










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.60 no.4 Palmira Oct./Dec. 2011
1Facultad de Ciencias Agrícolas. Universidad de Córdoba. A.A 354. Montería - Córdoba, Colombia.
2The University of Texas at Austin. USA.
3Instituto de Investigación de Recursos Biológicos Alexander von Humboldt - CIAT. Palmira, Valle del Cauca, Colombia.
*Autor para correspondencia: hriveraj@gmail.com, hernandorivera@sinu.unicordoba.edu.co; †andresalvarez864@hotmail.com; ‡jdpalacio@utexas.edu; ††genyovana@gmail.com; ‡‡dianalopez430@gmail.com
Conocer la variabilidad genética del ñame, Dioscorea spp., permite apoyar estrategias de mejoramiento y conservación de este recurso fitogenético. El objetivo de este estudio fue la caracterización molecular de 20 accesiones de Dioscorea spp. mediante la técnica molecular de AFLP para determinar cómo se distribuye la variabilidad genética de manera intra e inter-específica. Los datos fueron analizados mediante los métodos de agrupación de correspondencia múltiple y análisis de similaridad de Dice, estableciendo los niveles de confiabilidad de los grupos genéticos mediante remuestreos. En términos de diversidad interespecífica, los valores promedios de similitud variaron entre 41.81% entre D. alata L. y D. rotundata Poir., y 33.51% entre D. trifida L.f. y D. esculenta (Lour.) Burkill, lo que sugiere alta diversidad genética entre las accesiones estudiadas, que formaron cuatro grupos genéticos: D. alata, D. rotundata, D. esculenta y D. trifida, confirmando correspondencia entre la caracterización morfológica, clasificación botánica y la caracterización molecular. En términos de diversidad intraespecífica para la especie D. alata, el análisis también reveló una composición heterogénea en la región Caribe colombiana. Estos estudios ayudarán a definir una estrategia adecuada para fines de conservación y apoyar los esfuerzos futuros en los programas de mejoramiento genético.
Palabra clave: Características agronómicas, caracterización molecular, Dioscorea spp., marcadores genéticos, recursos fitogenéticos, región Caribe de Colombia.
Knowing the genetic variability of yams, Dioscorea spp., is a good tool to support development and conservation strategies of this plant as genetic resource. The aim of this study was to carried out the molecular characterization of 20 accessions of Dioscorea spp. using the AFLP molecular technique to determine how genetic variation is distributed intra-and inter-specifically. Using multiple correspondence analysis and level of reliability of the genetic groups by resampling, the results showed high genetic variability among the accessions studied giving as a result four genetic groups: D. alata L., D. rotundata Poir., D. esculenta (Lour.) Burkill and D. trifida L.f., which confirmed a correspondence between the morphological and molecular characterization. The average values of similarity ranged from 41.81% in D. alata and D. rotundata, and 33.51% in D. trifida and D. esculenta. These data are consistent with previous morphological characterizations and systematics of the species in relation to their botanical sections. The analysis also revealed the heterogeneous composition of D. alata in the colombian Caribbean region; these studies will help to define an appropriate strategy for conservation to support future efforts in breeding programs.
Key words: Agronomy characters, colombian Caribbean region, Dioscorea spp., genetic marker, molecular characterization, plant genetic resources.
En muchos lugares del trópico, el ñame, Dioscorea spp., es un alimento de gran importancia, principalmente en áfrica occidental, parte del Asia sub-oriental, India, algunas zonas de Brasil y otros países de América tropical (Tamiru, 2006; Lebot et ál., 1998). El ñame muestra una amplia diversidad genética, tanto a nivel inter como intra-específico (Martin y Rhodes 1977; Okoli, 1991). La diversidad del ñame silvestre se ve aumentada por la domesticación del cultivo en varios países (Mignouna y Dansi, 2003). No obstante, el grado de diversidad genética de muchas especies de Dioscorea y sus relaciones genéticas aún no han sido totalmente investigados. Los intentos de caracterizar el ñame utilizando caracteres morfológicos (Hamon y Toure, 1990b;), isoenzimas (Hamon y Toure, 1990a; Dansi et ál., 2000) y marcadores moleculares no han dado resultados concluyentes debido al alto grado de variabilidad de este cultivo (Tamiru et ál., 2007).
