metafase II (Figura 2b); telofase II temprana (Figura 2c) y telofase II tardía. Como en la Meiosis I la normalidad en las diferentes fases de la Meiosis II fue del 100%. (Cuadro 2), los resultados encontrados tanto en la Meiosis I como en la Meiosis II de S. hirtum, S. quitoense y S. sessiliflorum son corroborados por los altos valores de IM (98,68 – 99,17%), NM (100%) y VP (91,98 – 97,33%).
En las Dicotiledóneas, las dos citocinesis ocurren inmediatamente al final de la telofase II (citocinesis simultánea), formando una tétrada de microsporas dentro de la pared de calosa (Figura 2d). Generalmente, el resultado consiste en un arreglo tetraédrico (Da Silva et al, 2001). Por otro lado, en las Monocotiledóneas se dan dos citocinesis sucesivas, la primera al final de la meiosis I y la segunda al final de la meiosis II, produciendo una tétrada isobilateral (Figura 3b).
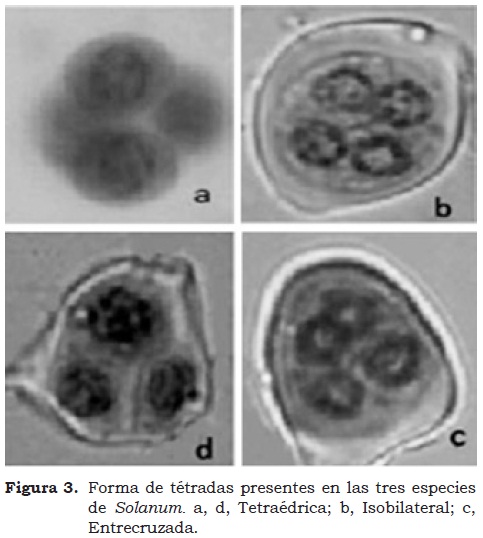
Por consiguiente, la forma de la tétrada es un indicador bastante confiable del tipo de citocinesis, a menos que otra reorientación haya tenido lugar en una fase más tardía. Como una regla, cada especie presenta un arreglo típico y único de tétradas de microsporas, sugiriendo un control genético para la característica (Da Silva et al, 2001 y Lagos, 2006). De igual manera, se encontraron otros tipos de tétrada como tetraédrica (Figura 3a, Figura 3d) y entrecruzada (Figura 3c).
El IM es importante determinarlo para saber si la planta puede ser un buen parental en un programa de cruzamientos. Desde el punto de vista genético, mientras más alto sea el porcentaje de tétradas viables, mejor será el genotipo evaluado (Caetano, 2004). Estos datos demuestran el buen comportamiento de todos los genotipos evaluados, los cuales no presentan barreras de cruzabilidad y son una fuente de variabilidad genética, y progenitores potenciales para incluir en programas de cruzamientos (Lagos et al., 2001).
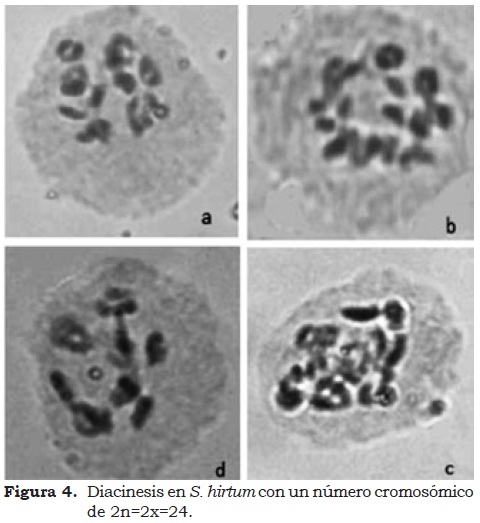
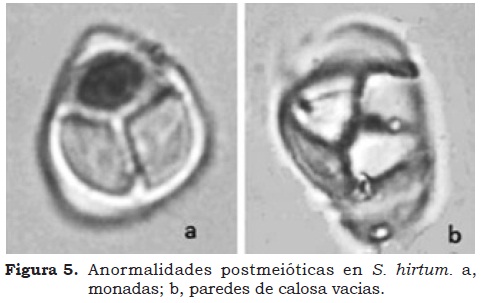
Las tres especies evaluadas presentaron igual número cromosómico (NC). De 6761 células evaluadas, el 100% correspondió a un NC 2n=2x=24 (Figura 4). La VP se relaciona directamente con la normalidad de la microsporogénesis (comprendiendo la pre-meiosis, la meiosis y la post-meiosis). La frecuencia de anormalidades se registro durante la post-meiosis (Figura 5a) con formación de monadas por fusión de microsporocitos. También existió un degeneramiento cromatínico y citoplasmático, que según Olaya (2002) produce unidades de calosa vacías (Figura 5b).
Lagos (2006) encontró resultados similares en su estudio sobre el comportamiento citogenético de la uchuva. Las fases de la Meiosis I y II, tanto en P. peruviana como en P. philadelphica, fueron normales, presentándose en escasa frecuencia, fallas en la segregación cromosómica causada por las ascensiones precoces en las Meiosis I y II y cromosomas retardados en Anafase II, siendo de principal importancia las primeras. La mayor parte de la asincronía en la migración de los cromosomas no tuvo repercusiones en el desarrollo normal de las tétradas. En cuanto a la viabilidad polínica y el índice meiótico fueron altos en las dos especies. En todas las accesiones de P. peruviana se presentaron más de un número cromosómico (NC). Los NC más característicos fueron 2n=24, 2n=36, 2n=48 y 2n=32.