Algunas de las más importantes variaciones morfológicas pueden ser el resultado de las diferencias en sólo unos pocos genes (Bradley et ál., 1997), observaciones que han sido apoyadas por estudios que muestran la ocurrencia de un alto grado de diversidad genética, tanto en especies de ñame de origen silvestre como cultivadas del sur de Etiopía y especies cultivadas en áfrica occidental, como D. alata L., D. bulbifera L., D cayenensis L. y D. rotundata Poir. (Tamiru et ál., 2007). Sin embargo, estudios de diversidad genética en esta especie realizados por Bustamante et ál. (2003) muestran que el género Dioscorea presenta alta similaridad entre las accesiones caracterizadas de Colombia. Para estudios taxonómicos, filogenéticos y de diversidad genética en ñame se han utilizado marcadores moleculares, entre ellos, la técnica de polimorfismo de longitud del fragmento amplificado (AFLP) (Vos et ál., 1995), (Ramser et ál., 1997; Mignouna et ál., 1998; Dansi et ál., 2000), lo cual ha permitido la detección de diferencias entre las variedades que se consideran similares con base en marcadores morfológicos y de isoenzimas, demostrando su utilidad como herramienta para discriminar introducciones en Dioscorea spp. (Dansi et ál., 2000) y permitiendo, inclusive, la detección de duplicados en las colecciones de germoplasma (Mignouna et ál., 2003).
El objetivo principal de este estudio fue el análisis de la diversidad genética intra e interespecífica de la colección de accesiones de ñame de la Universidad de Córdoba, Colombia, mediante la técnica de marcadores AFLP.
Material vegetal. Para el estudio se recolectó tejido foliar de 20 accesiones existentes en la colección de accesiones de ñame de la Universidad de Córdoba (Cuadro 1), provenientes de diferentes regiones de los departamentos de Córdoba, Sucre y Bolívar (Colombia). El tejido foliar se almacenó en bolsas plásticas con 50 g de sílica-gel, suficientes para mantener en condiciones favorables hasta 5 g de tejido. Posteriormente el tejido se maceró con nitrógeno líquido y se conservó a -196 °C.
Extracción de ADN. El componente molecular se obtuvo en el laboratorio de biología molecular del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Para la extracción del ADN se tomó tejido foliar de las diferentes accesiones previamente maceradas y conservadas con nitrógeno líquido. Para la extracción de ADN se utilizó el kit comercial de Qiagen® modificado mediante la reducción de la temperatura de incubación a 60 °C y tiempo de centrifugado a 5 min. La calidad del ADN se visualizó mediante electroforesis en gel de agarosa al 1% teñido con bromuro de etidio y visualizados en luz UV; la cuantificación se hizo mediante el uso del marcador de peso de ADN Lambda 20-bp.
Marcadores AFLP y amplificación por PCR. El ADN aislado de cada una de las accesiones se sometió a digestión con las enzimas de restricción EcoRI y MseI, posteriormente fueron adicionados dos adaptadores a los fragmentos de ADN generados en el proceso de restricción y ligados mediante la acción de la enzima T4 ligasa de ADN suplida por el kit comercial de Invitrogen.® El proceso de amplificación mediante el uso de PCR (Polymerase Chain Reaction, por sus siglas en inglés) se hizo siguiendo las instrucciones del kit comercial de Invitrogen.® Se utilizaron cebadores complementarios a la secuencia de los adaptadores y se hizo una preamplificación +1/+1 con nucleótidos adicionales. Las condiciones de amplificación se efectuaron en un termociclador PTC-100tm MJ Research Inc., a partir de los fragmentos generados en la reacción de digestión de cada una de las muestras en estudio. Se amplificaron fragmentos con selección de cebadores +3/+3 nucleótidos mediante la utilización de varias combinaciones de cebadores, lo que permitió determinar las combinaciones que generan el mayor número de bandas polimórficas.
Para el estudio de variabilidad genética de las especies de ñame (D. alata L., D. rotundata Poir., D. esculenta (Lour.) Burkill y D. trifida L.f.) la reacción de amplificación se realizó utilizando combinaciones diferentes de pares de cebadores. Se encontró que la combinación de cebadores en los tratamientos E-ACA/M-CAT, E-AAC/M-CAC, E-AAG/MCTC, presentaron un mayor polimorfismo, por lo tanto éstas se escogieron para dicho estudio. El estudio de diversidad genética para la especie D. alata se realizó con la combinación de cebadores más polimórficos, E-ACA/M-CAT, en 14 accesiones.
El programa de PCR para el inicio del ciclo +3/+3 fue el siguiente: un ciclo a 94 °C por 30 s, 65 °C por 30 s, y 72 °C por 60 s. La temperatura de apareamiento se redujo 0.7 °C en cada ciclo durante 12 ciclos y se realizaron 23 ciclos para un tiempo total de 2 h, 2 min, así: 94 °C por 30 s, 56 °C por 30 s y 72 °C por 60 s, el producto amplificado se almacenó, finalmente, a 4°C. Las reacciones de amplificación fueron reveladas en un gel de poliacrilamida al 6% teñido con plata, a partir del cual se capturó la distribución de los loci que estuvieron en un rango de lectura entre 40 y 330 pb.
Análisis de resultados. Debido a la naturaleza dominante de los marcadores AFLP, las matrices de datos binarios fueron codificadas para cada nivel mediante la presencia (1) o ausencia (0) de bandas originadas en la amplificación de los fragmentos digeridos. Con la matriz de datos presencia/ausencia se calculó el índice de similitud de Dice, adaptado por Nei y Li (1979) para datos moleculares. Este índice promedia los valores de similitud por par de individuos, mediante la ecuación siguiente:
Sij = 2a/(2a+b+c), donde Sij = similitud entre los individuos i y j;
a = número de loci compartidos por i y j;
b = número de loci presentes en i pero ausentes en j; y
c = número de loci presentes en j pero ausentes en i.
Las matrices y los dendrogramas de similitud se construyeron con el programa NTSYS-PC, versión 2.02i (Rohlf, 1998) mediante el método UPGMA y el agrupamiento SAHN, respectivamente. Se analizaron además, las relaciones entre individuos mediante correspondencia múltiple (ACM) con toda la población, para obtener una representación gráfica de la distancia entre las accesiones. Para estimar la confiabilidad de los grupos genéticos designados en los dendrogramas se realizó la confirmación del análisis de conglomerados y de los grupos de diversidad mediante remuestreo (1000 permutaciones), para lo cual se utilizó el software WinBoot (Nelson, 1996).
Polimorfismo de los AFLP
Las combinaciones de cebadores empleados en esta investigación presentaron patrones altamente polimórficos entre y dentro de las especies, mostrando así perfiles de loci diferentes, garantizando un buen poder de discriminación e identificando grupos genéticos disímiles de las especies de Dioscorea spp. Los fragmentos de ADN amplificados mediante AFLP oscilaron en un rango de 40 hasta 330 pb. La combinación de cebadores E-ACA/M-CAT y E-AAC/M-CAC presentaron el mayor porcentaje de polimorfismo, el número total de loci que se obtuvo en todo el estudio fue de 206 fragmentos (Cuadro 2), los que se extendieron en un rango de 74 loci para la combinación de cebadores E-ACA/MCAT, hasta 64 loci para la combinación de cebadores E-AAG/M-CTC, con un promedio de 68.66 loci por par de cebadores. La combinación de cebadores E-AAC/M-CAC (Figura 1) mostró mayor polimorfismo, obteniéndose un total de 68 loci, de los cuales 64 mostraron polimorfismo (94.11%). Utilizando la combinación de cebadores E-ACA/M-CAT se obtuvo un 91.89% de polimorfismo y 74 loci en total, de los cuales 68 mostraron polimorfismo (91.89%). Estos resultados son similares a los encontrados por Tamiru et ál. (2007) en ñame cuando cuantificaron polimorfismo por encima del 90% y resaltaron el alto número de bandas polimórficas encontradas cuando se usan estas combinaciones de cebadores.
Diversidad genética inter-específica de Dioscorea spp.
En este trabajo se destaca la importante variación que tiene Dioscorea spp. en su amplia distribución en la región Caribe colombiana. La caracterización molecular mediante la técnica AFLP tuvo una alta sensibilidad lo que permitió separar las accesiones de ñame en cuatro grupos, según la especie a la que pertenecen: D. alata, D. rotundata, D. esculenta y D. trifida. Las comparaciones de similitudes genéticas entre las cuatro especies se presentan en el Cuadro 3. Los valores promedios oscilaron entre el 41.81% entre D. alata y D. rotundata, y 33.51% entre D. trifida y D. esculenta. Estos resultados son consistentes con la clasificación de las especies con respecto a sus secciones botánicas, D. alata y D. rotundata perteneciente a la sección Enanthiophillum del género Dioscorea spp., especies cuyo centro de origen es el sudeste asiático y áfrica occidental; mientras que D. trifida pertenece a la sección Macrogynodium originaria de América tropical y D. esculenta a la sección Combilium, originaria del sudeste asiático. La comparación de similitud genética entre pares de especies indica que D. alata e s g enéticamente m ás c ercana d e D. rotundata (41.81%) que de D. esculenta (32.21%), mientras que D. trifida se encuentra genéticamente mucho más distante de D. alata (29.95%) (Cuadro 3). Esta relación se ve soportada por el análisis de conglomerados (Figura 1) de los valores de confianza 'bootstrap' de la matriz de distancia 100% para la relación entre D. alata vs. D. rotundata, 49.8% para la relación entre D. esculenta con ambas especies, y por último 97.9% D. trifida. Los niveles de variabilidad entre grupos taxonómicos han sido estudiados por Malapa et ál. (2005) quienes encontraron variabilidad genética entre grupos taxonómicos en Dioscorea spp., y discriminan seis especies de la sección Enantiophyllum mediante el uso de marcadores AFLP (Vos et ál., 1995). Estos resultados eran de esperar debido a la alta variabilidad del género Dioscorea, como lo demostraron Sonibare et ál. (2010) cuando realizaron estudios de variabilidad genética entre el género Dioscorea a partir de muestras recolectadas al oeste y centro de áfrica. Hildebrand et ál. (2002) encontraron alto grado de diversidad en variedades criollas de ñame al suroeste de Etiopía, separando 23 tipos de ñames nativos.