El Cuadro 2 muestra los porcentajes de VP, siendo S. hirtum el que presenta el porcentaje mas alto 97,33%, seguido de S. quitoense con un 93,7% y S. sessiliflorum con el más bajo, 91,9%. Estos porcentajes permiten establecer que las especies presentan alto grado de adaptación a las condiciones ambientales del altiplano de Pasto. Con respecto al rango de adaptación, la especie crece en zonas altas desde Chile hasta Venezuela como planta silvestre y semisilvestre, entre los 1.500 y los 3.000 msnm (Fischer, 2000 y Zapata et al., 2002).
La viabilidad polínica fue alta en todas las especies evaluadas. Además, presentaron igual número cromosómico (2n=2x=24). Las fases que componen la meiosis I y II, en S. hirtum, S. quitoense y S. sessiliflorum, fueron normales.
El Índice Meiótico para las tres especies fue alto. Las tres especies pueden considerarse como buenos parentales, ya que son fuente de variabilidad genética que pueden incluirse en programas de cruzamientos.
Al Grupo de Investigación en Producción de Frutales Andinos de la Facultad de Ciencias Agrícolas de la Universidad de Nariño.
Belling, J. 1926. The iron acetocarmine method of fixing and staining chromosomes. Bull – Bull. 50: 160 – 162. [ Links ]
Bernal, J.; Lobo, M.; Londoño, M. 1998. Documento de presentación del material "Lulo La Selva". Corpoica. Rionegro, junio de1998. 77p. [ Links ]
Caetano, C. M. 2003. La aplicabilidad de la citogenética en Zea mays L.: genes mutantes meióticos. Revista de Ciencia Agrícolas Vol. XX. No I – II San Juan de Pasto 27 – 49p. [ Links ]
Caetano, C. M. 2004. Citogenética: notas de clase. Palmira, Universidad Nacional de Colombia, sede Palmira. [ Links ]
Corporación Colombiana de Investigación Agropecuaria-Corpoica. 2004. Inventario de bancos de germoplasma. C.I. La Selva. Rionegro, Antioquia. Base de datos en medio magnético. [ Links ]
Dempsey, E. 1993. Tradicional análisis of maizepachytene chromosomes. In: Freeling M. and Walbot V. (eds.). the maize Handbook , New York, Springer-Verlag New York Inc., pp. 432-441. [ Links ]
Dennis, F. G.; Herner, R.C.; Camacho, S. 1985. Naranjilla a potencial cash crop for the small farmer in latinoamerica. Acta Horticultural 158:475-481. [ Links ]
Da Silva, N.; Caetano, M.C.; Pagliarini, M. S. 2001. Cytogenetic characterization of Pedilanthus tithymaloides (L.) Poit (Euphorbiaceae). The Nucleus. 44(1, 2): 7 -12. [ Links ]
Fisher, G. 2000. Crecimiento y desarrollo. En: Flores, V., Fisher,G y Sora, A (Editores). Producción, post cosecha y explotación de la uchuva (Physalis peruviana L.). Bogotá, Fondo Nacional de fomento hortofrutícola, Ministerio de Agricultura y Desarrollo Rural. 175p. [ Links ]
Gardner, J.; Simmons, M.; Snustad, P. 2000. Principios de genética. 4a. Ed. Mexico, Limusa Wiley. 649p. [ Links ]
Goicoechea, P.; Giraldes, R. 1996. Plant chromosomes at meiosis. In Fukui and Nakayama Shigeki (Editors). Plant chromosomes laboratory methods. Boca Raton, Crc 274p. [ Links ]
Huertas, C.; Salazar, F.; Varón, F. 1996. Manejo integrado del cultivo del lulo (Solanum quitoense Lam.) en el Valle del Cauca. Boletín técnico. Palmira. ICA Mayo 22p. [ Links ]
Lagos, T.C.; Criollo, H.; Mosquera, C. 2001. Evaluación preliminar de cultivares de uvilla o uchuva (Physalis peruviana L.) para escoger materiales con base en la calidad del fruto. Revista de Ciencias Agrícolas (Colombia). 18(2): 82-94. [ Links ]
Lagos, T. 2006. Comportamiento citogenético de algunos genotipos de Physalis peruviana L. Tesis de doctorado en Ciencias Agropecuarias. Palmira, Universidad Nacional de Colombia. 92p. [ Links ]
McClintock, B. 1929. A method for making acetocarmin smears permanent. Stain Technol. 4:53 – 56. [ Links ]
Olaya, C. A. 2002. Primer estudio de la meiosis en Passiflora tripartita var. mollissima (Kunth) Hola-nielsen & Jorgensen, Passiflora tarminiana Coppens & Barney, Passiflora mixta L. y tres de sus híbridos. Tesis Ing. Agr. Manizales, Universidad de Caldas. 21 p. [ Links ]
Singh, R. J. 1993. Plant cytogenetics. Boca Raton, CRC. 391p. [ Links ]
Talledo, D.; Escobar, C. 1994. Análisis cariotípico comparativo de nueve especies del genero Solanum L. Biotempo: 11 -18. [ Links ]
Vivar, H. E.; Pinchinat, A. M. 1970. Viability of seed from interspecific crosses with naranjilla Solanum quitoense. Crop Science. 10: 450-451. [ Links ]
Zapata, J.L.; Saldarriaga, a.; Londoño, M.; Díaz, C. 2002. Manejo del cultivo de la uchuva en Colombia. Boletín técnico 14. Rionegro, Antioquia, Corpoica, Pronatta 40p. [ Links ]