El análisis de correspondencia múltiple (ACM) estimó la variación de los individuos en tres dimensiones (ejes x, y, z) (Figura 2) y muestra información similar a la obtenida en el dendrograma diferenciando cuatro grupos. Los resultados obtenidos en esta investigación confirman y son consistentes con los obtenidos por Durango y Padilla (1998) en caracterizaciones morfológicas previas en los mismos genotipos. En el grupo 1 se observan 14 accesiones (9506-27 a 9605-54) con una tendencia de agrupamiento hacia el centro de la coordenada conformado por la especie D. alata, con un índice promedio de similitud del 89.51% (Cuadro 3) y caracterizadas por tener tallo cuadrangular sin espinas con enrollamiento dextral y presentar cuatro alas o pliegues, hojas con ápices muy agudos, algunas accesiones producen tubérculos subterráneos de formas cilíndricas, esféricas, deltoides e irregulares de color café. Estos resultados concuerdan con los hallazgos de Egesi et ál. (2003) quienes encontraron diferencias significativas en color, sabor, consistencia, harinosidad y viscosidad en tubérculos de 40 accesiones en D. alata. Estos tubérculos tienen buena aceptación en el mercado local y potencial de exportación, pero algunos genotipos son susceptibles a la antracnosis por el hongo Colletotrichum gloeosporioides (Campo et ál., 2009).
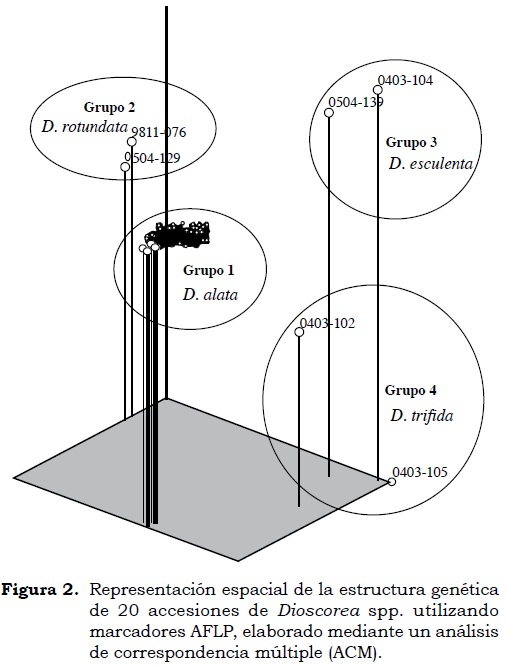
El grupo 2, conformado por la especie D. rotundata (Figura 2), incluye dos accesiones (9811-076 y 0504-129) que comparten un índice promedio de similitud del 79.12% (Cuadro 3) y se caracterizan por presentar tallo redondo, delgado, con espinas y enrollamiento dextral. Las hojas son más anchas en la parte superior, no tiene espinas en el pecíolo, presentan inflorescencia en forma de espiga simple. El genotipo 9811-076 presentó tubérculos subterráneos de forma cilíndrica, epidermis de color marrón y pulpa de color blanca. El genotipo 0405-129 produjo tubérculos de buenas características organolépticas de gran aceptación en el mercado local e internacional, pero es susceptible a la enfermedad foliar por antracnosis (Campo et ál., 2009).
El grupo 3, conformado por la especie D. esculenta (Figura 2), incluye dos accesiones (0403-104 y 0504-139) que comparten un índice promedio de similitud del 82.22% (Cuadro 3). En este grupo se observa una menor dispersión debido a una mayor cercanía genética entre ambas accesiones. Estas se caracterizan por tener tallo redondo de color marrón y presencia de espinas, con enrollamiento sinestral. Las hojas son anchas, con presencia de espinas en el pecíolo, produce varios tubérculos subterráneos pequeños de color marrón claro y no se le conocen floración ni presencia de tubérculos aéreos.
El último conglomerado está conformado por la especie D. trifida (Grupo 4) e incluye dos accesiones (0403-105 y 0403-102) que comparten un índice promedio de similitud del 55.56% (Cuadro 3). Se observa que las accesiones que conforman este grupo se encuentran dispersas a lo largo de la coordenada (ejes x, y, z) (Figura 2) con cierto grado de separación de accesiones de la misma especie. Estas accesiones se caracterizan por presentar tallo de color verde, con dos alas de color marrón; al igual que D. esculenta tiene enrollamiento sinestral, es muy rustica y las hojas presentan cinco lóbulos. Esta especie produce flores de tipo hermafrodita, los tubérculos son de forma oval, oblongos y de color marrón, en forma de racimos que penden de varias raíces que cuelgan de la base del tallo.
Los resultados obtenidos en esta investigación son coincidentes con los de Malapa et ál. (2005) quienes evaluaron la diversidad genética en Dioscorea spp., mediante la utilización de marcadores AFLP, encontrando consistencia entre la sistemática de la especie y sus respectivas secciones botánicas. De la misma forma Tamiru et ál. (2007) evaluaron diversidad y estructura genética en ñame procedente de Etiopía, relacionándolas con especies del oeste de áfrica, encontrando alta variabilidad genética en las accesiones avaluadas. Resultados similares obtuvieron Mignouna et ál. (2002) mediante la utilización de marcadores enzimáticos que confirman la variabilidad genética en genotipos de Dioscorea cayenensis Lam y Dioscorea rotundata. En el presente estudio se comprobó la amplia variabilidad del género Dioscorea spp., pertenecientes a la colección de accesiones de ñame de la Universidad de Córdoba, las cuales fueron recolectadas en los departamentos de Córdoba, Sucre y Bolívar (Colombia) y a su vez, se demostró que estos resultados moleculares presentan similitudes con las caracterizaciones morfológicas previas y la clasificación botánica. No obstante, estos resultados se diferencian de los reportados por Bustamante et ál. (2001) donde empleando el marcador molecular DNA amplificación 'fingerprinting' (DAF) encontró alta similaridad en los genotipos estudiados procedentes de la colección de accesiones de ñame de la Universidad de Córdoba y algunas muestras provenientes del IITA (International Institute of Tropical Agriculture, Ibadan, Nigeria).
Diversidad genética de Dioscorea alata L.
Estudios en otras especies han permitido estimar la variabilidad genética en especies cultivadas comerciales de ñame en diferentes regiones (Arnau et ál., 2009; Tostain et ál., 2006). Esto sugiere que es posible utilizar marcadores moleculares que presenten un mayor polimorfismo en las bandas generadas, con el fin de diferenciar genotipos dentro de la especie de D. alata (Malapa et ál., 2005). En este estudio se hizo el análisis de similaridad de los datos a través de AFLP (Figura 3) para evaluar la relación dentro de las accesiones de D. alata. Esta agrupación mostró cuatro grupos principales con cierto grado de variabilidad genética. El grupo 1 está conformado por los genotipos 9506-027, 0406-100, 0406-002, 0406-009, 0406-098, 0406-062, 0406-094, recolectados en el departamento de Córdoba- Colombia, y 0406-130 recolectados en el departamento de Bolívar (Colombia). Se separa de las demás accesiones un primer subgrupo representado por los genotipos 9506- 027 y 0406-100, caracterizado por presentar flores masculinas y presencia en el tallo de alas de color verde, tubérculos subterráneos de color de pulpa blanca; este grupo presenta similaridad con un valor de 0.976. Un segundo subgrupo (Figura 3) conformado por los genotipos 9404-002, 9504-009, 9603-037, 9605- 062 y 0406-094 presenta un valor de similaridad de 0.972 y se caracteriza por presentar floración femenina, el tallo es cuadrado y presenta alas muy pequeñas de color morado; este genotipo es completamente rústico. Las hojas muestran ápice agudo y sus flores son de color amarillo, simples y en forma de racimo. Los tubérculos son de forma irregular, poseen vellosidad y muchas raíces de fijación. Estos resultados son coincidentes con los de Malapa et ál. (2005), quienes demostraron que esta es una especie altamente heterogénea, presentando alta variabilidad genética dentro de la especie, lo que fue evidente en la morfología de las accesiones estudiadas donde se revelaron características fenotípicas muy variables como color, forma y tamaño de hojas, tallos y tubérculos. Los genotipos 9404-002, 9504-009 y 9603-037 presentan resistencia a la antracnosis, lo que es de gran importancia agronómica, ya que dichas accesiones pueden ser tenidas en cuenta para futuros trabajo de fitomejoramiento que generen cultivares mejorados resistente al hongo C. gloeosporioides. Esta resistencia fue evaluada mediante estudios de incidencia y severidad de la enfermedad por Campo et ál. (2009). En la región Caribe de Colombia es muy conocido con diferentes nombres vulgares entre los cuales están 9404-002 ('ñame pepita'), 9504-009, 9603-037, y 0406-094 ('mampuján'), 9605-062 ('ñame manteco'), y el 0406-130 ('te encontré'). Los resultados muestran que las accesiones 9504-009 y 9603-037 son el mismo genotipo, lo cual se comprueba por tener un coeficiente de similaridad de 1 y compartir semejanzas morfológicas como coloración verde y morado en el tallo; hojas simples de forma acorazonada y ápice agudo; flores de color amarillo, simples y compuestas; tubérculos de gran tamaño con vellosidad y muchas raíces de fijación; y resistencia a antracnosis.
El grupo 2, conformado por la accesión 0406-008 (Ñame peludo) colectado en el departamento de Córdoba-Colombia revela un coeficiente de similaridad de 0.82. Tiene como característica principal la forma cuadrada del tallo, su color es verde con pigmentos marrón, presenta alas muy pequeñas, su crecimiento es indeterminado y alcanza de cuatro a seis metros de largo y muestra enrollamiento de forma dextral. Las hojas son de forma sagitiforme y lobulado poco profundo separado por un ápice muy agudo. Sus f lores son femeninas, los tubérculos son ramificados, la pulpa es de color blanca y textura lisa, posee vellosidad y raíces de fijación de color marrón oscuro, su forma es irregular, también posee tubérculos aéreos de forma redonda.
El grupo 3, conformado por la accesión 0406-054 ('ñame seda') recolectado en el departamento de Magdalena, Colombia, presenta un coeficiente de similaridad de 0.871, tiene como característica principal la ausencia de bulbillos aéreos y de raíces secundarias en los tubérculos, posee precocidad y tiene espinas en la base de los tallos jóvenes.
El grupo 4, conformado por las accesiones 9502-005 recolectadas en el departamento de Sucre y la accesión 9506-022 y 0504-140 recolectadas en el departamento de Córdoba, Colombia, con un coeficiente de similaridad de 0.92 comparte las características de formación de bulbillos aéreos y presencia de raíces secundarias en los tubérculos, con resistencia al hongo C. gloeosporioides (Campo et ál., 2009).
En estudios previos que incluyeron criterios fenotípicos (Lebot et ál., 1998; Malapa, 2000) se demostró una amplia variabilidad morfológica en D. alata donde se destaca la alta variabilidad en el tubérculo. Esta variación, si bien puede ser importante como fuente de variabilidad genética en procesos de mejoramiento (Malapa et ál., 2005), debe ser considerada de manera particular con el fin de preservar las características agronómicas y de producción del cultivo.
El alto grado de intercambio de semilla procedente de los departamento de Córdoba, Sucre, Bolívar y el Magdalena por parte de los agricultores, indica que los clones han sido ampliamente distribuidos en estas zonas, no obstante, esta afirmación se debe tomar con precaución ya que no hay poblaciones suficientemente grandes que incluyan diversos transeptos geográficos de la región Caribe colombiana que permitan hacer un estudio más detallado de dicha variabilidad.
- La técnica AFLP permitió detectar alta variabilidad genética en las veinte accesiones de ñame evaluadas, permitiendo diferenciar especies mediante un patrón de organización morfológico bien definido en cuatro grupos genéticos pertenecientes a las especies D. alata, D. rotundata, D. esculenta y D. trifida.
- Los grupos genéticos comparten características como presencia o ausencia de alas y espinas en tallos, forma y color de hojas y tubérculos, además se presenta agrupación de algunas accesiones teniendo en cuenta atributos agronómicos como la resistencia a antracnosis. Estos caracteres están más relacionados con las características de tipos cualitativos y cuantitativos propios de cada genotipo y no de acuerdo con la ubicación geográfica de los sitios de recolección. La variabilidad genética encontrada en este estudio es de gran utilidad para seguir conservando este recurso genético y mirar las prospecciones de ampliación de la colección.
- La caracterización molecular también permitió detectar variabilidad genética entre las accesiones pertenecientes a la especie D. alata, donde se aprecia una alta divergencia morfológica como presencia de dioicismo, color y forma de tubérculos. Esta información generada puede ser de gran utilidad para el desarrollo de estrategias en futuros programas de fitomejoramiento.
Los autores expresan sus agradecimientos a la Universidad de Córdoba por la financiación de esta investigación a través del Proyecto Obtención de Genotipos de Ñame Dioscorea alata L., Resistentes a Antracnosis, Colletotrichum gloeosporioides Penz y Sacc, en la costa norte colombiana. Igualmente, al personal del grupo técnico de producción de ñame del Programa de Ingeniería Agronómica de la Universidad de Córdoba (Colombia), y al personal del Laboratorio de Biología Molecular del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, al CIAT, Palmira, Valle del Cauca (Colombia).
Arnau, G; Nemorin, A; Maledon, E; y Abraham, K. 2009. Revision of ploidy status of Dioscorea alata L. (Dioscoreaceae) by cytogenetic and microsatellite segregation analysis. Theor Appl Genet 118: 1239 - 1249. [ Links ]
Bradley, D.; Ratchliff, O.; Vincent, C.; Carpenter, R.; y Coen, E. 1997. Inflorescence commitment and architecture in Arabidopsis. Science 275: 80 - 83. [ Links ]
Bustamante, S; Guzmán, M.; y Buitrago, G. 2001. Caracterización molecular de algunas especies y variedades de ñame presentes en la Costa Atlántica Colombiana. Rev. Col. Biot. 3(2): 38 - 43. [ Links ]
Bustamante, S; Guzmán, M.; y Buitrago, G. 2003. Caracterización molecular del germoplasma de ñame colombiano utilizando DNA Amplificaron Fingerprinting (DAF) en condiciones radiactivas. Rev. Col. Biot. 5(2): 57 - 63. [ Links ]
Campo, R.; Luna, J. M.; y Jiménez, Y. 2009. Selección de genotipos de ñame Dioscorea spp. resistentes a la antracnosis (Colletotrichum gloeosporiodes Penz). Fitop. Col. 33(1): 7 - 10. [ Links ]
Dansi, A.; Mignouna, H. D.; Zoundjihekpon, J.; Sangare, A.; Asiedu, R.; y Ahoussou, N. 2000. Using isozyme polymorphism to assess genetic variation within cultivated yams (Dioscorea cayenensis/ Dioscorea rotundata complex) of the Republic of Benin. Gen. Res. Crop Evol. 47: 371 - 383. [ Links ]
Durango, R. y Padilla, A. 1998. Caracterización morfológica de un banco de germoplasma de ñame Dioscorea spp. recolectado en la costa norte colombiana. Trabajo de grado. Universidad de Córdoba, Montería, Colombia. 69 p. [ Links ]
Egesi, C. N.; Asiedu, R.; Egunjobi, J. K.; y Bokanga, M. 2003. Genetic diversity of organoleptic properties in water yam (Dioscorea alata L.). J. Sci. Food Agric. 83: 858 - 865. [ Links ]
Hamon, P. y Toure, B. 1990a. Characterization of traditional yam varieties belonging to the Dioscorea cayenensis-rotundata complex by their isozymic patterns. Euphytica 46: 101 - 107. [ Links ]
Hamon, P. y Toure, B. 1990b. The classification of cultivated yams (Dioscorea cayenensis-rotundata complex) of West Africa. Euphytica 47: 179 - 187. [ Links ]
Hildebrand, E.; Demissew, S.; y Wilkin, P. 2002. Local and regional disappearance in species of Dioscorea L. (Yams) in southwest Ethiopia. En: Stepp, J. R.; Wyndham, F. S.; Zarger, R. R. (eds.). Ethnobiology and biocultural diversity. Proceedings of the 7th international congress of ethnobiology. University of Georgia Press. p. 678 - 695. [ Links ]
Lebot, V.; Trilles, B.; Noyer, J. L.; y Modesto, J. 1998. Genetic relationships between Dioscorea alata L. cultivars. Gen. Res.. Crop Evol.. 45: 499 - 508. [ Links ]
Malapa, R. 2000. Etude de la diversite genetique des cultivars de D. alata L. du Vanuatu par les marqueurs morpho-agronomiques et AFLP. DEA de Genetique, Adaptation et Productions Vegetales, ENSA de Rennes I, France, 29 p. [ Links ]
Malapa, R.; Arnau, G.; Noyer, J. L.; y Lebot, V. 2005. Genetic diversity of the greater yam (Dioscorea alata L.) and relatedness to D. nummularia Lam. and D. transversa Br. as revealed with AFLP markers. Gen. Res. Crop Evol. 52: 919 - 929. [ Links ]
Martin, F. W.; Rhodes, A. M. 1977. Intra-specific classification of Dioscorea alata. Trop. Agric. 54: 1 - 13. [ Links ]
Mignouna, H. D.; Ellis, N. T. H.; Knox, M. R.; Asiedu, R.; y Ng, Q. N. 1998. Analysis of genetic diversity in Guinea yams (Dioscorea spp.) using AFLP finger printing. Trop. Agric. 75: 224 - 229. [ Links ]
Mignouna, H. D.; Dansi, A.; Zok, S. 2002. Morphological and isozymic diversity of the cultivated yams (Dioscorea cayenensis/Dioscorea rotundata complex) of Cameroon. Genet. Resourc. Crop Evol. 49: 21 – 29. [ Links ]
Mignouna, H. D.; y Dansi, A. 2003. Yam (Dioscorea spp.) domesticated by the Nago and Fon ethnic groups in Benin. Gen. Res. Crop Evol. 50: 519 - 528. [ Links ]Nei, M.; y Li, W. H. 1979. Mathematical model for studying genetic variation in terms of restriction endo nucleases. Proc. Nat. Acad. Sci. United States of America 76(10): 5269 - 5273.
Mignouna, H. D.; Abang, M. M.; Fagbemi, S. A. 2003. A comparative assessment of molecular marker assays (AFLP, RAPD and SSR) for white yam (Dioscorea rotundata) germplasm characterisation. Annals of Applied Biology 142: 269 - 276. [ Links ]
Nei, M.; y Li, W. H. 1979. Mathematical model for studying genetic variation in terms of restriction endo nucleases. Proc. Nat. Acad. Sci. United States of America 76(10): 5269 - 5273. [ Links ]
Nelson, R. J. 1996. WinBoot Yap. IV. a program for performing bootstrap analysis of binary data to determine the confidence limits of UPGMA-based dendrograms, IRRI, Manila Filipinas, 1996. http://www.irri.org/science/software/winboot.asp. [ Links ]
Okoli, O. O. 1991. Yam germplasm diversity, uses and prospects for crop improvement in Africa. p. 109-117. En Ng, N. Q. et ál. (eds.). Crop genetic resources of Africa. Vol. 2. Proc. of an Int. Conf. on Crop Genetic Resources in Africa, Ibadan, Nigeria. 17-20 Oct. 1988. IITA/IBPGR/UNEP/ CNR, Ibadan, Nigeria. [ Links ]
Ramser, J.; Weising, K.; Lopez-Peralta, C.; Terhalle, W.; Terauchi, R.; y Kahl, G. 1997. Molecular marker based taxonomy and phylogeny of Guinea yam (Dioscorea rotundata-D. cayenensis). Genome 40: 903 - 915. [ Links ]
Rohlf, F. 1998. NTSYSpc. Numerical taxonomic and multivariated analysis system, vol 2.0, Exeter Software, Setauket, Nueva York. [ Links ]
Sonibare, M.; Asiedu, R.; y Albac, D. 2010. Genetic diversity of Dioscorea dumetorum (Kunth) Pax using Amplified Fragment Length Polymorphisms (AFLP) and cpDNA. Bioch. Syst. Ecol. 38: 320 - 334. [ Links ]
Tamiru, M. 2006. Assessing diversity in yams (Dioscorea spp.) from Ethiopia based on morphology, AFLP markers and tuber quality, and farmers' management of landraces. PhD thesis, Georg- August-University Goettingen, Germany. Cuvillier Verlag, Goettingen, Germany, 155 p. [ Links ]
Tamiru, M.; Becker, H.; y Maass, B. 2007. Genetic diversity in yam germplasm from Ethiopia and their relatedness to the main cultivated Dioscorea species assessed by AFLP markers. Crop Sci. 47: 1744 - 1753. [ Links ]
Tostain, S; Scarcelli, N; Brottier, P; Marchand, J. L.; Pham, J. L.; y Noyer, J. L. 2006. Development of DNA microsatellite markers in tropical yam (Dioscorea sp.). Mol. Ecol. Notes 6: 173 - 175. [ Links ]
Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; y Zabeau, M. 1995. AFLP: A new technique for DNA fingerprinting. Nucl. Acids Res. 23: 4407 - 4414. [ Links ]
